










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkColombia Médica
On-line version ISSN 1657-9534
Colomb. Med. vol.39 suppl.3 Cali Sept. 2008
Alteración de la organización laminar y de la dendroarquitectura de la corteza cerebral del humano post-trauma craneoencefálico
Martha Isabel Escobar, MSc1, Francisco Guzmán, MD2, Efraín Buriticá, MSc3, David Riascos, MD4, Liliana Villamil, MD4, Hernán Pimienta, MSc5
1. Profesora Titular, Investigadora Centro de Estudios Cerebrales, Universidad del Valle, Cali, Colombia.
e-mail: maiesbe@yahoo.com
2. Investigador, Centro de Estudios Cerebrales, Universidad del Valle, Cali, Colombia. e-mail: frangupe@hotmail.com
3. Estudiante de doctorado, Centro de Estudios Cerebrales, Universidad del Valle, Cali, Colombia.
e-mail: triburi@yahoo.com
4. Estudiante de Maestría, Centro de Estudios Cerebrales, Universidad del Valle, Cali, Colombia.
e-mail: davisdarm@gmx.net lilika_villamil@hotmail.com
5. Profesor Distinguido, Director Centro de Estudios Cerebrales, Facultad de Salud, Universidad del Valle, Cali, Colombia. e-mail: hernpim@yahoo.com
Recibido para publicación diciembre 20, 2007 Aceptado para publicación junio 26, 2008
RESUMEN
Introducción: El trauma craneoencefálico (TCE)es un fenómeno heterogéneo desde el punto de vista molecular, celular y en la respuesta clínica. Se considera que esta diversidad se debe a la intensidad de la injuria primaria, eventos secundarios asociados (hipoxia, isquemia, edema, inflamación), al estado metabólico del paciente, su base genética, edad, género, etc. Para determinar la integridad anatomo-funcional de las células nerviosas es importante verificar el estado de la cito, dendroarquitectura y preservación laminar como un requisito para garantizar conectividad.
Objetivo: Valorar la respuesta de las neuronas al trauma con dos marcadores neuronales selectivos sensibles a la lesión NeuN y MAP2.
Materiales y métodos: Se utilizaron muestras (4 de lóbulo temporal y 2 de lóbulo frontal) de 6 pacientes que habían sufrido TCE. Las muestras se fijaron en PLP, cortadas en vibrátomo a 50 µm, incubadas con los anticuerpos NeuN y MAP2 y procesadas con el sistema avidina-biotina. Como control se utilizó tejido humano post-mortem.
Resultados: La inmunorreactividad (IR) para NeuN fue anormal en todas las muestras, con sectores que mostraron IR ligeramente alterada, otros con perdida parcial de las capas supragranulares, sobre todo la lámina III y otros con pérdida drástica de todas las láminas. La IR para MAP2 se alteró en todas las muestras con diferentes grados de compromiso. Los procesos dendríticos fueron difíciles de seguir, especialmente los procedentes de la lámina V, los cuales se observaron tortuosos, fragmentados y con orientación aberrante.
Conclusiones: Con el propósito de conocer el estado de las neuronas después de un evento lesivo se recomienda el uso de los marcadores NeuN y MAP2 complementarios a los métodos clásicos. El presente trabajo muestra la diversidad de respuestas histopatológicas en sectores adyacentes de una misma muestra con ambos marcadores, como un indicador de los diferentes estados de neurodegeneración.
Palabras clave: Trauma craneoencefálico; MAP2; NeuN; Corteza cerebral humana.
Changes in laminar and dendritic organization of the cerebral cortex after traumatic brain injury
SUMMARY
Introduction: Traumatic brain injury (TBI) is a heterogeneous phenomenon from a molecular, cellular and pathological perspective. Clinical outcome is also extremely variable. It is considered that such a diversity response to TBI is related to the primary injury intensity, associated secondary events (hypoxia, ischemia, oedema and inflammation), metabolic patient state, genetic background, age, gender, etc. After injury the histopathological outcome is variable in time and space. In order to determine the anatomofunctional integrity of nerve cells in the cerebral cortex, it is important to verify the state of the cito and dendroarchitecture and the laminar preservation as a requisite to guarantee connectivity.
Objective: The aim of the present work was to evaluate the response of human cortical neurons using two selective neuronal markers, NeuN and MAP2, which recognize citoarchitecture and dendritic arrangement, respectively.
Materials and methods: In the present study we utilized six tissue samples (4 temporal and 2 frontal cortices) from TBI patients. Tissues from four post-mortem human brains were used as controls. Tissue samples were fixed in PLP, cut at 50 um in a vibratome, incubated with NeuN and MAP2 and processes with the avidin/biotin complex.
Results: NeuN-IR was abnormal in all samples analyzed with some sectors showing slight NeuN-IR, others with NeuN-IR partial loss in supragranular layers, especially layer IIII, and other with a drastic reduction in staining in all cortical layers. MAP2-IR was altered across sections with sectors showing different degrees of changes in the normal pattern of MAP2-IR. Dendritic processes were difficult to follow because of its discontinuity. Layer V apical dendritic processes appear tortuous and its IR was fragmented in some cases they take aberrant orientations.
Conclusions: in order to get new insights about the neuronal outcome after brain injury it is recommended to use MAP2- and NeuN-IR as a complement of classical histopathological methods. The present study shows a diversity of histopathological response in adjacent sectors of a same sample with both markers MAP2 and NeuN, as an indicative of different states of neurodegeneration.
Keywords: Traumatic brain injury; MAP2; NeuN; Human cerebral cortex.
El trauma craneoencefálico (TCE) es un problema de salud a nivel mundial. En los Estados Unidos se presentan 1500,000 casos nuevos cada año, de los cuales 3.3% son fatales y 6% presentan incapacidad permanente1. Los datos mundiales indican que la lesión cerebral estructural más frecuente en las personas con TCE es la contusión cerebral y su mortalidad si no se maneja adecuadamente llega a cifras tan elevadas como 60%2. Las personas que sobreviven al TCE pueden presentar secuelas psicológicas, neurológicas y psiquiátricas tales como depresión, déficit de memoria, epilepsia y mayor riesgo para desarrollar demencia3,4. El sustrato biológico para estos eventos no está claramente establecido; sin embargo, se admite que es el resultado de procesos que se instauran en el momento del trauma y se continúan, lo cual induce muerte neuronal en las fases aguda, subaguda y crónica. La lesión primaria, al momento del impacto, ocasiona muerte neuronal, glial, lesión axonal y vascular. En zonas adyacentes o distantes o relacionadas por la conectividad, se pueden presentar eventos tardíos o secundarios como edema cerebral, hipoxia, aumento de la presión intracraneal, modificaciones del flujo sanguíneo cerebral, etc5. Estos eventos se pueden relacionar con alteración en la homeostasis iónica, disfunción mitocondrial, alteraciones del citoesqueleto, del ADN y deaferentación. Estos procesos continúan por días, meses y/o años. La mayor parte del conocimiento que se tiene actualmente con respecto a los mecanismos fisiopatológicos del TCE se ha derivado de estudios en modelos animales o de tejido cerebral de humano post-mortem. En los últimos años se han desarrollado estudios a nivel molecular en tejido cerebral de humano posterior a lesión traumática, en los cuales se ha evaluado la activación de calpainas6, la expresión de proteínas asociadas con cascadas apoptóticas como caspasa 7, Bcl2, proteínas de la vía de supervivencia como NFk-ß y de la proteína precursora de amieloide (APP)7.
Los anticuerpos contra las proteínas NeuN y MAP2, que identifican neuronas de manera selectiva, se utilizan ampliamente en modelos experimentales de trauma y de isquemia como marcadores de injuria y de su evolución temporal8,9. En tejido lesionado la inmunorreactividad (IR) para estos dos marcadores decrece en las zonas de impacto y las perilesionales y su disminución se asocia con la intensidad de la lesión a nivel neuronal o como predictores de muerte neuronal10,11. En el presente estudio se evalúa la expresión inmunohistoquímica de NeuN y MAP2 en muestras de corteza cerebral de humano obtenidas de cirugía decompresiva después de TCE, con el fin de establecer la integridad cito y dendroarquitectónica de las zonas contusas y la presencia o no de vulnerabilidad laminar selectiva. Los datos obtenidos en el presente trabajo aportan información sobre los cambios celulares en estados agudos y subagudos en el humano post TCE, permitiendo inferencias sobre alteraciones en la conectividad en zonas traumáticas con aparente conservación de su organización citoarquitectónica.
MATERIALES Y MÉTODOS
El presente estudio se realizó en 6 muestras de tejido contuso derivadas de 6 personas con diagnóstico de contusión cerebral focal, con hipertensión endocraneana (HTEC) severa y que, de acuerdo con el criterio del neurocirujano tratante, se le podía ofrecer como alternativa quirúrgica una craniectomía decompresiva con apertura y/o plastia de la dura, y/o drenaje del parénquima cerebral contuso para el manejo de la HTEC secundario al efecto focal de la contusión. En ningún caso, se hizo remoción adicional de tejido cerebral para efectos del presente estudio.
Criterios de inclusión de la muestra
- Haber ingresado al Hospital Universitario del Valle (HUV) y el paciente estar a cargo del Instituto Neuroquirúrgico del HUV.
- Haber sufrido un TCE contuso.
- Tener una contusión cerebral con criterios quirúrgicos para el neurocirujano tratante.
- Tener consentimiento firmado por el paciente o acudientes tanto para la realización de la cirugía como para la inclusión en el estudio.
- No tener contraindicaciones para la cirugía.
- No cumplir los criterios de exclusión.
Criterios de exclusión de la muestra
- Antecedente de consumo de psicotrópicos y alcoholismo.
- Antecedente de infarto cerebral previo, malformaciones vasculares, angiopatía amiloidea, aneurismas o tumores cerebrales.
- Glasgow de 3 y/o ausencia de reflejos de tallo.
- Coagulopatías.
- Hipoxia cerebral o hipotensión durante más de 10 minutos confirmada desde el trauma hasta el momento de la cirugía.
El procedimiento quirúrgico se llevó a cabo de acuerdo con las normas del Comité de Ética de la Facultad de Salud de la Universidad del Valle y del HUV que respetan los principios y procedimientos de la Declaración de Helsinki de la Asamblea Médica Mundial y de la Ley 44 de 1993 de la República de Colombia.
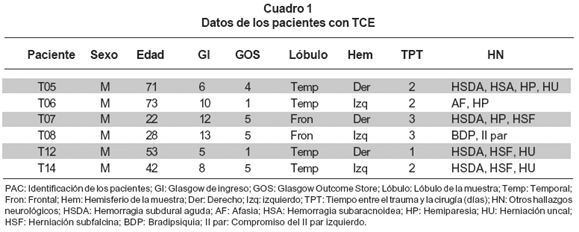
De las 6 muestras de corteza cerebral, 4 eran de corteza temporal y 2 de corteza frontal. Ninguna de las 6 muestras procedía de pacientes que hubieran presentado hipoxia o hipoperfusión antes o durante la cirugía. Todas las muestras eran de pacientes masculinos con edades comprendidas entre 22 y 73 años (promedio de 49.3 años) los cuales fueron operados entre los días 1 y 4 post-trauma, con un promedio de 57 h post-trauma. El Glasgow de ingreso fue menor de 8 para 3 pacientes y entre 10 y 13 para los restantes. El GOS (Glasgow Outcome Score) fue de 1 para 2 pacientes, 5 para 3 y de 4 para 1. De estos pacientes, cuatro presentaron hemorragia subdural aguda, dos acompañados con hemiparesia y tres con herniación uncal (Cuadro 1).
El tejido resecado se lavó en solución salina al 0.9% por unos pocos segundos y se fijó inmediatamente por inmersión en PLP (paraformaldehído, L-lisina, metaperiodato de sodio), pH 7.4 a 4ºC por 7 días. Las muestras se cortaron en un vibrátomo (Vibratome Lancer Serie 1000®) a 50 µm. Los cortes se procesaron de manera seriada para inmunohistoquímica utilizando los anticuerpos 5F9 que identifica la proteína asociada con microtúbulos MAP2 (1:100; cedido por Kenneth Kosik, Santa Bárbara, CA) y el marcador específico del núcleo neuronal anti-NeuN (1:2500, Chemicon). Las secciones se procesaron utilizando el complejo avidina-biotina, se revelaron con diaminobencidina y se contrastado con níquel. El control negativo se realizó procesando las secciones en ausencia del anticuerpo primario, siguiendo el protocolo descrito previamente. Como controles positivos se utilizaron muestras de corteza cerebral procedentes de 4 sujetos post-mortem de género masculino, sin historia clínica de enfermedad neurológica, con edad promedio de 30.8 años y tiempo post-mortem de 10.2 horas. Las secciones se montaron en placas recubiertas con cromoaluminio, se deshidrataron y se cubrieron con laminillas. Tres observadores independientes hicieron el análisis de los tejidos. Las observaciones se hicieron a magnificaciones de 4x, 10x y 40x, y se fotografiaron con una cámara Canon Power Shot A-85. Las imágenes digitalizadas se analizaron con la asistencia del programa Sigma ScanPro 5.
RESULTADOS
En el Cuadro 2 se resumen los hallazgos de las observaciones para ambos marcadores, NeuN y MAP2, en el tejido traumático. Con respecto al marcador NeuN los datos expresados en elCuadro 2 indican que:
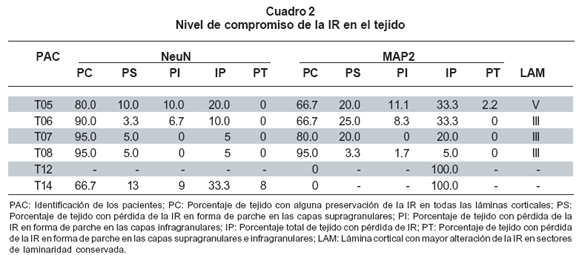
- En todos los sujetos hubo alteraciones de la IR para NeuN.
- La pérdida de la IR es heterogénea, porque en una misma placa se pueden apreciar zonas de pérdida total de la IR para NeuN, zonas de pérdida de IR en forma de parches y sectores donde se conserva la laminaridad (Figura 1).
- De los sectores donde hay pérdida de la IR para NeuN, ésta puede darse en las capas supragranulares (dos de seis muestras) o en capas supra e infragranulares (tres de seis muestras). Es decir, que en cinco de seis sujetos, se encontró compromiso al menos de las capas supragranulares. Es de anotar que en la muestra T12 si bien no se encontraron parches, se presentó una disminución extensa de la IR.
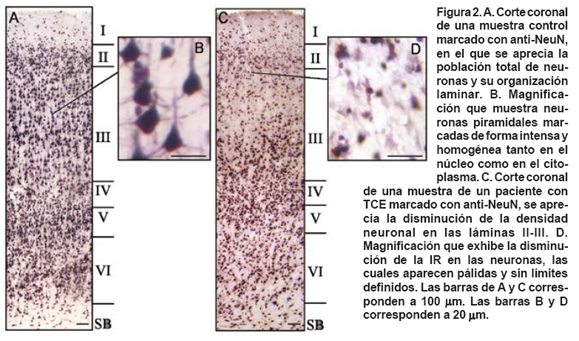
- En todos los sujetos se apreciaron sectores con preservación de las láminas corticales. Con excepción del sujeto T12, en los sectores de citoarquitectura conservada observados a baja magnificación (4x), la IR tiene una apariencia normal; sin embargo, al analizarlo con mayor magnificación (40x) el patrón de IR para NeuN se aleja del patrón normal de expresión de este marcador. Es decir, mientras en los sujetos control la IR para NeuN es intensa y homogénea en el núcleo, y un poco menos intensa en el citoplasma pero igualmente homogénea, en las muestras de tejido traumático se aprecia que la intensidad de la reacción en el núcleo está disminuida, discontinua o fragmentada, y en el citoplasma aumentada respecto del núcleo, aunque igualmente discontinua (Figura 2). Este hecho fue más notable en la lámina III, en tres de las cuatro muestras donde fue posible evaluar esta característica.
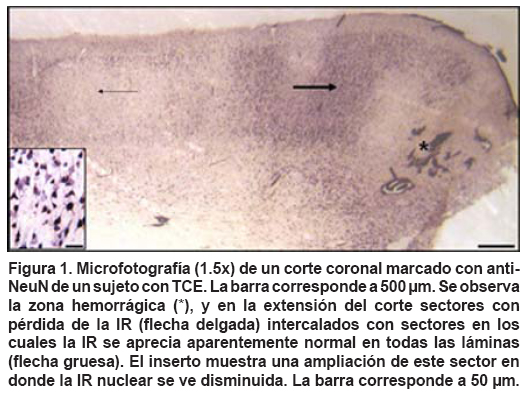
En sectores en los cuales aparece más de un parche de pérdida de la IR, por lo general estos se observan limitados por zonas con citoarquitectura conservada (Figura 1). En cinco de las seis muestras donde aparecen parches de pérdida de IR con NeuN, se encontraron microhemorragias localizadas en la sustancia gris.
La inmunohistoquímica para la proteína asociada con los microtúbulos MAP2, mostró zonas de lesión más extensa y de mayor uniformidad que las mostradas por la proteína NeuN. La IR para MAP2 que en el tejido control se aprecia marcando de manera homogénea los somas, esencialmente de neuronas piramidales y los procesos dendríticos tanto basales como apicales de estas, en el tejido traumático se aprecia disminuida e interrumpida (fragmentada). De esta manera, cuando se observó la expresión de MAP2 en cortes adyacentes a los que mostraron una ligera disminución de la IR para NeuN, se apreció la IR notablemente alterada para la proteína asociada con los microtúbulos MAP2 (Figura 3). De los 6 pacientes analizados, en 3 se encontró pérdida de la IR en parches que comprometió tanto a las capas supra como a las infragranulares y sólo en uno se observó compromiso supragranular. Los dos pacientes restantes presentaron una pérdida generalizada de la IR. Se aprecian interrumpidos (fragmentados) los procesos dendríticos de origen en la lámina V, que ascienden hacia las capas supragranulares formando fascículos edematizados y tortuosos y en algunos sectores con alteración en su dirección, es decir, en lugar de distribuirse de manera vertical adoptan una dirección oblicua (Figura 4).
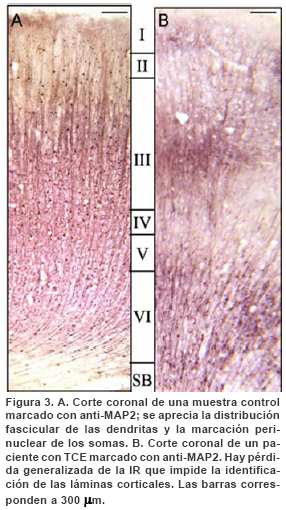
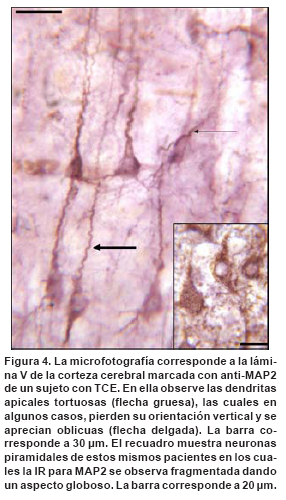
DISCUSIÓN
La histopatología básica del tejido cerebral contuso ha sido bien caracterizada. Esta zona está constituida por sectores hemorrágicos donde se aprecian zonas necróticas caracterizadas por la presencia de acidosis y sectores peri-contusionales donde en ocasiones se aprecia acidosis y más frecuentemente basofilia con la presencia de infiltrados parenquimatosos y vasculares. Aunque en el ser humano se han hecho algunos intentos por dilucidar los mecanismos celulares que se desencadenan posterior a un TCE no es claro hasta el momento en qué estado se encuentran las células de las regiones inmediatamente adyacentes a la contusión (peri-contusionales). Por esta razón, es tan valioso para este tipo de análisis el tejido humano procedente de cirugía decompresiva con resección de la contusión.
El anticuerpo anti-NeuN es un marcador selectivo para neuronas. En el caso de la corteza cerebral marca tanto neuronas piramidales como interneuronas12, por esta razón se lo ha utilizado para valorar la estructura laminar de la corteza cerebral en modelos experimentales de trauma13 y en cultivos organotípicos en humano14.
La IR para NeuN disminuye bajo condiciones patológicas experimentales tales como isquemia-hipoxia, excitotoxicidad mediada por órganofosforados, en enfermedades inflamatorias y en modelos de trauma craneano9-11,15. En el caso de tejido humano se ha encontrado reducción de la expresión de NeuN en tejido obtenido de hipocampo de individuos epilépticos con antecedentes de TCE severo, observándose pérdida neuronal en 94% de los casos de un total de 11 evaluados16.
En el presente trabajo se encontró una disminución de la IR para NeuN en todas las muestras de corteza cerebral analizadas de los pacientes con trauma. El evento es heterogéneo de tal manera que se observan sectores en los cuales hay pérdida general de la IR, sectores de pérdida en parches y otros sectores en los cuales la IR es aparentemente normal, siendo visible en estas la laminaridad cortical.
El evento más notorio en todos los casos es el inicio de la pérdida de IR por las capas supragranulares, particularmente la capa III, evento que va involucrando a las capas infragranulares, hasta constituirse en un parche que compromete un sector de láminas tanto supra como infragranulares. En ninguno de los casos observados el proceso se inició por alteraciones en las capas infragranulares; lo que indica que posiblemente exista algún tipo de vulnerabilidad selectiva hacia la capa III. El compromiso de la lámina III, principal fuente de las conexiones cortico-corticales, daría cuenta de las alteraciones neurológicas y psiquiátricas tales como depresión, demencia y alteraciones de la memoria, que presentan los pacientes que han sufrido un TCE. Este resultado estaría apoyado por los hallazgos de Ikomonovic et al.7, en individuos que presentaron incrementos en los depósitos de APP posterior a TCE, lo cual ha permitido proponer que posterior a TCE se acelera en 10 años la aparición de Alzheimer. En los sectores en los cuales la laminaridad está conservada, se aprecia a altas magnificaciones alteraciones sutiles de la IR para el marcador NeuN, lo cual indicaría que estos sectores podrían estar ingresando a un proceso de neurodegeneración o de alteración metabólica y no de pérdida neuronal, tal como lo han informado algunos autores en la literatura9,17.
Diversos autores consideran que la reducción de la IR para NeuN se asocia con pérdida neuronal15,16. Sin embargo, otros autores afirman que la pérdida de la IR es un evento transitorio, que reflejaría sólo un cambio en el estado de fosforilación de la proteína NeuN y por tanto el proceso podría ser reversible17. Se ha demostrado, en un modelo de isquemia cerebral moderada en ratón, disminución de la IR para NeuN sin cambios en los niveles de la proteína, lo cual sugiere pérdida transitoria de la antigenicidad y no pérdida neuronal9. Sin embargo, más recientemente en un modelo de envenenamiento en ratones con órganofosforados se indica que la pérdida de la IR de NeuN se puede correlacionar con muerte celular si se evalúa en tiempos más prolongados de sobrevivencia (72 horas posterior a la injuria)10.
En el presente estudio es difícil correlacionar si la pérdida de la IR es transitoria o permanente, porque no encontramos diferencias entre los sujetos, a pesar de que las muestras se obtuvieron en un tiempo quirúrgico promedio de 57 horas post-trauma. Cuando se evaluó en secciones consecutivas la expresión de NeuN y MAP2 colocando en registros las secciones, la ausencia de NeuN se acompañaba de pérdida de la expresión de MAP2, mientras que la presencia de NeuN puede o no estar acompañada de la presencia de MAP2. De acuerdo con lo anterior, es indispensable la valoración simultánea de la expresión de NeuN y MAP2 para definir la magnitud de la lesión y las zonas en riesgo de degeneración. Es importante anotar que la pérdida de la IR para NeuN, se ha señalado como un predictor de muerte tardía10. Este hecho se refuerza con la observación de que las células que pierden la IR para NeuN presentan fragmentación del ADN revelado por la técnica de TUNEL.
El único antecedente de pérdida de la IR de NeuN en seres humanos asociada con trauma se informó a partir de sujetos cuya cirugía para epilepsia del lóbulo temporal ocurrió 19 años después del trauma16. En el presente estudio la pérdida de la IR para NeuN fue consistente a lo largo de las diferentes láminas corticales, pero este hecho fue más marcado en la lámina III. Esto podría deberse a un efecto adicional de axotomía considerando que estas neuronas generan axones largos, lo cual las haría más vulnerables al estiramiento generado por las fuerzas de aceleración y desaceleración generadas durante el evento traumático. Sin embargo, sólo existe evidencia parcial de que la axotomía afecta la IR de las neuronas para NeuN.
MAP2 es una proteína altamente fosforilable en múltiples sitios de su estructura primaria in vivo, siendo sustrato para varias quinasas y fosfatasas. Su estado de fosforilación afecta la dinámica de los microtúbulos y su afinidad por estos, de tal manera que la IR de la proteína MAP2 puede variar de acuerdo con su estado de fosforilación8. El anticuerpo contra MAP2 permite observar con claridad las neuronas de la corteza cerebral tanto piramidales como no piramidales, con alguna preferencia por las primeras, además de la organización fascicular vertical de las dendritas apicales, sobre todo en láminas supragranulares, como se describió inicialmente en el ratón18 y luego en la neocorteza de humano19.
Estudios recientes involucran a los cambios en la IR de MAP2 como indicador de alteraciones neuronales, en particular por disrupción en la integridad del citoesqueleto8,11. Las reducciones cuantitativas y la pérdida de patrones normales de la IR a MAP2 se han relacionado con mecanismos de neurodegeneración producidos por isquemia (hipoxia, hipercapnia, acidosis, hipoglicemia), excitotoxicidad, estrés oxidativo y trauma experimental20. En el ser humano, se han informado disminuciones en la IR para MAP2 en corteza prefrontal dorsolateral post-mortem derivada de personas autistas, en pacientes que han tenido eventos hipóxico-isquémicos8,20 y en neocorteza de pacientes que sobrevivieron a isquemia cerebral19. En este último informe se encontró pérdida de la IR para MAP2 en las cortezas frontal (área 10), motora, temporal y visual, en especial en las capas II y III, con reducción de las ramificaciones secundarias y terciarias de los procesos dendríticos y aparición de pequeñas dilataciones en ellas, la pérdida de la IR para MAP2 la correlacionaron con la duración del período post-isquémico, siendo mayor en períodos más prolongados. Hasta el presente no se han hecho estas evaluaciones en tejido humano ex-vivo de sujetos que han sufrido TCE, por tanto los resultados del presente trabajo son inéditos y sólo comparables con las aproximaciones hechas en modelos animales o en tejido post-mortem derivado de otros síndromes.
Estos resultados muestran disminución generalizada de la IR para MAP2 tanto en los somas neuronales como en los procesos dendríticos, comprometiendo desde el inicio del proceso traumático tanto a las capas supra como las infragranulares. Algunos autores señalan que estos cambios se podrían deber a las alteraciones en los epítopes. Sin embargo, el anticuerpo 5F9, utilizado por nosotros reconoce a la proteína MAP2 independientemente del estado de fosforilación y de su estado de integridad, reconociendo aún a la proteína degradada. Por tanto, los cambios en la IR para MAP2 observados en este trabajo representan pérdida de esta proteína y por ende inestabilidad del citoesqueleto, dada probablemente por la activación de las calpaínas inducida por las concentraciones incrementadas de calcio que se generan en los eventos excitotóxicos que subyacen a los eventos traumáticos.
Las alteraciones en la organización de las proteínas del citoesqueleto podrían explicar los cambios en la dirección de los procesos apicales, la tortuosidad de los mismos, y la presencia de ensanchamientos. Estas alteraciones se pueden reflejar en alteraciones de los patrones normales de conectividad de la corteza cerebral debido a la desorganización de las espinas dendríticas, lo cual reduciría los contactos sinápticos, en especial los asociativos y comisurales que son los que se originan de las capas supragranulares, el compromiso observado en el citoesqueleto de las capas infragranulares podrían dar cuenta de alteraciones en las entradas talámicas tanto específicas como inespecíficas que no encontrarían el sustrato apropiado para su conectividad, y por tanto generaría una inadecuada regulación de las aferencias talámicas a la corteza cerebral.
Finalmente, las observaciones del presente trabajo aportan información inédita sobre el comportamiento neuronal utilizando la expresión de dos marcadores de injuria como NeuN y MAP2 en TCE de humanos, mostrando nítidamente la heterogeneidad de la lesión, la cual no sólo es macro sino microscópica, con preferencias laminares. Esta heterogeneidad permitió además observar que existen sectores necróticos adyacentes a sectores con compromiso exclusivo de capas supragranulares, de ambas y de otros con conservación de la organización laminar. Estos últimos sectores podrían representar zonas perilesionales o aún exofocales a la lesión, los cuales normalmente no es posible evaluar en el ser humano, pero que podrían indicar que a partir de estos sectores se pueden continuar emitiendo señales nocivas para el tejido sano, que agravaría en el tiempo la evolución del paciente y que explicaría las secuelas neurológicas, psicológicas y psiquiátricas de los mismos.
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento a Colciencias por la financiación del proyecto «Estudio cualitativo y cuantitativo del trauma craneoencefálico en humanos», Código 1106-04-16329, parte de cuyos resultados se muestran en el presente trabajo.
REFERENCIAS
1. Thurman DJ, Alverson C, Dunn KA, Guerrero J, Sniezek JE. Traumatic brain injury in the United State: A public health perspective. J Head Trauma Rehab. 1999; 14: 602-15. [ Links ]
2. Dubing AB, French BN, Rennick JM. Computed tomography in head trauma. Radiology. 1977; 122: 365-9. [ Links ]
3. Verger K, Serra-Grabulosa JM, Junqué C, Álvarez A, Bartrés-Faz D, Mercader JM. Estudio de las secuelas a largo plazo de los traumatismos craneoencefálicos: evaluación de la memoria declarativa y procedimental y de su sustrato neuroanatómico. Rev Neurol. 2001; 33: 30-4. [ Links ]
4. Guo Z, Cupples LA, Kurz A, Auerbach SH, Volicer L, Chui H, et al. Head injury and the risk of AD in the MIRAGE study. Neurology. 2000; 54: 1316-23. [ Links ]
5. Mueller CA, Schluesener, Conrad S, Meyermann R, Shwab JM. Lesional expression of a proinflamatory and antiangiogenic cytokine EMAP II confined to endothelium and microglia/macrophages during secondary damage following experimental traumatic brain injury. J Neuroimmunol. 2003; 135: 1-9. [ Links ]
6. McCracken E, Hunter AJ, Patel S, GrahamDI, Dewar D. Calpain activation and cytoskeletal protein breakdown in the corpus callosum of head injury. J Neurotraum. 1999; 16: 749-61. [ Links ]
7. Ikonomovic MD, Uryu K, Abrhamson EF, Ciallella JR, Trojanoski JQ, Lee VMY, et al. Alzheimer´s pathology in human temporal cortex surgically excised after severe injury. Exp Neurol. 2004; 190: 192-203. [ Links ]
8. Kuhn J, Meissner C, Ochmichen M. Microtubule-associated protein 2 (MAP2) a promising approach to diagnosis of forensic types of hypoxia-ischemia. Acta Neuropathol. 2005; 110: 579-86. [ Links ]
9. Unal-Cevik I, Kilinc M, Gursoy-Ozdemir Y, Gurer G, Dalkara T. Loss of NeuN immunoreactivity after cerebral ischemia does not indicate neuronal cell loss: a cautionary note. Brain Res. 2004; 1015: 169-74. [ Links ]
10. Collombet JM, Masqueliez C, Four E, Burckhart MF, Bernabe D, Baubichon D, et al. Early reduction of NeuN antigenicity induced by soman poisoning in mice can be used to predict delayed neuronal degeneration in the hippocampus. Neurosci Lett. 2006; 398: 337-42. [ Links ]
11. Macaya A. Muerte celular en la hipoxia-isquemia neonatal. Rev Neurol. 2000; 31: 784-9. [ Links ]
12. Mullen RJ, Buck CR, Smith AM. NeuN, a neuronal specific nuclear protein in vertebrates. Development. 1992; 116: 201-11. [ Links ]
13. Kobori N, Clifton GL, Dash P. Enhanced catecholamine synthesis in the prefrontal cortex after traumatic brain injury: Implications for prefrontal dysfunction. J Neurotraum. 2006; 23: 1094-102. [ Links ]
14. Verwer R, Hermens W, Dijkhuizen P. Cells in human postmortem brain tissue slices remain alive for several weeks in culture. FASEB J. 2002; 16: 54-60. [ Links ]
15. Robertson CL, Puskar A, Hoffman, Murphy AZ, Saraswati M, Fiskum G. Physiologic progesterone reduces mithocondrial dysfunction and hippocampal cell loss after traumatic brain injury in female rats. Exp Neurol. 2005; 197: 235-43. [ Links ]
16. Swartz BE, Houser CR, Tomiyasu U, Walsh GO, DeSalles A, Rich R, et al. Hippocampal cell loss in posttraumatic human epilepsy. Epilepsia. 2006; 47: 1373-82. [ Links ]
17. Lind D, Franjen S, Kappler J. Characterization of the neuronal marker NeuN as a multiply phosphorylated antigen with discrete subcellular localization. J Neurosci Res. 2005; 79: 295-302. [ Links ]
18. Escobar M, Pimienta H, Jacobson M, Crandall J, Kosik K, Caviness V. Architecture of apical dendrites in the murine neocortex: dual apical dendritic systems. Neuroscience 1986; 17: 975-89. [ Links ]
19. Akulinin VA, Dhlstrom A. Quantitative analysis of MAP2 immunoreactivity in human neocortex of three patients surviving after brain ischemia: Neurochem Res. 2002; 28: 373-8. [ Links ]
20. Haranashi Y, Kawara R, Fukuda S, Kiyoshima T, Morimoto Y, Matsumoto M, et al. Moderate hypothermia but not calpain inhibitor 2 attenuates the proteolysis of microtubule-associated protein 2 in the hippocampus following traumatic brain injury in rats: Eur J Anaesth. 2005; 22: 140-7. [ Links ]

