










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Ingenierías Universidad de Medellín
versão impressa ISSN 1692-3324
Rev. ing. univ. Medellín vol.11 no.21 Medellín jul./dez. 2012
ARTÍCULO ORIGINAL
FLUCTUACIÓN DE LOS ENSAMBLES PLANCTÓNICOS EN LA CIÉNAGA DE AYAPEL (CÓRDOBA-COLOMBIA) DURANTE UN CICLO SEMANAL*
FLUCTUATION OF PLANKTON ASSEMBLAGES AT AYAPEL SWAMP IN CÓRDOBA (COLOMBIA) DURING A WEEKLY CYCLE
Juan Carlos Jaramillo-Londoño**; Néstor Jaime Aguirre-Ramírez***
** Biólogo, M. Sc. Candidato a Doctor en Biología. Grupo GAIA. Universidad de Antioquia. Profesor Asociado. Facultad de Ingenierías. Universidad de Medellín. Carrera 87 N° 30-65. Apartado Aéreo 1983. Medellín - Colombia. Tel: (4) 340 5465. Fax: (4)340 5216. Correo electrónico jcjaramillo@udem.edu.co.
*** Doc. rer. nat. Profesor Facultad de Ingeniería. Grupo GAIA. Sede de Investigación Universitaria. Torre 2 Lab. 231. Universidad de Antioquia. Tel: 219 6562 Correo electrónico naguirre@udea.edu.co.
Recibido: 18/01/2012
Aceptado: 05/10/2012
RESUMEN
En esta investigación se evaluó la variación espacial y temporal de los ensamblajes fitoplanctónicos y zooplanctónicos, diariamente y durante un ciclo semanal, en seis estaciones de muestreo en la ciénaga de Ayapel (Córdoba, Colombia), tomando muestras integradas de la columna de agua. Ambos ensamblajes presentaron diferencias espaciales y una alta homogeneidad temporal. El fitoplancton estuvo dominado por cianobacterias principalmente Cylindropermopsis raciborskii y Planktolyngbya limnetica, y el zooplancton por rotíferos del género Brachionus.
PALABRAS CLAVE
fitoplancton, zooplancton, ciénagas, Colombia.
ABSTRACT
This research was intended to spatially and temporally evaluate phytoplankton and zooplankton assemblages on a daily basis during a weekly cycle, in six sampling stations at Ayapel swamp in Córdoba (Colombia), taking integrated samples of the water column. Both assemblages showed spatial differences and a high temporal homogeneity. Phytoplankton was dominated by cyanobacteria, specifically Cylindropermopsis raciborskii and Planktolyngbya limnetica; and zooplankton was dominated by Brachionus-genus rotifers..
KEY WORDS
Phytoplankton, Zooplankton, Swamps, Colombia
INTRODUCCIÓN
Las planicies de inundación se encuentran caracterizadas principalmente por anegamiento y/o inundación periódica, y son reconocibles como mosaicos de ecosistemas altamente dinámicos, de bordes lábiles, donde la estabilidad y la diversidad de las comunidades presentes están condicionadas principalmente por la hidrología y los flujos de materiales [1].
En Colombia, las planicies de inundación son denominadas regionalmente como ''ciénagas'', se encuentran definidas como ecosistemas poco profundos (usualmente no sobrepasan los 6 m) y están localizados a alturas inferiores a 1000 metros sobre el nivel del mar. Colombia posee aproximadamente 1 938 ciénagas que ocupan 478 419 hectáreas, cobertura que las sitúa como el principal sistema léntico del país [2]. Los servicios ambientales de estos ecosistemas se pueden resumir en tres grandes categorías: suministro de agua para diferentes usos, suministro de biomasa y suministro de beneficios no extractivos como la recreación y el transporte, entre otros [1, 3].
La determinación de los ciclos estacionales de la comunidad planctónica así como sus patrones de distribución espacial y sus interacciones tróficas con otras comunidades constituyen uno de los conjuntos de características básicas necesarias para la comprensión de los procesos ecológicos de cualquier ambiente de aguas lénticas [4]. Las ciénagas pueden sufrir cambios profundos en sus condiciones limnológicas y con ello afectar a las comunidades biológicas presentes; entender la dinámica del plancton en estos cuerpos de agua puede ser útil para evaluar la resiliencia de estos ecosistemas.
Una de las características relacionada con los patrones de desarrollo estacional de las comunidades planctónicas es la ocurrencia de fluctuaciones en cortos períodos de tiempo y su efecto sobre dichas comunidades. Este tema ha sido abordado principalmente en embalses de zonas templadas y subtropicales [5, 6] y en embalses tropicales [4, 7, 8]. Este estudio pretende evaluar la variación espacial y temporal de los ensambles fitoplanctónicos y zooplanctónicos de la Ciénaga de Ayapel durante un ciclo semanal.
1 MATERIALES Y MÉTODOS
La ciénaga de Ayapel (figura 1) está ubicada al norte de Colombia (8° 19' Norte y 75° 6' Oeste); en época de aguas altas tiene una extensión aproximada de 106 Km2, se ubica a 22 metros de altura sobre el nivel del mar, en una zona de bosque húmedo tropical, en donde se presenta una época de lluvias de abril a noviembre y una época seca de diciembre a marzo. El promedio anual de lluvias fluctúa entre 2 000 y 2 500 mm. La temperatura ambiente alcanza valores mayores a 25°C.
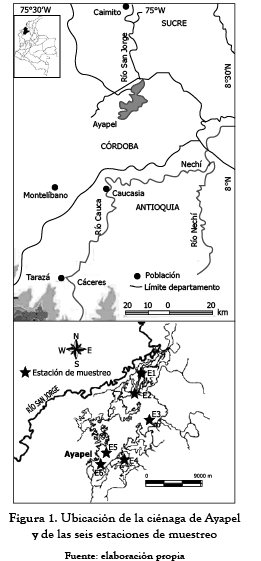
Para la toma de las muestras del plancton se construyó un tubo de PVC de 2 m de longitud y 10 cm de diámetro (figura 2). Las muestras de plancton y de variables físicas y químicas fueron tomadas diariamente en seis estaciones de muestreo, entre el 8 abr 2008 y el 14 abr 2008, correspondiente al período de menor nivel de agua de la ciénaga.
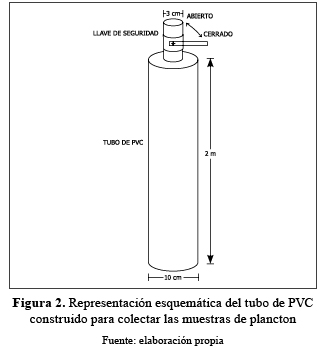
El fitoplancton se tomó introduciendo el tubo hasta una profundidad máxima de 2 m; el material colectado se filtró en una red cónica de 20 m de abertura de malla y 24 cm de diámetro con un frasco vial de 50 mL atado en el extremo inferior de la misma, concentrándose el material filtrado; el volumen total filtrado fue de 20 L y una vez colectado, el frasco se retiró y fijó con una solución de Lugol al 10%, para la posterior identificación y conteo de los organismos en el laboratorio.
Para la observación de las muestras se utilizó un microscopio invertido Leica DMIN provisto de reglilla ocular; la muestra de agua proveniente del campo fue agitada y de ella se extrajo una alícuota de 1 mL que se depositó en una cámara de Sedgwick-Rafter para su conteo.
Por medio de una curva de acumulación de especies se definieron 30 campos de observación siguiendo una trayectoria sinusoidal. Las observaciones se realizaron en un aumento de 400X, y para calcular la densidad de organismos por mililitro se aplicó la expresión de Ros [9]. La determinación de los taxones fitoplanctónicos se realizó hasta el mayor nivel taxonómico posible.
El zooplancton se tomó introduciendo el tubo hasta una profundidad máxima de 2 m; el material colectado se filtró en una red cónica de 64 m de abertura de malla y 24 cm de diámetro con un frasco vial de 50 mL atado en el extremo inferior de la misma concentrándose el material filtrado; el volumen filtrado fue de 100 L y una vez colectado el frasco se retiró y fijó con una solución de formalina al 4%, para la posterior identificación y conteo de los organismos en el laboratorio.
La observación de las muestras se realizó en un microscopio Leica DMLB; la muestra de agua proveniente del campo fue agitada y de ella se extrajo una alícuota de 1 mL que se depositó en una cámara de Sedgwick-Rafter para su conteo. La cámara fue contada en su totalidad y hasta completar un mínimo de individuos de 400, para garantizar una precisión del 90% [4] en un aumento de 100X y se calculó la densidad de organismos por litro; la determinación de los taxones zooplanctónicos se realizó, en su mayoría, hasta especie.
Las variables medidas diariamente, in situ, en cada estación y los métodos empleados para las mediciones se presentan en la tabla 1.
A cada una de las variables físicas y químicas se les calculó la media como medida de tendencia central, la desviación estándar como medida de dispersión absoluta y el coeficiente de variación de Pearson como medida de dispersión relativa.
Con el fin de establecer diferencias de las variables físicas y químicas entre las estaciones y días de colecta se aplicó un análisis bilateral de la varianza de Friedman y para establecer las estructuras de los ensamblajes fitoplanctónicos y zooplanctónicos se calcularon la riqueza numérica de especies, la diversidad d Shannon, la equidad de Pielou y la dominancia de Simpson.
2 RESULTADOS
Los valores medios de las variables físicas y químicas en cada una de las estaciones de muestreo y el análisis exploratorio de las mismas se muestran en la tabla 2.
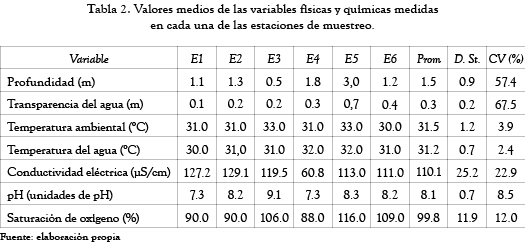
La transparencia del agua y la profundidad fueron las variables que más variación presentaron; los coeficientes de variación obtenidos fueron de 67.5% y 57.4%, respectivamente; las demás variables presentaron fluctuaciones bajas y, en general, fueron menores al 22.9%.
La prueba de Friedman (tabla 3 y figura 3) mostró diferencias estadísticamente significativas entre estaciones para todas las variables físicas y químicas evaluadas, con excepción de la temperatura ambiental.
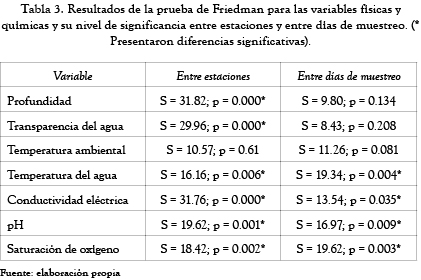
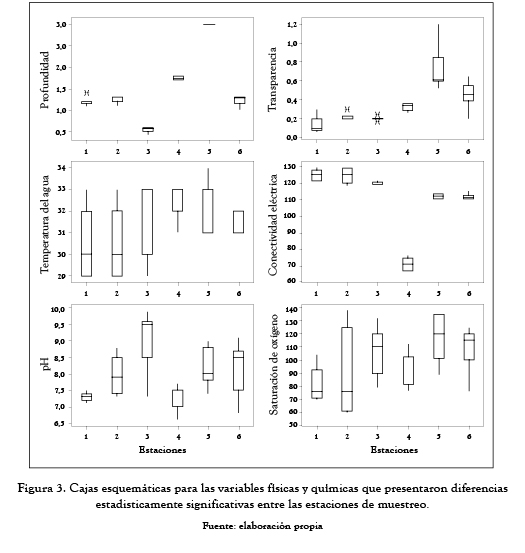
Las estaciones 1, 2, 4 y 6 presentaron una profundidad similar entre sí; la mayor diferencia se encontró entre la estación 3 (menos profunda) y la estación 5 (más profunda); la transparencia del agua, por su parte, fue baja en todas las estaciones de muestreo y se incrementó de la estación 1 a la estación 6; la conductividad eléctrica presentó los valores más bajos en la estación 4; las demás estaciones presentaron valores mayores y con una tendencia a disminuir de la estación 1 a la estación 6; el pH mostró un ligera tendencia alcalina de la estación 1 a la estación 3 y de la estación 4 a la estación 6, y finalmente, la saturación del oxígeno se incrementó de la estación 1 a la 6.
El ensamblaje fitoplanctónico en la ciénaga de Ayapel (tabla 4 y figura 4) estuvo compuesto por 74 morfoespecies pertenecientes a siete divisiones: Chlorophyceae (31 morfoespecies), Cyanobacteria (17 morfoespecies), Bacillariophyceae (12 morfoespecies), Euglenophyta y Zygnemophyceae (5 morfoespecies cada una), Xantophyceae (2 morfoespecies) y Rhodophyta y Dynophyta (1 morfoespecie cada una).

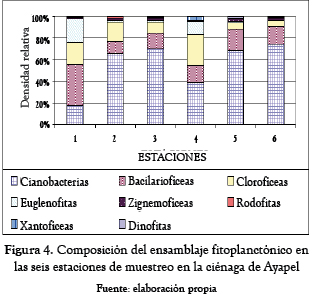
La mayor densidad de organismos correspondió a Cyanobacteria con el 68.8% del total, le siguen Bacillariophyceae (14.6%) y Chlorophyceae (12.3%); los demás grupos presentaron densidades muy bajas, Euglenophyta (2.1%), Zygnemophyceae (1.3%) y Rhodophyta, Dinophyta y Xanthophyceae no superaron cada una el 0.5% de la densidad total.
Las especies fitoplanctónicas con las mayores densidades fueron Cylindrospermopsis raciborskii (635.0 ind/mL), Planktolyngbya limnetica (513.2 ind/mL), Aphanizomenon sp (374.3 ind/mL), Aulacoseira granulata (277.3 ind/mL), Oscillatoria sp (228.3 ind/mL), Synedra sp (141.0 ind/mL) y Anabaena circinalis (109.0 ind/mL).
La estación 1 mostró un predominio de las bacilariofíceas, y otros grupos como las cianobacterias, clorofíceas y euglenofitas también presentaron contribuciones considerables en su densidad relativa; la estación 4 mostró un patrón similar, aunque la contribución de las cianobacterias y clorofíceas fue mayor que los otros dos grupos y se apreció una ligera contribución de las xantofíceas. Para las estaciones 2, 3, 5 y 6 las cianobacterias fueron el grupo predominante y, en menor proporción, las bacilariofíceas y cloroficeas.
El ensamblaje zooplanctónico en la ciénaga de Ayapel (tabla 5 y figura 5) estuvo compuesto por 35 especies de los grupos Rotifera (25 especies), Cladocera (7 especies) y Copepoda (3 especies).
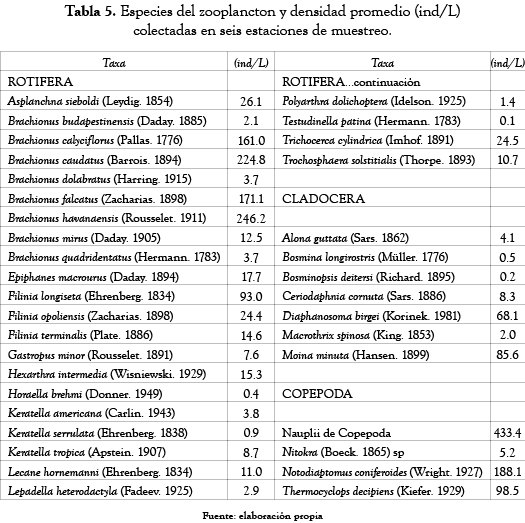
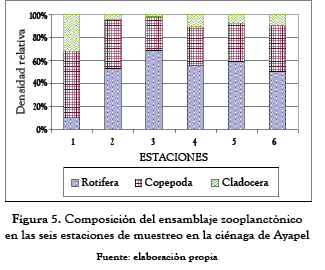
La mayor densidad fue para los rotíferos (54.9%), le siguieron los copépodos (36.6%) y finalmente los cladóceros con el 8.5% de la densidad total.
En el grupo de los rotíferos las mayores densidades fueron para las especies Brachionus havanaensis (246.2 ind/L), B. caudatus (224.8 ind/L), B. falcatus (171.1 ind/L), B. calyciflorus (161 ind/L); en el grupo de los copépodos, las formas inmaduras (433.4 nauplios/L) fueron los que presentaron la mayor densidad de todos los organismos colectados y los adultos de la especie Notodiaptomus coniferoides alcanzaron densidades de 188.1 ind/L. Por último, las especies Moina minuta (85.6 ind/L) y Diaphanosoma birgei (68.1 ind/L) fueron los cladóceros que presentaron las mayores densidades durante el período de estudio.
Los copépodos mantuvieron unas densidades similares a lo largo de las seis estaciones de muestreo, mientras que los rotíferos y cladóceros mostraron un comportamiento inverso; en aquellos sitios donde la densidad de rotíferos fue mayor los cladóceros presentaron las menores densidades y viceversa.
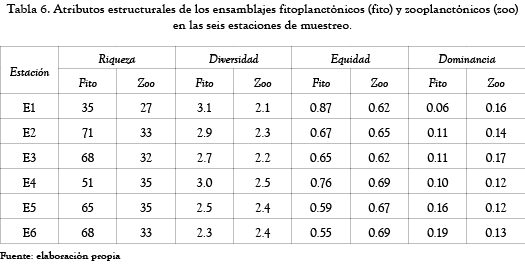
La tabla 6 muestra los valores de los atributos estructurales calculados para cada uno de los ensamblajes planctónicos en la ciénaga. Los valores de la riqueza de especies, la diversidad y la equidad del fitoplancton fueron mayores que los obtenidos para el zooplancton y en el caso de la riqueza de especies del fitoplancton prácticamente duplicó en número a la riqueza de especies de zooplancton encontradas.
3 DISCUSIÓN Y CONCLUSIONES
Durante el período de estudio la ciénaga de Ayapel mostró una alta homogeneidad temporal y diferencias notables en el nivel espacial con respecto a las variables físicas y químicas medidas. Las estaciones 4 y 5 fueron las de mayor profundidad; las demás estaciones presentaron profundidades bajas. La turbiedad del agua es un característica importante en la ciénaga, lo que ocasiona una transparencia reducida en todas las estaciones de muestreo, principalmente en aquellas estaciones más someras; esa disminución de la transparencia puede ser causada por la resuspensión de los sedimentos del fondo como efecto de la acción del viento sobre el espejo de agua que genera una zona de mezcla que puede llegar a alcanzar, en algunos sitios, a toda la columna de agua; adicionalmente se observó un gradiente de la estación 1 (menor transparencia) a la estación 6 (mayor transparencia), una clara evidencia del efecto ejercido por el río San Jorge sobre la ciénaga el cual aporta gran cantidad de material en suspensión que influye directamente en la transparencia del agua.
Con referencia a la temperatura, se considera que los ecosistemas tropicales presentan como característica fundamental una temperatura aproximadamente uniforme a lo largo de todo el año; esta uniformidad se vio reflejada en los coeficientes de variación tan bajos que se obtuvieron.
Teniendo en cuenta que para la ciénaga se encontraron valores de conductividad inferiores a 130 µS/cm, este sistema puede considerarse como un ambiente con baja concentración de iones.
Los niveles de pH encontrados en la ciénaga muestran un sistema alcalino, que puede ser el producto de la remoción del CO2 por parte del fitoplancton, que durante esta época incrementa su densidad en la ciénaga. Y la saturación de oxígeno alcanzó valores mayores a 88%, incluso valores superiores al 100% en algunas de las estaciones, lo que muestra una cierta condición autotrófica del sistema para este momento hidrológico.
El ensamble fitoplanctónico en el sistema de Ayapel estuvo compuesto por 74 morfoespecies, cifra bastante mayor, si se compara con la riqueza reportada para este ecosistema por Hernández-Atilano et al. [10], quienes encontraron solo 37 taxones; sin embargo, se mantiene el predominio del grupo de las cianobacterias y las bacilariofíceas, que aportaron las mayores densidades, y el grupo de las clorofíceas, el mayor número de especies.
Las especies de los géneros Cylindrospermopsis y Planktolyngbya fueron las más abundantes, estas especies se caracterizan por presentar bajas tasas de crecimiento, muy baja herbivoría por parte del zooplancton y, adicionalmente, poseen una alta capacidad de suspensión, gracias a lo cual, las pérdidas de individuos por sedimentación son muy bajas. Todo lo anterior hace que estas poblaciones sean muy estables una vez que se establecen, compensando así sus bajas tasas de crecimiento; adicionalmente, Cylindrospermopsis es un típico representante de aguas cálidas de capas mezcladas y supremamente tolerante a deficiencias de nitrógeno y luz, situación común en la ciénaga, donde se presenta una zona fótica bastante reducida (inferior a un metro en la mayoría de las estaciones). Las cianobacterias compiten con otras especies fitoplanctónicas por luz y nutrientes, y proliferan en condiciones turbias o cuando los niveles de luz son bajos [11] y, dependiendo del sistema, esta capacidad puede hacer que se conviertan en un grupo altamente dominante y resiliente. En términos generales, se ha reportado que muchas cianobacterias parecen ser menos comestibles para el zooplancton y los peces, comparados con otros grupos algales. Pese a que C. raciborskii alcanzó densidades de 635 ind/mL en la ciénaga esta especie no se constituye como una buena oferta alimenticia para el sistema y el peso recaería sobre otras poblaciones como las bailariofíceas, que fueron abundantes durante el período de muestreo.
El ensamble zooplanctónico en la ciénaga de Ayapel estuvo conformado por los grupos Rotifera, Copepoda y Cladocera. Su composición presentó homogeneidad temporal y variaciones espaciales notorias; las densidades más altas fueron para los rotíferos; igualmente, este grupo presentó la mayor cantidad de especies. La dominancia de rotíferos ha sido asociada al incremento en las condiciones tróficas debido a su capacidad para ingerir pequeñas partículas, tales como bacterias y detritos orgánicos, para [12], y puede estar relacionada con sus características oportunistas (especies r-estrategas, adaptadas a rápido crecimiento poblacional durante estaciones favorables cortas), situación que se presenta en ambientes inestables y dinámicos como la ciénaga de Ayapel.
Otro factor que contribuye más al éxito de los rotíferos limnéticos es su plasticidad para adaptarse a diferentes fuentes alimenticias; esta característica, sumada a la baja presión de predación, por su pequeño tamaño, les proporciona ventajas competitivas sobre los otros grupos zooplanctónicos y este grupo juega un papel muy importante en la ciénaga, principalmente debido a su habilidad para filtrar bacterias y fitoplancton de pequeño tamaño, que son comunes en este ecosistema.
También se destaca una alta densidad de formas larvales de los copépodos; esas formas inmaduras son de gran importancia en la estructura de los ensambles zooplanctónicos en cuanto a su dinámica y aspectos tróficos. Mientras los nauplios y primeros estadios de desarrollo de los copepoditos del orden Cyclopoida son filtradores y predominantemente herbívoros, los últimos estadios de los copepoditos y los adultos tienen hábito raptorial y son predominantemente carnívoros [13], y de acuerdo con Edmondson [14], la alta proporción de formas inmaduras de copépodos es el resultado de la reproducción continua de estos organismos en regiones tropicales, y la alta densidad de nauplios en relación con los copépodos adultos puede ser un indicador de una alta tasa de mortalidad durante las diferentes etapas de desarrollo de estos organismos [15]. Un factor que podría estar determinando la proporción de formas jóvenes y adultos es la intensidad de la predación y el balance entre la predación por invertebrados y vertebrados, puesto que los copépodos representan gran parte de los elementos alimenticios de peces jóvenes y adultos [16]. Para la ciénaga de Ayapel no se tiene información sobre el impacto de los peces sobre la comunidad zooplanctónica; sin embargo, la alta abundancia y riqueza de especies de peces observada en la ciénaga es un importante factor que puede ser considerado en futuros estudios.
Los cladóceros presentaron en términos generales densidades muy bajas, solo Moina minuta y Diaphanosoma birgei fueron ligeramente dominantes; las bajas densidades de cladóceros pueden ser consecuencia de la gran cantidad de sólidos presentes en el agua; en general, se considera que las partículas suspendidas pueden afectarles ya sea por interferencia mecánica o indirectamente debido a la disminución de la luz y la producción del fitoplancton; sin embargo, el comportamiento alimenticio de estos organismos y otras interacciones biológicas en aguas turbias aún no está bien comprendido.
M. minuta es una especie tolerante de la turbidez, por ello es abundante en este ecosistema, y de acuerdo con [17], este cladócero tiene un alto valor nutricional y por medio de su reproducción partenogénica alcanza altas densidades poblacionales en cortos períodos de tiempo.
Se ha considerado que los cladóceros son mejores competidores que los rotíferos porque son de mayor tamaño, son más eficientes filtrando su alimento, e interfieren mecánicamente con los rotíferos que son atrapados en su cámara branquial [18]. Sin embargo, en la ciénaga de Ayapel los rotíferos son abundantes y los cladóceros son escasos; de acuerdo con [19] esta escasez puede ser una consecuencia del descenso en la rata de ingestión de fitoplancton debido a la alta concentración de partículas inorgánicas suspendidas en la columna de agua, y en general, cuando la densidad de rotíferos se incrementaba, la densidad de los cladóceros disminuía, y viceversa.
Finalmente, considerando que el pulso hidrológico se configura como el principal factor determinante de la dinámica de ecosistemas de planicie de inundación [19, 20] se espera que los ensambles fitoplanctónicos y zooplanctónicos sean profundamente influenciados por los pulsos hidrológicos de los ríos. En el caso de la ciénaga de Ayapel los cambios en estos ensambles no fueron fácilmente evidenciados debido a que la ciénaga se encontraba en un momento en el que el nivel del agua permaneció constante para el período de muestreo; se esperaría que esos cambios fueran mucho más evidentes si el muestreo se realizara en momentos en que el nivel se estuviera incrementando o decreciendo en función del pulso antes mencionado.
REFERENCIAS
[1] W. J. Mitsch y J. G. Gosselink, Wetlands. Third Edition, New York, John Wiley & Sons, Inc., 2000, 920 p. [ Links ]
[2] P. Arias, ''Las ciénagas en Colombia,'' Divulgación Pesquera, INDERENA (Bogotá), vol. XXII, n.° 2, 3, 4, pp. 39-70, 1985. [ Links ]
[3] A. Ramírez y G. Viña, Limnología Colombiana: Aportes a su conocimiento y estadísticas de análisis. Fundación Universidad de Bogotá Jorge Tadeo Lozano, 1998, 294 p. [ Links ]
[4] R. M. Pinto-Coelho, ''Fluctuações sazonais e de curta duração na comunidade zooplanctônica do lago Paranoá, Brasília-DF, Brasil,'' Revista Brasileira de Biologia, vol. 47, no. 1/2, pp. 17-29, 1987. [ Links ]
[5] R. J. Bernot et al., ''Spatial and temporal variability of zooplankton in a great plains reservoir,'' Hydrobiologia, vol. 525, pp. 101-112, 2004. [ Links ]
[6] G. Parra et al., ''Short term fluctuations of zooplankton abundance during autumn ciculation in two reservoirs with contrasting trophic state,'' Limetica, vol. 28, no. 1, pp. 175-184, 2009. [ Links ]
[7] R. Henry et al., ''Annual and short-term variability in primary productivity by phytoplankton and correlated abiotic factors in the Jurumirin reservoir (São Paulo, Brazil),'' Brazilian Journal of Biology, vol. 66, n.°. 1B, pp. 239-261, 2006. [ Links ]
[8] P. Acuña et al., ''Short-term responses of phytoplankton to nutrient enrichment and planktivorous fish predation in a temperate South American mesotrophic reservoir,'' Hydrobiologia, vol. 600, pp. 131-138, 2008. [ Links ]
[9] J. D. Ros, Prácticas de Ecología, Barcelona, Ediciones Omega, SA, 1979, 235 p. [ Links ]
[10] E. Hernández-Atilano et al., ''Variación espacio temporal de la asociación fitoplanctónica en diferentes momentos del pulso hidrológico en la cienaga de Ayapel (Córdoba), Colombia,'' Actualidades Biológicas, vol. 30, n.°o. 88, pp. 67-81, 2008. [ Links ]
[11] J. Padisák, ''Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, hightly adaptive cyanobacterium: worldwide distribution and review of its ecology,'' Archiv fur Hydrobiologie Supplement, vol. 4, pp. 563-593, 1997. [ Links ]
[12] T. Matsumura-Tundisi y J. G. Tundisi, ''Plankton richness in an eutrophic reservoir (Barra Bonita Reservoir, SP, Brazil),'' Hydrobiologia, vol. 542, pp. 367-378, 2005. [ Links ]
[13] J. F. Neves et al., ''Zooplankton community structure of two marginal lakes of the river Cuiabá (Mato Grosso, Brazil) with analysis of Rotifera and Cladocera diversity,'' Brazilian Journal of Biology, vol. 63, n.° 2, pp. 329-343, 2003. [ Links ]
[14] W. T. Edmondson, Freshwater Biology, 2a ed., John Wiley, 1959, 1248 p. [ Links ]
[15] L. P. Sartori et al., ''Zooplankton fluctuations in Jurumirim Reservoir (São Paulo, Brazil): a three-year study,'''' Brazilian Journal of Biology, vol. 69, n.° 1, pp. 1-18, 2009. [ Links ]
[16] F. A. Lansac-Tôha et al., ''Composição da dieta alimentar de Hypophthalmus edentatus Spix, 1982 (Pisces, Hypophthalmidae) no reservatório de Itaipu e no rio Ocoí. 1978,'' Revista Unimar, Maringá, vol. 13, no. 2, pp. 147-162, 1991. [ Links ]
[17] L. H. Sipaúba-Tavares y O. Rocha, ''Produção de plâncton (fitoplâncton e zooplâncton) para alimentação de organismos aquáticos,'' São Carlos, Rima, 2003, 106 p. [ Links ]
[18] J. C. Jaramillo-Londoño y R. M. Pinto-Coelho, ''Interaction between Hexarthra intermedia (Rotifera) and Bosmina longirostris (Cladocera): a case of opportunistic nutrition or interference competition?,'' Journal of Plankton Research, vol. 32, n.° 6, pp. 961-966, 2010. [ Links ]
[19] K. L. KIRK, ''Inorganic particles alter composition in grazing plankton: the role of selective feeding, '' Ecology, vol. 72, pp. 915-923, 1991. [ Links ]
[20] W. J. Junk et al., ''The ''flood pulse'' concept in river floodplain systems,'' Canadian Journal of Fisheries and Aquatic Science (Spec. Publ.), vol. 106, pp. 110-127, 1989. [ Links ]
[21] J. J. Neiff, ''El régimen de pulsos en ríos y grandes humedales de Sudamérica, '' en Tópicos sobre humedales subtropicales y templados de Sudamérica, A. I. Malvárez, ed., 229 p, Oficina Regional de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe. Montevideo, Uruguay, 1999. [ Links ]
Notas:
*Artículo resultado de la investigación asociada a la tesis de doctorado en Biología del autor principal.