










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiosalud
Print version ISSN 1657-9550
Biosalud vol.13 no.2 Manizales July/Dec. 2014
IMPACT OF HEAT STRESS ON REPRODUCTIVE ACTIVITY IN CATTLE AND CONSIDERATIONS TO MITIGATE ITS EFFECTS ON REPRODUCTION
Faider A. Castaño1
Clara C. Rugeles P.2
César A. Betancur H.3
Camilo J. Ramirez-López4
1 Universidade Federal de Viçosa, MG, Brasil.
2 Laboratorio de Andrología. Facultad de Medicina Veterinaria y Zootecnia, Universidad de Córdoba, Montería, Colombia. Autor para correspondencia, correo electrónico: crugeles@correo.unicordoba.edu.co
3 Facultad de Medicina Veterinaria y Zootecnia, Universidad de Córdoba, Montería, Colombia.
4 Laboratorio de Andrología. Facultad de Medicina Veterinaria y Zootecnia, Universidad de Córdoba, Montería, Colombia.
Los mamíferos, al igual que todos los organismos vivos, deben cubrir sus requisitos nutricionales para lograr la sobrevivencia y evolución de sus especies, para lo cual requieren unas condiciones que les garanticen su bienestar, de ello depende el aprovechamiento de los nutrientes. Las fases del desarrollo que implican procesos reproductivos son de altos requerimientos nutricionales y elevada actividad metabólica, por lo que pueden verse afectadas por condiciones climáticas severas. El estrés calórico produce efectos negativos sobre la gametogénesis, los pulsos hormonales, la expresión del celo, el ovocito, la fecundación, la implantación, el desarrollo embrionario y el metabolismo basal, inclusive, en la respuesta a la alteración funcional del eje neuroendocrino. En los países tropicales es importante adoptar algunas estrategias para reducir el choque térmico en los animales, para mejorar su rendimiento productivo y reproductivo. La opción de primera mano es utilizar animales que han evolucionado en condiciones climáticas similares y que, por ende, han alcanzado cierto grado de adaptación. Otra medida, es el uso de sistemas que ofrezcan a los animales menor exposición directa a los rayos solares, bien sea a través de sistemas silvopastoriles o por el uso de polisombras; o bien el uso de sistemas de enfriamiento directo con rociadores de agua o ventiladores que han mostrado mejorar las tasas de concepción. Actualmente, con la implementación de biotécnicas reproductivas se está apuntando a suplir fallas de tipo hormonal, con técnicas como la IATF (sincronización de la ovulación con inseminación a tiempo fijo). Con la presente revisión se pretende estudiar los efectos del estrés térmico sobre la actividad reproductiva de la vaca en el trópico, y considerar la implementación de algunas medidas que ayuden a mitigar el impacto negativo de las condiciones climáticas del trópico.
Palabras clave: estrés calórico, fertilidad, hembras, hormonas, biotecnología reproductiva.
ABSTRACTMammalian like all living organisms must cover their nutritional requirements in order to achieve both survival and evolution of their species for which reason the conditions that ensure their welfare, depend from the nutrient exploitation. The phases of development involving reproductive processes imply high nutritional requirements and high metabolic activity, which may be affected by severe weather. The heat stress has adverse effects on gametogenesis, hormonal pulses, estrus expression, the oocyte, fertilization, implantation, embryonic development and basal metabolism, even in the response to functional alteration of the neuroendocrine axis. In tropical countries is important to adopt some strategies to reduce the thermal shock in animals, to improve their productive and reproductive performance. The first option is to use animals that have evolved in similar climatic conditions and thus have reached a certain degree of adaptation. Another measure is the use of systems that offer animals less direct exposure to sunlight, either through silvopastoral systems, the use of shading coefficient, the use of direct cooling systems with water sprinklers or fans that have shown to improve conception rates. Currently the implementation of reproductive biotechnology is aiming to supply hormone-like failures with techniques such as FTAI (fixed time artificial insemination). The present review is aimed to study the effects of heat stress over cow reproductive activity in the tropics and to consider implementing some measures intended to mitigate the negative impact of climatic conditions in the tropic.
Key words: fertility, females, heat stress, hormones, reproductive biotechnology.
Los bovinos, al igual que todos los mamíferos, son animales endotérmicos; es decir, organismos que a pesar de las fluctuaciones en la temperatura ambiental son capaces de mantener relativamente constante la temperatura corporal. Esta homeostasis es esencial para una multitud de reacciones bioquímicas y procesos fisiológicos asociados con el normal metabolismo; pero esta capacidad se ve amenazada por fuerzas adversas intrínsecas o extrínsecas, los estresores. Un estresor es cualquier cambio medioambiental que rompe la homeostasis (1).
El estado de dishomeostasis debido a respuestas adaptativas prolongadas, en las cuales el animal sobrevive pero sufre consecuencias adversas ha sido denominado alostasis; en esta condición, los sistemas homeostáticos responden a situaciones de agresión como pueden ser las temperaturas extremas. A diferencia de los sistemas homeostáticos, que mantienen las variables fisiológicas dentro de rangos estrechos, los sistemas alostáticos comprenden tanto mecanismos para desencadenar la respuesta con activación de una compleja vía adaptativa como para detener la respuesta cuando la amenaza ha desaparecido (2).
El estrés calórico es la fuerza ejercida por los componentes del ambiente térmico sobre el organismo, causando en él una reacción fisiológica proporcional a la intensidad de la fuerza aplicada y a la capacidad del organismo en compensar las desviaciones causadas por esta fuerza (3); por lo tanto, el estrés térmico afecta todo el sistema neuroendocrino, desencadenando eventos fisiológicos y comportamentales con el objetivo de mantener la homeostasis en detrimento de los procesos productivos y reproductivos (4).
La presente revisión se enfoca en la neuroendocrinología del estrés calórico y sus efectos en el metabolismo, en el balance energético y en la actividad reproductiva en los bovinos.
Eje hipotalámico-hipofisario-adrenal
El organismo animal reacciona al estrés generando un estado de respuestas adaptativas y de comportamiento, las cuales pueden tener consecuencias adversas en funciones fisiológicas, tales como reproducción, metabolismo y producción del animal (5).
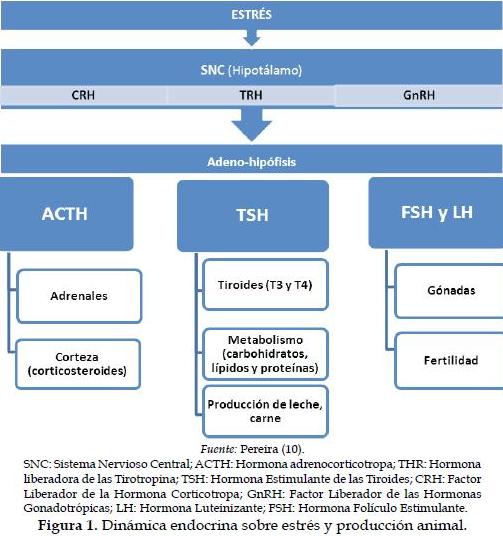
Desde el punto de vista hormonal, el cortisol parece ser la hormona más importante en las respuestas ante situaciones de estrés (6). El estímulo estresor determina la secreción de la CRH (factor liberador de la hormona corticotropa) por el hipotálamo (SNC), que estimula en la adenohipófisis la secreción de la hormona adrenocorticotropa (ACTH) lo que conlleva la liberación de hormonas producidas en las glándulas suprarrenales y adrenales (7). La CRH puede interferir en la función reproductiva, estimulando la producción de B-endorfinas de propiedades opioides y ACTH (8).
Las hormonas tiroideas también se presentan como potenciales indicadores de estrés, modificando su homeostasis ante condiciones ambientales adversas. La reacción más característica de las hormonas tiroideas ante eventos de estrés calórico es su disminución; inducen respuestas metabólicas que acarrean gastos energéticos y, consecuentemente, disminución de la producción de carne y leche (9).
Por último, el estrés disminuye la secreción de GnRH en el hipotálamo y la producción de FSH y LH en la adenohipófisis y, por lo tanto, de esteroides sexuales en las gónadas. Esto causa una disminución en los índices de fertilidad del ganado (Figura 1).
Efectos del estrés sobre el metabolismo
El estrés térmico (ET) afecta todo el sistema endócrino, desencadenando eventos fisiológicos y de comportamiento con el objetivo de mantener la homeostasis en detrimento de los procesos productivos y reproductivos (4). Además de sus efectos catabólicos directos, los glucocorticoides también antagonizan las acciones de la hormona del crecimiento (GH) y los esteroides sexuales en el catabolismo del tejido graso (lipólisis). La asociación entre estrés crónico, hipercortisolismo y manifestaciones relacionadas con el síndrome metabólico han sido reportadas en monos (11).
La porción medular de las glándulas adrenales secreta catecolaminas (adrenalina y noradrenalina), que responden a la excitación y proporcionan al organismo reacciones rápidas e inmediatas. Por su parte, la porción cortical de las adrenales secreta hormonas sexuales, y corticosteroides (gluicocorticoides, glucocorticosteroides y mineralocorticoides); hormonas esteroidales que ejercen funciones esenciales, como la regulación del metabolismo de los carbohidratos y el mantenimiento del equilibrio electrolítico (7).
En caso de estrés crónico, altos niveles de glucocorticoides, pueden ser perjudiciales para los tejidos y órganos ya que inhiben el crecimiento, ocasionando una hiperactividad crónica de las glándulas adrenales que causa atrofia del tejido linfoide y puede llevar al colapso de las defensas inmunitarias (7).
La tiroides es otra glándula que está involucrada con el estrés; el hipotálamo regula por medio de la TSH (Hormona Estimulante de las Tiroides) la producción de Hormonas Tiroideas (HT) en la tiroides. Las HT actúan en la regulación del metabolismo y la producción de calor, además de potenciar otras hormonas como las catecolaminas y la GH. De esta forma, en el estrés térmico, las concentraciones de las hormonas tiroideas disminuyen, llegando a generar hipertiroidismo o hipotiroidismo. El hipertiroidismo es común en la fase inicial del estrés, y el hipotiroidismo genera agotamiento y colapso del sistema endocrino cuando el estrés es crónico (7).
El balance energético provoca reducción de las concentraciones plasmáticas de insulina, glucosa, IGF-I, aumento de GH y ácidos grasos no esterificados. Estos actúan en el eje hipotálamo-hipófisis-gonadal (HHP) mediando los efectos inhibitorios del balance energético negativo en la fertilidad posparto. Tanto el IGF-I como la glucosa estimulan el crecimiento folicular y la implantación embrionaria. La glucosa es la fuente de energía primaria para los ovarios. La disponibilidad de glucosa está directamente relacionada con la modulación de la secreción de LH, y la hipoglucemia severa inhibe la secreción pulsátil de LH, impidiendo la ovulación. Por otra parte, la prolactina puede inhibir el crecimiento folicular (12).
Efectos del estrés sobre la reproducción en bovinosLa temperatura ambiente elevada puede reducir la eficiencia reproductiva tanto en hembras como en machos, afectando la gametogénesis, la libido, el estro, la ovulación, la fertilidad, la implantación, la supervivencia embrionaria, la duración de la gestación y la habilidad materna de las madres, así como aumento de los problemas en el momento del parto (13, 14).
Las concentraciones de FSH están aumentadas durante el estrés calórico, probablemente debido a la reducción plasmática de la inhibina producida por los folículos comprometidos. Pero el aumento de FSH parece no ser suficiente para revertir los efectos de la caída en la concentración de LH y la reducción de la disponibilidad de precursores androgénicos para la síntesis de estradiol, ocasionando fallas en la expresión del estro y reducción de la fertilidad (13).
Así, Katayama (15) encontró interferencia en los niveles de cortisol sobre la onda pulsátil de la LH en ovejas y en novillas medicadas con ACTH, con reducción en la concentración de estradiol, retraso del pico de LH y de la ovulación y celos de poca intensidad.
En los machos el estrés altera la calidad, la motilidad y el desarrollo espermático, conllevando a fenómenos de subfertilidad en el hato (16). Ese efecto negativo es resultante de la degeneración testicular, una patología ocasionada entre otros muchos factores, por cualquier proceso que determine la elevación de la temperatura testicular, como por ejemplo el exceso de grasa escrotal, edema, periorquitis y elevación de la temperatura ambiente con consecuente estrés térmico (17).
Clínicamente, la degeneración testicular lleva inicialmente a la flacidez y discreta disminución del tamaño de los testículos, siendo que en etapas más avanzadas estos pueden tornarse atrofiados, con tamaño reducido, fibróticos y con consistencia firme a la palpación. Histológicamente, se nota en los túbulos seminíferos disminución de la espermatogénesis debido a la muerte de las células del linaje germinativo. Las espermatogonias presentan citoplasmas vacuolizados y núcleo picnótico, las espermátidas se degeneran y hay aparecimiento de células gigantes multinucleadas resultantes de la fusión de varias espermátidas (18).
Endocrinológicamente, el principal mecanismo por medio del cual el estrés puede dar lugar a una inadecuada calidad seminal se basa en la hormona conocida como CRH, la cual desencadena la cascada del estrés con acciones de tipo inhibitorio tanto a nivel testicular como a nivel central, con la inhibición de la hormona ICSH (Hormona estimuladora de las células intersticiales) (19).
La CRH es sintetizada a nivel hipotalámico, pero en situaciones estresantes también es secretada por las células intersticiales o de Leydig a nivel testicular (iniciado por serotoninas), actuando en los receptores de membrana como un potente regulador negativo para la ICSH, dando lugar a un bloqueo por medio de una proteína kinasa como respuesta al estrés. De esta manera, se impide la producción de andrógenos por dichas células, recordando el papel fundamental que la testosterona y la dihidrotestoterona ejercen a nivel de la espermatogénesis (19).
Las hormonas relacionadas con el estrés pueden influir en la función sexual en tres niveles del eje HHG: en el hipotálamo, a través de la CRH donde se inhibe la secreción de GnRH; en la hipófisis, disminuyendo la liberación de LH y FSH en respuesta a la inhibición de la GnRH y, en las gónadas, alterando el efecto estimulador de las gonadotrofinas (10).
Al disminuir la liberación de gonadotropinas (LH y FSH), la producción de estrógenos también se verá afectada, lo que conlleva a diversos trastornos reproductivos como fallas en la detección del celo provocadas por la presencia de estros silenciosos. Además de fallas en el desarrollo y calidad de los ovocitos, en la fertilización y la implantación del embrión (12) debido a la incorrecta preparación del útero y la formación de un cuerpo lúteo de mala calidad (20).
La interacción de la fisiología del estrés con la reproducción ha sido motivo de estudios recientes, que han demostrado que el estrés crónico o agudo puede afectar la dinámica folicular y la actividad del cuerpo lúteo, inhibiendo la liberación de LH (21).
La disminución en las tasas de concepción puede ser debido al desarrollo inadecuado del CL y a la producción insuficiente de P4 lútea (22, 23) o por un desarrollo embrionario deficiente. Bajos niveles de P4 circulante, en tiempos específicos de la gestación, ponen en riesgo su sostenimiento (24).
Los efectos del estrés sobre el comportamiento del estro incluyen una acción independiente del eje hipófisis-adrenal. Wilson et al. (25) reportan disminución en la concentración periférica de 17 B-estradiol a causa del Estrés Calórico (EC) que, según Pereria (10), es más una consecuencia de la reducción de la actividad física y metabólica, como respuesta adaptativa para limitar la producción de calor corporal.
El estrés por calor compromete el eje hipotálamo-hipófisis afectando la pulsatilidad de las gonadotropinas, lo cual a su vez incide de manera negativa sobre la expresión de signos que hagan evidente el celo al ocasionar alteraciones sobre el crecimiento folicular y conduciendo a la inhibición del desarrollo embrionario (26). En vacas de la raza Holstein expuestas al estrés por calor, con temperaturas superiores a los 29°C durante 20-50 días antes de la inseminación artificial (27), se observaron tasas de gestación menores que en vacas que no fueron expuestas a esa condición ambiental (23% vs 31,3%, p < 0,001).
En Israel se observó que, al refrescar las vacas Holstein un día antes del día esperado del estro, se logró un incremento en los signos de comportamiento estral (70% y 45%) (28).
Todavía no se conoce, con exactitud, el momento de mayor susceptibilidad al EC antes de la ovulación (29), sin embargo, Al-Katanani et al. (30) encontraron una asociación entre las altas temperaturas el día 10 antes de la ovulación, con una reducida fertilidad después de la inseminación.
En el ovarioEl EC actúa directamente sobre el ovocito y la función folicular, comprometiendo la calidad del mismo y promoviendo alteraciones de la dinámica folicular (31). En vacas bajo estrés entre los días 3 y 5 del ciclo estral se incrementa la concentración de androstenediona y se reduce la de estradiol en el líquido del folículo dominante, al reducirse la actividad de la enzima aromatasa (32).
Al promover estrés térmico (ET) en el día 11 del ciclo estral se aumenta el número de folículos mayores de 10 mm, y se ocasiona la emergencia precoz del folículo dominante de la segunda onda folicular (26).
Además, como consecuencia del estrés, se ha reportado una mayor ocurrencia de ciclos estrales de tres ondas de crecimiento folicular y reducción de la concentración de 17 B-estradiol circulante (25). El aumento de la longitud de los ciclos estrales genera una reducción de la concentración de E2, quien participa en el mecanismo uterino de luteólisis. Hansen y Aréchiga (26) observaron que, al interrumpir la exposición al EC, las vacas presentaban luteólisis y reinicio de su desarrollo folicular, y aunque los ovocitos existentes continuaran creciendo eran subfértiles al haber sido dañados por la exposición a temperaturas elevadas (33).
Hansen y Aréchiga (26) demostraron que el EC altera las concentraciones de P4 cirulantes, al afectar la tasa metabólica de las células luteales.
Las concentraciones de estradiol en el fluido folicular al octavo día del ciclo de vacas mantenidas en condiciones de confort, de acuerdo con Guzeloglu et al. (34), fueron más altas, al compararlas con las del grupo de animales bajo condiciones de ET (1662±189; 1493±188 ng/ml). Mientras que las concentraciones de progesterona mostraron el pico más alto en el grupo de vacas sometidas a estrés (54,1±5,1; 44,7±5,0 ng/ml), como se observa en la Tabla 1.

Durante la gestación
Se ha establecido que la susceptibilidad al EC en los estados tempranos de desarrollo embrionario, se debe a la ausencia de una proteína de choque térmico como HSP70 y disminución o ausencia en los niveles de glutatión (35), producidas por el propio embrión. Sin embargo, a medida que avanza su desarrollo el embrión logra producirlas, lo que le brinda termoprotección (36). Este grupo de proteínas actúa como chaperonas y cumple importantes funciones biológicas como el plegamiento y síntesis de otras proteínas, que favorecen a la vez su paso a través de las membranas en los diferentes compartimentos celulares como el retículo endoplásmico y la mitocondria, previniendo su desnaturalización (37).
Los efectos del EC sobre la preñez se relacionan con la disminución del peso del ternero al nacimiento, alteración de las concentraciones hormonales de la madre y del feto, y reducción de la producción de leche durante el posparto. La disminución en el peso del ternero del orden del 18% se atribuyó a una disminución en las concentraciones de sulfato de estrona (38).
Las alteraciones de comportamiento observadas generalmente por ET son: aumento en el consumo de agua (39), disminución de la rumia (40), anestro, disminución en las manifestaciones de los signos de estro, disminución en la cantidad y calidad del semen eyaculado (41, 42).
Los cambios fisiológicos causados por el estrés pueden ser diagnosticados mediante un examen clínico observando el grado de deshidratación, la frecuencia respiratoria, la frecuencia cardiaca, la temperatura rectal y de piel (43). También, pueden ayudar en el diagnóstico los análisis de laboratorio, bioquímicos, hematológicos y hormonales (31, 44).
El primer aspecto a considerar en la adaptación de los animales frente al estrés calórico es la selección de la raza, la cual debe ser adaptada a las difíciles condiciones de calor y humedad del clima tropical. Las razas cebuínas muestran mayor adaptación a las condiciones del trópico bajo. En el caso de traer animales de otras latitudes al trópico bajo, su exposición prolongada, desde temprana edad a las condiciones de estrés térmico, conduce a que los individuos sobrevivientes adquieran cierto grado de adaptación, sacrificando a su vez el rendimiento en los niveles de producción y en la capacidad reproductiva (45).
Una segunda opción, aceptando una justificación económica, consiste en una serie de medidas destinadas a mejorar las condiciones ambientales en que se encuentran los bovinos, como la implementación del modelo de ganadería silvopastoril, proporcionando sombra natural a los animales. Existe además, la alternativa del uso de sombras artificiales. Para la utilización de sombras artificiales, se pueden usar materiales como madera, aluminio, teja o malla polisombra en las fincas. Por otro lado, se puede recurrir al uso de sistemas de enfriamiento con agua para reducir la temperatura corporal, menos recomendables por su costo y operabilidad.
La tercera opción es el manejo nutricional del estrés calórico. Esta alternativa se basa en la respuesta natural del ganado de reducir de manera voluntaria el consumo de alimento para disminuir el calor metabólico. El uso de suplementos alimenticios energéticos y proteicos, son las medidas más utilizadas (45).
Selección de animales termotoleranesHansen et al. (46) reportan que al ser la tolerancia al calor una característica de alta heredabilidad (h² = 0,25 a 0,65), existen genes específicos que pueden ser seleccionados para aumentar la capacidad de termorregulación.
Así, Rocha et al. (47) no observaron cambios en las tasas de fertilización y el desarrollo embrionario en vacas Bos indicus, en comparación con vacas Bos taurus, donde se presentó mayor proporción de ovocitos anormales y menor porcentaje de oocitos fertilizados en verano en el estadío de 8 a 16 células. Además, Lopes et al. (48) encontraron mayores tasas de clivaje en Brahman (87%) que en Holstein y Angus (76 y 68%, respectivamente).
SombríoLos nuevos modelos de producción pecuaria, como el sistema silvopastoril, se basan en los principios de la sustentabilidad, que tiene como prioridad el confort térmico y el bienestar de los animales. La sombra surge como una alternativa para proteger los animales de la radiación solar, y se considera la modificación básica y más importante de las condiciones ambientales para disminuir el efecto del estrés calórico al reducir el esfuerzo del animal para el mantenimiento de su homeotermia y generar un menor gasto de energía. Está documentado que bajo la sombra de los árboles se logra reducir la temperatura ambiente entre 2 y 9°C en las horas más calurosas del día (48).
Investigadores como Pires (40) han demostrado que el bienestar animal es el factor determinante en la viabilidad técnica y económica de los sistemas de producción de leche.
Terapias hormonalesVarios estudios apuntan a la sincronización de la ovulación (IATF) como una de las técnicas que pueden solucionar los problemas por fallas en la detección de celo, aunque no reduce las tasas de mortalidad embrionaria (49).
El uso de protocolos para la inseminación artificial a tiempo fijo, ha permitido evitar la necesidad de detección de celo e inseminar las vacas a tiempo fijo. En vacas sometidas a estrés calórico se ha logrado incrementar la tasa de preñez después del parto (50).
Formulación de dietas específicas para animales bajo estrés térmicoEl uso de antioxidantes en la dieta ha sido estudiado para aumentar la fertilidad de los animales en verano, esta idea surgió del hecho de que el estrés térmico reduce la concentración intracelular de antioxidantes como el glutatión, en mórulas de ratones, y que la adición de varios antioxidantes en el medio de cultivo (taurina, glutation y vitamina E), promovieron algún grado de termoprotección en mórulas de ratones y bovinos. Sin embargo, Hansen y Aréchiga (26) no encontraron efecto positivo del suministro de la vitamina E por un período de 6 días antes del estro, sobre la fertilidad de los animales (estrés térmico).
Transferencia de Embriones (TE)Otra opción, para incrementar la proporción de hembras gestantes en la época calurosa, es optar por la transferencia de embriones de 6 a 7 días de edad. Los embriones deben ser colectados y almacenados antes de que la vaca donadora sea sometida al estrés calórico, como forma de aumentar el índice de éxito de la TE. Estos embriones evitan los efectos deletéreos del calor, evitando el momento crítico del desarrollo inicial (35). El uso de transferencia directa de embriones frescos presenta mejores resultados, con incrementos entre 10,8 y 15,7% en la tasa de concepción con respecto a la IA (26).
En términos generales el EC actúa en el animal de modo multifactorial, y su control depende de las medidas preventivas en varios niveles de actuación, desde la fase inicial del embrión hasta la final de la gestación. Todo productor, técnico y profesional que actúa en esta área de producción-reproducción debe estar atento a los efectos provocados por el estrés, particularmente severos en animales de alto potencial genético.
1. Cunningham JG, Klein BG. Fisiología Veterinaria. 5nd Edición. Elsevier Saunders; 2014. p. 700. [ Links ]
2. McEwen BS, Wingfield JC. The concept of allostasis in biology and biomedicine. Horm Behav 2003; 43:2-15. [ Links ]
3. Columbiano VS. Identificação de QLT nos cromossomos 10, 11 e 12 associados ao estresse calórico em bovinos. Dissertação Mestrado, Departamento de Zootecnia, Universidade Federal de Viçosa, Minas Gerais; 2007. [ Links ]
4. Ramos R. Avaliação de Estresse Térmico em Vacas Leiteiras Mestiças (Bos taurus x Bos indicus) Criadas em Clima Tropical Quente Ùmido no Estado do Ceará. Dissertação Graduação, Departamento de Zootecnia, Universidade Federal de Ceará; 2008. [ Links ]
5. Habib KE, Gold PW, Chrousos GP. Neuroendocrinology of stress. Endocrinol. Metab. Clin. North Am. 2001; 30:695-728. [ Links ]
6. Arias RA, Mader TL, Escobar PC. Factores climáticos que afectan el desempeño productivo del ganado bovino de carne y leche. Arch Med Vet 2008; 40:7-22. [ Links ]
7. Greenspan FS. Endocrinologia Básica e Clínica. 7nd Edición. Porto Alegre, Brasil: McGraw-Hill; 2006. p. 521. [ Links ]
8. Spinosa HS, Górniak SL, Bernardi MM. Farmacologia aplicada à Medicina Veterinária. 3nd Edición. Rio de Janeiro, Brasil: Guanabara Koogan; 2002. p. 752. [ Links ]
9. Kadzare CT, Murphi MR, Silanikove N, Maltz E. Heat stress in lactating dairy cows: a review. Livest Pro Sci 2002; 77(1):59-91. [ Links ]
10. Pereira CC. Fundamentos de Bioclimatologia Aplicados à Produção Animal. 1nd Edición. Belo Horizonte: FEPMVZ; 2005. [ Links ]
11. Chrousos GP. The role of stress and the hypothalamic-pituitary-adrenal axis in the pathogenesis of the metabolic syndrome: neuro-endocrine and target tissuerelated causes. Int. J. Obes. Relat. Metab. Disord. 2000; 249(2):50-55. [ Links ]
12. De Rensis F, Scaramuzzi JR. Heat Stress and seasonal effects on reproduction in the dairy cow. Theriogenology 2003; 60:1139-1151. [ Links ]
13. Furtado E, Araujo Filho. Influencia da temperatura ambiente na reprodução animal (Breve Revisão Literária). 2002 [consultado El 24 de mayo 2012]. Disponible en: http://www.ebah.com.br/content/ABAAABQMYAF/influencia-temperatura-ambiente-na-reproducao-animal [ Links ]
14. Perassoli E. Variação climática influencia nos índices de reprodução. ENIPEC; 2006. Disponible en: http://www.secom.mt.gov.br [ Links ]
15. Katayama KA. Efeito do manejo e de variáveis bioclimáticas sobre a taxa de gestação em vacas receptoras de embriões. Dissertação Mestredo, Facultade de Veterinaria, Universidade Federal de Mato Grosso do Sul. Mato Grosso do Sul; 2006. [ Links ]
16. Nardone A, Lacetera N, Bernabucci U, Ronchi B. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. Journal of dairy Science 1997; 80(5):838-44. [ Links ]
17. Nascimento E, Santos R. Patologia da reprodução dos animais domésticos. 2 edición. Rio de Janeiro: Guanabara Koogan; 2003. p. 93-104. [ Links ]
18. Santos R, Silva C, Ribeiro A, Vasconcelos A, Pesquero J, Coelho S, et al. Effect or growth hormone and induced IGF-1 release on germ cell population and apoptosis in the bovine testis. Theriogenology 1989; 51(5):975-984. [ Links ]
19. Lozano D, Vásquez P, González P. Effect of heat stress and its interaction with other management and productive variables on pregnancy rate in dairy cows in Aguascalientes, Mexico. Vet. Mex. 2005; 36(3): 245-260. [ Links ]
20. Grunert E, Birge HE, Vale GW. Patologia clínica da reprodução dos animais mamíferos domésticos: ginecologia. São Paulo: Livraria Varela; 2005. [ Links ]
21. Breen KM, Karsh FJ. Does cortisol inhibitor pulsatile luteinizing hormone secretion at the hypothalamic or pituitary level. Endocrinology 2003; 145(2):692-698. [ Links ]
22. Alnimer M, De Rosa G, Grasso F, Napolitano F, Bordi A. Effect of climate on the response to three oestrous synchronisation techniques in lactating dairy cows. Anim. Reprod. Sci. 2002; 71:157-168. [ Links ]
23. Cruz VJ, Elizondo VC, Ulloa AR, Fernández GI. The effect of GnRH after insemination on progesterone concentrations and conception rates in repeat-breeding Holstein cows under heat stress conditions. Tec. Pecu. Mex. 2009; 47(1):107-115. [ Links ]
24. Inskeep EK. Preovulatory, postovulatory, and postmaternal recognition effects of concentrations of progesterone on embryonic survival in the cow. J. Anim. Sci. 2004; 82(Suppl):24-39. [ Links ]
25. Wilson SJ, Kirby CJ, Koenigsfield AT, Keisler DH, Lucy MC. Effects of controlled heat stress on ovarian function of dairy cattle. J. Dairy Sci. 1998; 81:2132-2138. [ Links ]
26. Hansen PJ, Aréchiga CF. Strategies for managing reproduction in the heat stressed dairy cow. J. Anim. Sci. 1999, 77:36-50. [ Links ]
27. Chebel R, Santos J, Reynolds J, Cerri R, Juchem S, Overton M. Factors affecting conception rate after artificial insemination and pregnancy loss in lactating dairy cows. Anim. Reprod. Sci. 2004; 84:239-255. [ Links ]
28. Her E, Wolfenson D, Flamenbaum I, Folman Y, Kaim M, Berman A. Thermal, productive and reproductive responses of high yielding cows exposed to short-term cooling in summer. J. Dairy. Sci. 1988; 71:1085-1092. [ Links ]
29. Hansen P. Exploitation of genetic and physiological determinants of embryonic resistance to elevated temperature to improve embryonic survival in dairy cattle during heat stress. Theriogenology 2007; 68S:S242-S249. [ Links ]
30. Al-Katanani Y, Webb D, Hansen P. Factors affecting seasonal variation in 90 day non-retourn rate to first service in lactating Holstein cows in a hot climate. J. Dairy. Sci. 1999; 82:2611-2615. [ Links ]
31. Ferreira F, Pires M, Martinez M. Parâmetros clínicos, hematológicos, bioquímicos e hormonais de bovinos submetidos ao estresse calórico. Arq. Bras. Med. Vet. Zootec. 2009; 61(4):769-776. [ Links ]
32. Wolfenson D, Lew B, Thacher W, Graber Y, Meidan R. Seasonal and acute heat stress effects on steroid production by dominant follicles in cows. Anim. Reprod. Sci. 1997; 47:9-19. [ Links ]
33. Lucy MC. Estratégias de manejo de vacas leiteiras para melhoria dos índices reprodutivos durante o verão. Anais do V curso. Novos enfoques na produção e reprodução de bovinos; 2001. p. 12-18. [ Links ]
34. Guzeloglu A, Ambrose J, Kassa T, Diaz T, Thatcher M, Thatcher W. Long-term follicular dynamics and biochemical characteristics of dominant follicles in dairy cows subjected to acute heat stress. Anim. Reprod. Sci. 2001; 66:15-34. [ Links ]
35. Eduards, J, Hansen, P. Elevated temperature increases heat shock protein 70, synthesis in bovine two-cells embryons and compromises function in mature oocytes. Biol. Reprod. 1996; 55:340-346. [ Links ]
36. Bañuelos R, Silva JM, Aréchiga CF. La función de las proteínas de choque calórico en la reproducción animal. Orinoquia 2001; 5(1):41-50. [ Links ]
37. Ju JC. Cellular responses of oocytes and embryos under thermal stress: hints to molecular signaling. Anim. Reprod. 2005; 2:79-90. [ Links ]
38. Collier R, Doelger S, Head H, Thatch W, Wilcox C. Effect of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and pospartum Milk yield of Holstein cows. J. Anim. Sci. 1982; 54:309-319. [ Links ]
39. Meyer U, Stahl W, Flachowsky G. Investigations on the water intake of growing bulls. Livest. Prod. Sci. 2006; 103:186-191. [ Links ]
40. Pires M, Campos A. Conforto Animal para maior produção de leite. Viçosa-MG: CPT, 2008; p. 254. [ Links ]
41. Hafez ES. Reprodução Animal. 7nd Edición. São Paulo: Manole, 2003; p. 530. [ Links ]
42. Coelho l, Sasa A, Nader C. Características do ejaculado de caprinos sob estresse calórico em câmara bioclimática. Arq. Bras. Med. Vet. Zootec. 2006; 58:544-549. [ Links ]
43. Ferreira F, Pires M, Martinez M. Parâmetros fisiológicos de bovinos cruzados submetidos ao estresse calórico. Arq. Bras. Med. Vet. Zootec. 2006; 58(5):732-738. [ Links ]
44. Starling J, Silva R, Cerón-Muñoz M, Barbosa G, Costa M. Análise de Algumas Variáveis Fisiológicas para Avaliação do Grau de Adaptação de Ovinos Submetidos ao Estresse por Calor. Rev. Bras. Zootec. 2002; 31(5):2070-2077. [ Links ]
45. Góngora A, Hernández A. La reproducción de la vaca se afecta por las altas temperaturas ambientales. Rev. U.D.C.A. Actual. Divulg. Cient. 2010; 13(2):141-151. [ Links ]
46. Hansen J, Drost M, Rivera M, Lopes F, Al-Katanani M, Krininger E, et al. Adverse impact of heat stress on embryo production: causes and strategies for mitigation. Theriogenology 2001; 55:91-103. [ Links ]
47. Rocha A, Randel RD, Broussard JR, Lim JM, Blair RM, Roussel JD, et al. high environmental temperature and humity decrease oocyte quality in Bos Taurus but not in bos indicus. Theriogenology 1998; 49: 657-665. [ Links ]
48. Lopes F, Chase Jr, Al-Katanani Y, Krininger C, Rivera R, Tekin S. Breed differences in resistance of bovine preimplantation embryos to heat shock. Theriogenology 2001; 55:436. [ Links ]
49. Navas A. Importance of silvopastoral systems on caloric stress reduction in tropical livestock productions. Ver. Med. Vet. 2010; (19):113-122. [ Links ]
50. De la Sota R, Burkle J, Risco C, Moreira F, De Lorenzo M, Thatcher W. Evaluation of timed insemination during summer heat stress in lactating dairy cattle. Theriogenology 1998; 49:761-770. [ Links ]
