










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkColombia Médica
On-line version ISSN 1657-9534
Colomb. Med. vol.43 no.3 Cali July/Sept. 2012
Relacion entre genotipos del grupo sanguíneo Duffy e infección malárica en diferentes etnias de Choco-Colombia
Relationship between Duffy blood groups genotypes and malaria infection in different ethnic groups of Choco- Colombia
Gonzalez, Lina1, Vega, Jorge1*, Ramirez, Jose L1**, Bedoya, Gabriel1***, Carmona-Fonseca, Jaime1****, Maestre, Amanda1*****
1Universidad de Antioquia, Medellin, Colombia. E mail: lgonzalez@cib.org.co1*Universidad de Antioquia, Medellin, Colombia. E mail: javegapa@gmail.com
1**Universidad de Antioquia, Medellin, Colombia. E mail: genemed@medicina.udea.edu.co
1***Universidad de Antioquia, Medellin, Colombia. E mail: gbedoya@quimbaya.udea.edu.co
1****Universidad de Antioquia, Medellin, Colombia.E mail: jaimecarmonaf@hotmail.com
1*****Universidad de Antioquia, Medellin, Colombia.E mail: aemaestre@gmail.com
Received : 8 August 2011, Accepted : 17 August 2012
RESUMEN
Introducción:La condición homocigótica negativa del grupo sanguíneo Duffy (Fy-/Fy-) confiere resistencia natural a Plasmodium vivax. En este sentido, los estudios realizados en Colombia son escasos.Objetivo: describir la relación entre los genotipos Duffy en tres comunidades étnicas de La Italia (Chocó), y la malaria.
Metodología: estudio descriptivo, transversal con individuos sintomáticos o asintomáticos de malaria. Tamaño muestral por etnia: afrocolombiana 73; amerindia 74, mestiza171. La infección plasmodial se estudió por gota gruesa y el estado del gen Duffy por PCR y RFLP con el fin de identificar los cambios T-46C y A131G que originan los genotipos T/T, T/C , C/C y G/G, G/A, A/A.
Resultados: se encontró infección plasmodial en 17% con 62% por P. falciparum y 27% por P. vivax. Se halló relación entre genotipos Duffy y etnia (p=0,003). Los individuos con diplotipos C/C,A/A se infectaron exclusivamente con P. falciparum, mientras que otros diplotipos por las dos especies. En amerindios y mestizos la frecuencia del alelo T-46 fué 0,90-1,00, en afrocolombianos fué 0,50, al igual que el alelo C, con ausencia de heterocigotos. En el locus 131 la máxima frecuencia del alelo G fue 0,30 en amerindios y la máxima del alelo A fue 0,69 en afrocolombianos.
Conclusiones: En las poblaciones amerindia y mestiza estudiadas predomina el alelo T-46 (FY+), pero no predomina en ellas la infección por P. vivax. Ningún individuo FY- tuvo infección por P. vivax.
Palabras claves: Sistema del Grupo Sanguíneo Duffy, grupo étnico, malaria, Plasmodium falciparum, Plasmodium vivax, Genotipos, PCR-RFLP, Colombia.
ABSTRACT
Background:The negative homozygous condition for the Duffy blood group (Fy-/Fy-) confers natural resistance to Plasmodium vivax infection. In this direction, studies carried out in Colombia are scarce
Objective: To describe the relationship between Duffy genotypes in three ethnic communities in La Italia (Chocó) and malaria infection.
Methodology: a descriptive, cross-sectional study in symptomatic and asymptomatic malaria subjects. Sample size : Afro American, 73; Amerindian (Emberá), 74 and Mestizo, 171. Presence of Plasmodium infection was assessed by thick smear and the status of the Duffy gene by PCR and RFLP in order to identify the substitutions T-46C y A131G which origin the genotypes T/T, T/C , C/C y G/G, G/A, A/A.
Results: Infection by Plasmodium was detected in 17% with 62% due to P. falciparum and 27% to P. vivax. Duffy genotypes were significantly associated to ethnicity (p=0,003). Individuals with the C/C, A/A diplotype were exclusively infected by P. falciparum, whereas other diplotypes were infected with either species. In the Amerindian and Mestizo populations, the frequency of the T-46 allele was 0,90-1,00, among Afrocolombians this was 0,50, equal to the C allele and with absence of heterozygous At locus 131, the highest frequency of the G allele was 0,30 in Amerindians and the A allele was 0,69 in Afrocolombians.
Conclusions: In the Amerindian and mestizo populations studied, a predominance of the allele T-46 (FY+) was observed, but P. vivax was not the most common. Infection by P. vivax was out ruled in all FY- individuals.
Keywords: Duffy Blood-Group System, ethnic groups, malaria, Plasmodium falciparum, Plasmodium vivax, Genotypes, PCR-RFLP, Colombia.
INTRODUCCIÓN
En los humanos, la presencia de algunos polimorfismos en genes o en proteínas que se expresan en la membrana plasmática de los glóbulos rojos se relaciona con resistencia innata a la infección malárica1. La anemia de células falciformes, la deficiencia de glucosa 6 fosfato deshidrogenasa (G6PD) y las talasemias α y β son el resultado de polimorfismos que confieren protección parcial contra la infección por Plasmodium falciparum1. Por otra parte, el fenotipo Duffy negativo (FY-) confiere resistencia a la infección sanguínea por P. vivax, es decir, la ausencia de la proteína no permite la unión del merozoíto a la superficie de la membrana del eritrocito1. El polimorfismo eritroide Fya y Fyb de la proteína Duffy, resulta de la sustitución de un aminoácido, glicina por asparagina en la posición 42 del dominio extracelular del polipéptido, mientras que la ausencia de expresión de Fy resulta de la sustitución de una citosina por una timina en la región promotora del gen. Estos cambios generan cuatro principales fenotipos: Fya+b+, Fya+b-, Fya-b+ y Fya-b- 2.
A pesar de la importante evidencia clínica y epidemiológica en relación con la protección contra P. vivax en individuos Duffy negativo, varios estudios han sugerido presencia de infección por esta especie en ausencia de expresión de la proteína 3, 4, 5, 6, 7, 8. Lo cual podría encontrar explicación en el hecho de que Fyb es el alelo ancestral y que la expresión de Fya resulta en una disminución en la eficiencia de la unión de P. vivax y, por lo tanto, en disminución de la susceptibilidad sin protección completa contra la invasión3, 9, 10. Fenómeno que podría resultar, hipotéticamente, en parasitemia asintomática.
Se ha postulado que P. vivax ha ejercido presión a favor de la selección del alelo C (FYBnull), ya que en África se confirma una fuerte correlación entre la ausencia o baja endemicidad de la malaria por P. vivax y la alta prevalencia del fenotipo Duffy negativo en población nativa1, 11, 12, 13, 14. En Colombia son pocos los estudios con el grupo sanguíneo Duffy como marcador genético para identificar individuos afrocolombianos15, aún más pocos son los estudios en los que se relacionan los diferentes fenotipos y genotipos del gen Duffy con malaria. En 1994, Montoya y cols16, encontraron infección por P. vivax en sujetos FY- (evaluado por aglutinación) con frecuencia de 8.9%. Recientemente, el análisis de la especie infectante en una población afroamericana de la región de la Costa Pacífica, ha confirmado la predominancia de la infección por P. falciparum, hecho que ha sido atribuido a la ausencia del receptor Duffy en este grupo étnico17. Otros investigadores han demostrado una alta frecuencia de individuos FY- en habitantes de esta región del país18
La presente investigación tuvo como objetivo conocer las características moleculares del gen Duffy en tres comunidades étnicas (negra, amerindia y mestiza) habitantes de una misma zona ecoepidemiológica de alta endemicidad malárica y explorar la relación con la presencia de infección por P. vivax y P. falciparum. Se planteó la inclusión de individuos tanto sintomáticos como asintomáticos de malaria como una primera aproximación a identificar una relación con los diferentes genotipos Duffy y la posibilidad de protección parcial contra la infección por P. vivax.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en el corregimiento La Italia, municipio San José del Palmar (Chocó) (4°51’ LN, 76°18’ LO). La Italia cuenta con una población de cuatro mil personas y tiene composición étnica de 2500 individuos afrocolombianos, 1000 mestizos (resultantes del cruce de dos etnias) y 500 amerindios (Emberá, residentes en su mayoría en el resguardo Campoalegre).
Población de estudio y muestra: Se aplicó un diseño descriptivo, transversal y prospectivo, para conocer la prevalencia del antígeno Duffy en los tres grupos étnicos y su asociación con las distintas especies infectantes de Plasmodium. Con base en un nivel de confianza de 95%, error de muestreo de 5%, prevalencia del antígeno Duffy negativo 0,95 para afrocolombianos, 0,05 en indígenas y 0,16 en mestizos, se calcularon muestras para cada grupo, así: 71 Afrocolombianos, 64 Amerindios, 171 Mestizos15, 19, 20, 21, 22. Se incluyeron, de manera secuencial, sujetos voluntarios mayores de 1 año de edad que se manifestaran ser Afroamericanos por ''pertenencia cultural a una comunidad negra por autoreconocimiento'', Amerindios si ''se identifica o reconoce a sí mismo como perteneciente a un grupo étnico determinado, con tradición cultural anterior a la Conquista y que vive en comunidad, es decir, en el territorio que ocupa su comunidad o grupo'' y Mestizos si ''se reconoce a sí mismo como proveniente de cruce de europeos, Africanos e indígenas a partir de 1492 en la América hispana''23. Como único criterio de exclusión se consideró el retiro del consentimiento informado.
Evaluación de clínica y parasitológica: Los voluntarios fueron evaluados clínicamente por médico con experiencia en el diagnóstico de malaria. Se registraron los síntomas y signos asociados con malaria, de acuerdo con el protocolo de la Organización Mundial de la Salud para la definición de caso clínico probable. Se consideró como asintomático aquel individuo que, en las últimas dos semanas, tuviera ausencia síntomas y signos clínicos de malaria al momento del examen. El diagnóstico de infección por Plasmodium se realizó en gota gruesa y extendido delgado, preparados con sangre capilar obtenida por punción del pulpejo de un dedo. Las placas fueron teñidas con Giemsa o Field, y examinadas por personal experto. La parasitemia se calculó con base en una constante de 8.000 leucocitos/mm. Las muestras se consideraron negativas después de examinar 200 campos microscópicos con objetivo de 100X.
Genotipificación del antígeno Duffy. El estado del gen Duffy fue determinado en todos los individuos, sin embargo el fenotipo Duffy no fue analizado. A partir de glóbulos blancos de sangre periférica o de células epiteliales de mucosa oral, se obtuvo ADN mediante ''Salting out'' o fenol-cloroformo. El ADN obtenido se precipitó y se almacenó a -20°C hasta su utilización.
La identificación de los diferentes alelos del gen Duffy se realizó con base en metodología publicada por Tournamille y cols.2, que permite evaluar el estado de las posiciones -46 y 131 del gen con el fin de identificar las substituciones T-46C y A131G responsables de los diferentes alelos. Brevemente, se aplicaron cebadores específicos para amplificar un fragmento de 1.0 kb del gen Duffy, el producto de esta primera reacción se utilizó como molde para (a) una segunda reacción de amplificación con nuevos cebadores para discriminar entre Fy+ y Fy- , y (b) en caso de Fy+ se realizó una segunda amplificación con cebadores específicos para definir el polimorfismo del genotipo que origina los fenotipos Fya+Fyb+, Fya+Fyb- y Fya-Fyb+. Los productos de las segundas amplificaciones fueron sometidos a análisis de los fragmentos de restricción (RFLP de la sigla en inglés: restriction fragment length polymorphism)2, con Sty 1 y BshNI (Fermentas®).
Los productos de digestión se observaron en gel de poliacrilamida al 15%, teñido con bromuro de etidio. El tamaño de los fragmentos se evaluó con el programa Quantity One (Bio-Rad,) así: bandas de 62, 77 y 82 pares de bases (pb) corresponden a Duffy positivo (Fy +), bandas de 12, 62, 65 y 82 pb se interpretan como Duffy negativo (Fy -), bandas de 86 y 73 pb son Fya+b- , bandas de 159, 86 y 73 pb son Fy a+b+ y una banda de 159pb es Fy a-b+ de. Los controles positivos y negativos correspondían a muestras de tipificación ya estandarizadas en el laboratorio del Grupo Salud y Comunidad.
Análisis estadístico: Se analizaron variables cualitativas y cuantitativas. Las cualitativas fueron analizadas mediante la prueba ji (chi) cuadrada, con el programa EpiInfo 6.0. La obtención de frecuencias genotípicas y alélicas y las pruebas de neutralidad y estructuración genética se hicieron con ayuda del programa ''Genepop'' versión 3.1. La generación de haplotipos más probables se hizo con el programa ''Arlequin'' versión 2.000; la estructura poblacional se estimó con la distancia genética de Cavalli-Sforza, mediante el algoritmo de Neighbor-Joining (UPGMA versión 3.572c).
Consideraciones éticas: La investigación cumplió con los requisitos de la Resolución 008430 de 1993 del Ministerio de Salud de Colombia, el Código de Nuremberg 1947 y la Declaración de Helsinki 1987 y fue avalada por el Comité de Ética del Instituto de Investigaciones Médicas, Facultad de Medicina, Universidad de Antioquia. El diseño de la propuesta tuvo la aprobación verbal del Gobernador local de la comunidad indígena y cada individuo, o su guardián, que aceptó participar firmó o dejó su huella digital en el formulario de consentimiento informado. Aquellos individuos diagnosticados con malaria recibieron gratuitamente el tratamiento recomendado por las normas colombianas. La investigación se consideró de riesgo mínimo.
RESULTADOS
Descripción de la población del estudio: En total se estudiaron 320 voluntarios, con una relación hombre/mujer de 131/189. La distribución por etnia fue 73 afrocolombianos (23%), 74 amerindios (23%) y 173 (54%) mestizos. La tabla 1 detalla las características generales de la población.
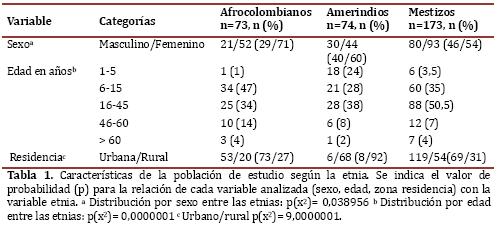
Frecuencia de infección por Plasmodium. Con la gota gruesa se encontró una frecuencia de infección plasmodial de 17% (52/320). Entre quienes presentaron infección, la frecuencia por especie fué: 62% (32/52) P. falciparum, 27%(14/52) P. vivax, 11% ambas especies (infección mixta) (6/52). La parasitemia de P. falciparum en el grupo de infectados (sintomáticos y asintomáticos) tuvo un intervalo de confianza para el promedio de 8082 a 17190. Por etnias, la parasitemia promedio de P. falciparum estuvo alrededor de 6.500 a 8.000 anillos/βL en amerindios y mestizos, y fué 12.641 anillos/βL en los afrocolombianos, sin diferencia estadísticamente significativa (p[F]= 0,110649), quizás por el pequeño tamaño de los grupos y por su notoria variación que se refleja en las altas desviaciones típicas (Tabla 2). Las parasitemias por P. vivax o mixtas no se contabilizaron.
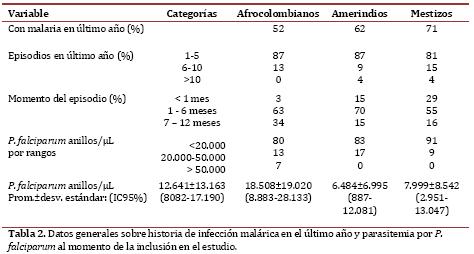
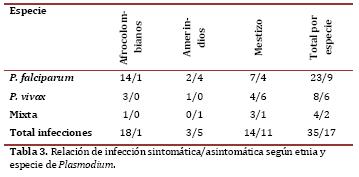
Entre los 52 casos de infección por Plasmodium, 33% se observaron en asintomáticos (17/52) (cuadro 3). No se encontró una relación significativa entre especie de Plasmodium y etnia.
En total, 65% (207/320) de la población informó antecedentes de infección malárica reciente, con promedio de 2 episodios en el último año; 124 individuos informaron algún episodio de malaria en los últimos 6 meses.
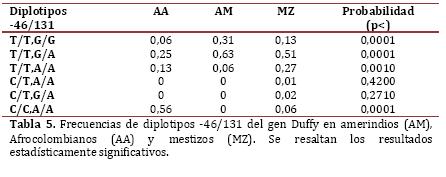
Genotipificación del grupo sanguíneo Duffy. La frecuencia del alelo T en el locus -46 en afrocolombianos fue de 0,50; en mestizos 0,91 y se encontró fijo en los amerindios. Los genotipos CC y TT mostraron frecuencia de 0,50 en afrocolombianos, en quienes hubo ausencia de heterocigotos; los mestizos presentaron los tres genotipos, con poca frecuencia de heterocigotos. En el locus 131, el alelo G se encontró con menor frecuencia en afrocolombianos (0,18) y la mayor en amerindios (0,62). En cuanto a la distribución genotípica, la menor frecuencia de heterocigóticos se halló en afrocolombianos (0,25) y la mayor en amerindios (0,63). En la Tabla 4 se detallan los datos de las frecuencias genotípicas y alélicas en cada una de las poblaciones para los loci evaluados.
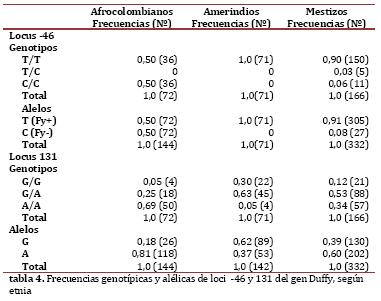
Según las pruebas de neutralidad de estos marcadores en cada una de las poblaciones, el locus -46 mostró diferencia altamente significativa en su frecuencia en afrocolombianos y en mestizos (p<0,00001) y en el locus 131 solamente mostró diferencia significativa en amerindios (p=0,00402).
La evaluación de la estructura genética de las poblaciones mediante Wright Fis, Fit y Fst para calcular endogamia y distancia genética, permitió observar que los valores de Fis para los loci -46 y 131 en afrocolombianos, muestran déficit de heterocigóticos, de la misma manera que el locus -46 para mestizos, mientras que en las poblaciones amerindia y mestiza existe un exceso de heterocigotos en el locus 131. Al evaluar cada locus según la población total, se observó que en el locus -46 existe un déficit de heterocigotos y en el locus 131 un exceso de ellos. Con respecto a los valores de Fst obtenidos (> 0,05), se observó que con el locus 131 se obtiene un grado de estructuración mediana, mientras que con locus -46 la estructuración es muy alta, > 0,25 entre las poblaciones. Con el fin de ratificar lo anterior, se compararon las diferencias genéticas mediante un árbol ''Neighbor Joining'' (NJ) y la distancia genética de Cavalli-Sforza. Se obtuvo alto grado de diferenciación genética entre las tres poblaciones. La mayor distancia se encontró entre afrocolombianos (AA) y amerindios (AM) (0,44), lo cual significa que el flujo génico entre estas dos etnias (mezcla) es el menor en la estructura de la población de La Italia. La menor distancia se encontró entre amerindios y mestizos (0,092), lo cual indica que la subpoblación mestiza proviene de mezcla con amerindios: la distancia de 0,28 entre AA y mestizos (MZ) es casi la mitad de la encontrada entre AA y AM.
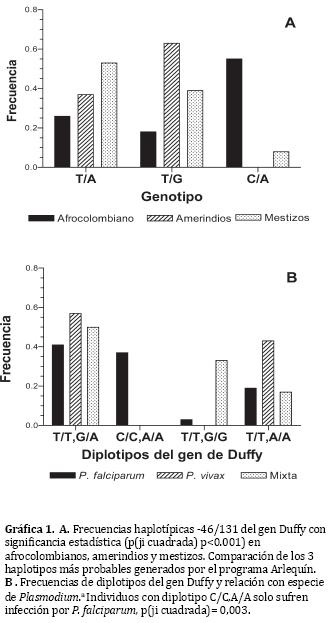
Adicionalmente, al analizar la asociación entre etnia y genotipo en los loci -46 y 131, se observó una diferencia altamente significativa (p<0,01) debido a que los afrocolombianos presentaron predominantemente el genotipo C-46C, contrario a lo ocurrido en amerindios y mestizos, quienes tuvieron predominantemente el genotipo G131A. Por otra parte, los amerindios exhibieron una frecuencia 1-5 veces mayor de G131G, mientras que los mestizos de 1- 3 veces mayor de A131A (gráfica 1).
Al comparar diplotipos y haplotipos (Tabla 5), se encontró diferencia estadísticamente significativa (p<0,001) en la distribución de las frecuencias en donde el diplotipo T/T, G/A predominó en amerindios y mestizos, el diplotipo C/C, A/A se encontró notablemente alto en afrocolombianos, muy poco frecuente en mestizos y ausente en amerindios, mientras C/T, A/A y C/T, G/A estaban ausentes o con muy baja frecuencia en las tres poblaciones.
Finalmente, la comparación de los haplotipos más probables permitió encontrar tres con diferencias estadísticamente significativas entre las poblaciones (p<0,0001) (Gráfica 1). El haplotipo TG presentó la mayor frecuencia en amerindios y la menor en afrocolombianos; mientras que el CA tuvo la frecuencia mayor en afrocolombianos, muy baja en mestizos y ausente en amerindios. El haplotipo TA predominó en mestizos.
Análisis de asociación de algunas variables. Al relacionar el promedio de episodios de malaria en el último año según diplotipo y etnia, no hubo diferencia estadísticamente significativa. Sin embargo, al analizar el número de episodios de malaria en el mismo período según etnia, se encontró diferencia significativa en los mestizos, quienes presentaron un promedio mayor de episodios de malaria en el último año (173 vs. 73 en afrocolombianos y 74 en amerindios; p<0,05).
También se encontró significancia estadística al relacionar diplotipo con etnia (p=0,003). Los individuos con diplotipo C/C, A/A solo sufrían infección por P. falciparum, mientras que los individuos con los otros diplotipos podían tener infección por P. vivax o P. falciparum (gráfica 2). No se observaron diferencias estadísticamente significativas al relacionar sexo y edad con el resultado positivo o negativo de la gota gruesa (p=0,054 y p=0,092).
DISCUSIÓN
El presente estudio encontró que más de una tercera parte de las muestras con parasitemia postiva provenían de individuos asintomáticos para malaria. Estudios realizados en Colombia han informado valores de infección asintomática de 0% en escolares de Quibdó17 y de 14,6 % en población de Tierralta, Cordoba (2-76 años)24. La diversidad en las poblaciones así como la edad y la composición étnica25 podrían influenciar la presentación de parasitemia asintomática. No obstante, al menos el presente estudio, no encontró ninguna asociación significativa entre edad y frecuencia de infección malárica en las diferentes etnias, por lo que la relevancia de la interacción entre la edad-etnia-infección asintomática, debe ser más ampliamente explorada.
Se observó una predominancia del alelo C en el locus -46 (Fy-) en afrocolombianos, pero en menor frecuencia a las informadas en poblaciones africanas11, 26. Otros investigadores han indicado bajas frecuencias de este alelo en poblaciones colombianas mestizas15. Esto probablemente se deba a que la población afroamericana ha sufrido un nivel significativo de mestizaje. Análisis adicionales de los genotipos del locus -46 confirmó que las etnias afrocolombiana y mestiza no están en equilibrio de Hardy Weinberg, mientras que en la etnia afroamericana existe un exceso de homocigotos T/T y C/C, lo que desfavorece la hipótesis de que el genotipo C/C se esté seleccionando por P. vivax, como sí se ha observado en otras poblaciones del Chocó27.
La frecuencia del alelo G en el locus 131 predomina en amerindios, mientras que la frecuencia del alelo A predomina en afrocolombianos y en mestizos. Datos similares se han reportado en individuos de Norteamérica (caucásico- americanos), afroamericanos, turcos y colombianos, en los que predomina el alelo A131, en contraste con individuos asio-americanos e hispanoamericanos en general, en los cuales predomina el alelo G15, 26.
La relación encontrada entre etnia y genotipo Duffy en la posición 131 probablemente se deba a que amerindios y mestizos portan predominantemente el genotipo GA, pero los individuos afrocolombianos el AA. Estos datos difieren de estudios hechos en distintos grupos étnicos de Papua, Nueva Guinea, en los que se informa que la gran mayoría de los individuos son GG. Las etnias afrocolombiana y mestiza muestran equilibrio de Hardy Weinberg en el locus 131, equilibrio que también se han encontrado en otras poblaciones en Turquía28. Por el contrario, el equilibrio de Hardy Weinberg no se detectó en amerindios como consecuencia del exceso de heterocigotos.
Los hallazgos de desequilibrio de Hardy Weinberg en el locus -46 en población afrocolombiana y mestiza, y en el locus 131 de la población amerindia podría explicarse por la presencia de endogamia en cada una de las tres poblaciones y por su historia de migración. Los valores de Fis para los loci -46 y 131 en afrocolombianos, muestran un déficit de heterocigóticos, lo que podría explicarse al revisar históricamente la migración de esta población afrocolombiana hacia esta localidad: según comunicación oral recolectados en el sitio. De acuerdo con ello, la población afrocolombiana proviene, a su vez, de dos localidades, unos individuos del sur del Chocó y otros del noroccidente. Este fenómeno podría explicar la ausencia de heterocigotos en La Italia, que es de reciente fundación (año 1.927). De manera similar, el desequilibrio que existe en el locus -46 en la población mestiza, en la cual también se observa un déficit de heterocigotos, podría explicarse por la colonización de poblaciones antioqueñas y, en los últimos años, los desplazamientos del Caquetá, Putumayo y Amazonas. En contraste, en el locus 131 sólo existe desequilibrio en la población amerindia, debido a un exceso de heterocigotos.
Los altos valores de Fst están demostrando que hay estructuración por origen étnico entre las tres poblaciones, por lo tanto, las poblaciones se diferencian genéticamente una de otra utilizando estos marcadores. Es preciso destacar que los valores de Fst más altos se encuentran en el locus -46, es decir, este marcador es el que señala la mayor diferencia en el origen étnico, lo cual corrobora su utilidad como marcador de etnia en esta poblaciones. Adicionalmente, al evaluar la estructura poblacional mediante la distancia genética de Cavalli-Sforza, se confirma, en general, un alto grado de diferenciación genética entre las poblaciones de las tres etnias de una misma área geográfica. La mayor diferenciación está entre amerindios y afrocolombianos y la distancia genética menor es la que se presenta entre amerindios y mestizos. Adicionalmente, la distancia que existe entre amerindios y mestizos es menor que la que se observa entre amerindios y afrocolombianos; esto significa que los mestizos tienen un poco más de componente amerindio que los afrocolombianos.
Los anteriores resultados indican que aunque estas poblaciones están ubicadas en una misma área geográfica, hay persistencia de la estructuración étnica, que reflejan un alto grado de endogamia y discriminación entre las poblaciones.
Este es el primer estudio colombiano sobre malaria y grupos sanguíneos Duffy en el que se hacen comparaciones de los diplotipos -46/131. Dado que hay evidencia de que el genotipo C-46C y A131A protegen contra la infección por P. vivax, es probable que el haplotipo C/A sea el protector en las poblaciones estudiadas, puesto que presentó la mayor frecuencia en la población afrocolombiana.
Al asociar diplotipos Duffy con etnia y especie de Plasmodium infectante, se observó que los individuos con diplotipo C/C, A/A sólo sufren infección por P. falciparum, mientras que los individuos con los otros diplotipos pueden ser infectados por P. vivax o P. falciparum. Esto permite confirmar que este sea el diplotipo protector contra P. vivax. Sin embargo, la baja frecuencia de infecciones por esta especie impidió obtener resultados significativos en relación con la asociación entre especie y etnia, aunque P. vivax predominó en mestizos y P. falciparum en afrocolombianos. Estos resultados difieren de los encontrados por Montoya y cols, quienes reportaron un predominio de infección por P. vivax en amerindios emberá y P. falciparum en indígenas kunas16
los resultados del genotipo Duffy, y de esta investigación confirman que, al igual que en la región de la amazonía brazilera29, no se encuentra ningún individuo FY- con infección por P. vivax. Lo que contradice lo informado en Colombia en 1.99416 y lo más reciente en varios países de Africa5, 6. Una explicación para la diferencia en resultados en el contexto colombiano puede radicar en la limitación de la técnica serológica para detectar el fenotipo débil FYx, así como en la existencia de diferentes variantes genéticas de P. vivax, fenómenos que no se exploraron en el presente estudio. Un futuro trabajo en Colombia debe aclarar, las asociaciones entre infección por diversas variantes de P. vivax y los genotipos Duffy, mediante la aplicación de técnicas basadas en la amplificación de ácidos nucléicos por reacción en cadena de la polimerasa además de la evaluación del tamaño de fragmentos después de digestión con enzimas de restricción (RFLP) o secuenciamiento de ácidos nucleicos.
REFERENCIAS
1. Rowe JA, Opi DH, Williams TN. Blood groups and malaria: fresh insights into pathogenesis and identification of targets for intervention. Curr Opin Hematol. 2009;16(6):480-7. [ Links ]
2. Tournamille C, Le Van Kim C, Gane P, Cartron JP, Colin Y. Molecular basis and PCR-DNA typing of the Fya/fyb blood group polymorphism. Hum Genet. 1995;95(4):407-10. [ Links ]
3. Cavasini CE, de Mattos LC, Couto AA, Couto VS, Gollino Y, Moretti LJ, et al. Duffy blood group gene polymorphisms among malaria vivax patients in four areas of the Brazilian Amazon region. Malar J. 2007:167. [ Links ]
4. Cavasini CE, Mattos LC, Couto AA, Bonini-Domingos CR, Valencia SH, Neiras WC, et al. Plasmodium vivax infection among Duffy antigen-negative individuals from the Brazilian Amazon region: an exception? Trans R Soc Trop Med Hyg. 2007:1042-4. [ Links ]
5. Menard D, Barnadas C, Bouchier C, Henry-Halldin C, Gray LR, Ratsimbasoa A, et al. Plasmodium vivax clinical malaria is commonly observed in Duffy-negative Malagasy people. Proc Natl Acad Sci. 2010:5967-71. [ Links ]
6. Mendes C, Dias F, Figueiredo J, Mora VG, Cano J, de Sousa B, et al. Duffy negative antigen is no longer a barrier to Plasmodium vivax--molecular evidences from the African West Coast (Angola and Equatorial Guinea). PLoS Negl Trop Dis. 2011:e1192. [ Links ]
7. Ryan JR, Stoute JA, Amon J, Dunton RF, Mtalib R, Koros J, et al. Evidence for transmission of Plasmodium vivax among a duffy antigen negative population in Western Kenya. Am J Trop Med Hyg. 2006:575-81. [ Links ]
8. Wurtz N, Mint Lekweiry K, Bogreau H, Pradines B, Rogier C, Ould Mohamed Salem Boukhary A, et al. Vivax malaria in Mauritania includes infection of a Duffy-negative individual. Malar J. 2011:336. [ Links ]
9. Li J, Iwamoto S, Sugimoto N, Okuda H, Kajii E. Dinucleotide repeat in the 3' flanking region provides a clue to the molecular evolution of the Duffy gene. Hum Genet. 1997;99(5):573-7. [ Links ]
10. Tournamille C, Blancher A, Le Van Kim C, Gane P, Apoil PA, Nakamoto W, et al. Sequence, evolution and ligand binding properties of mammalian Duffy antigen/receptor for chemokines. Immunogenetics. 2004;55(10):682-94. [ Links ]
11. Welch SG, McGregor IA, Williams K. The Duffy blood group and malaria prevalence in Gambian West Africans. Trans R Soc Trop Med Hyg. 1977;71(4):295-6. [ Links ]
12. Miller LH, Mason SJ, Clyde DF, McGinniss MH. The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med. 1976;295(6):302-4. [ Links ]
13. Mathews HM, Armstrong JC. Duffy blood types and vivax malaria in Ethiopia. Am J Trop Med Hyg. 1981;30(2):299-303. [ Links ]
14. Howes RE, Patil AP, Piel FB, Nyangiri OA, Kabaria CW, Gething PW, et al. The global distribution of the Duffy blood group. Nat Commun. 2011 Macmillan Publishers Limited; 2011:266. [ Links ]
15. Sandoval C, De la Hoz A, Yunis E, 1993. Estructura genética de la Población Colombiana. Revista Facultad de Medicina Universidad Nacional de Colombia 41:3-14. [ Links ]
16. Montoya F, Restrepo M, Montoya AE, Rojas W. Blood groups and malaria. Rev Inst Med Trop Sao Paulo. 1994;36(1):33-8. [ Links ]
17. Ochoa J, Osorio L. Epidemiology of urban malaria in Quibdo, Choco. Biomedica. 2006;26(2):278-85. [ Links ]
18. Herrera S, Gomez A, Vera O, Vergara J, Valderrama-Aguirre A, Maestre A, et al. Antibody response to Plasmodium vivax antigens in Fy-negative individuals from the Colombian Pacific coast. Am J Trop Med Hyg. 2005:44-9. [ Links ]
19. Singh N, Shukla MM, Uniyal VP, Sharma VP. ABO blood groups among malaria cases from district Mandla, Madhya Pradesh. Indian J Malariol. 1995;32(2):59-63. [ Links ]
20. Fischer PR, Boone P. Short report: severe malaria associated with blood group. Am J Trop Med Hyg. 1998;58(1):122-3. [ Links ]
21. Martinez Bencardino C. Estadística y muestreo; 2000. [ Links ]
22. Hadley TJ, Peiper SC. From malaria to chemokine receptor: the emerging physiologic role of the Duffy blood group antigen. Blood. 1997;89(9):3077-91. [ Links ]
23. DANE. Grupos étnicos de Colombia en el censo de 1993. Bogotá: Departamento Administrativo Nacional de Estadística. 1998. [ Links ]
24. Cucunuba ZM, Guerra AP, Rahirant SJ, Rivera JA, Cortes LJ, Nicholls RS. Asymptomatic Plasmodium spp. infection in Tierralta, Colombia. Mem Inst Oswaldo Cruz. 2008;103(7):668-73. [ Links ]
25. Njama-Meya D, Kamya MR, Dorsey G. Asymptomatic parasitaemia as a risk factor for symptomatic malaria in a cohort of Ugandan children. Trop Med Int Health. England; 2004:862-8. [ Links ]
26. Chiaroni J, Touinssi M, Frassati C, Degioanni A, Gibert M, Reviron D, et al. Genetic characterization of the population of Grande Comore Island (Njazidja) according to major blood groups. Hum Biol. 2004;76(4):527-41. [ Links ]
27. Bravo ML, Valenzuela CY, Arcos-Burgos OM. Polymorphisms and phyletic relationships of the Paisa community from Antioquia (Colombia). Gene Geogr. 1996;10(1):11-7. [ Links ]
28. Dönbak L RE. Frequency of blood group antigens and corresponding alleles in Mediterranean region of Turkey. 2002;5:326-29. [ Links ]
29. Weber SS, Pedro W, Martins T, Sotero A. Frecuencia alélica del Sistema del Grupo Sanguíneo Duffy en individuos de una población de la Amazonía brasilena y su relación con la infección por Plasmodium vivax. Revista Pan-Amazônica de Saúde. 2010;1(2):149-. [ Links ]
