










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiotecnología en el Sector Agropecuario y Agroindustrial
Print version ISSN 1692-3561
Rev.Bio.Agro vol.10 no.2 Popayán July/Dec. 2012
CORRELACIÓN DE LA CONCENTRACIÓN DE POLIAMINAS CON EL DESARROLLO 'IN VITRO' DE Annona muricata L.
CORRELATION OF THE CONCENTRATION POLYAMINES WITH THE 'IN VITRO' DEVELOPMENT OF Annona muricata L.
CORRELAÇÃO ENTRE A CONCENTRAÇÃO DE POLIAMINAS COM O DESENVOLVIMENTO 'IN VITRO' DE Annona muricata L.
ALEYDA M. ACOSTA R.1, ENRIQUE J. PEÑA S.2, MARTHA I. PAEZ3, RICARDO BENITEZ B.4
1 Magister, Departamento de Biología. Facultad de Ciencias Naturales y Exactas. Universidad del Valle.
2 Doctor, Departamento de Biología. Facultad de Ciencias Naturales y Exactas. Universidad del Valle.
3 Doctor, Departamento de Química. Facultad de Ciencias Naturales y Exactas. Universidad del Valle.
4 Doctor, Profesor titular Departamento de Química. Facultad de Ciencias Naturales, Exactas y de la Educación. Universidad del Cauca.
Correspondencia: leydaacosta@gmail.com
Recibido para evaluación: 08/12/2010 Aprobado para publicación: 18/05/2012
RESUMEN
Annona muricata es promisoria para el cultivo por su importancia alimenticia y la exquisitez de su fruto. El carácter recalcitrante de las yemas vegetativas de la especie ha dificultado la micropropagación de material vegetal adulto previamente seleccionado por su alta productividad. Este trabajo tuvo como objetivo identificar la relación entre el desarrollo vegetal 'In Vitro' de Annona muricata y los niveles endógenos de poliaminas. El desarrollo vegetal 'in Vitro' fue promovido por microinjertación en serie y los niveles endógenos de poliaminas libres fueron cuantificados por Cromatografía Líquida de Alta Eficiencia (CLAE). Los resultados indican que la espermina es la poliamina de mayor abundancia en los brotes (54,5%), seguida de putrescina (27,6%) y espermidina (17,9%). La espermina decrece con el aumento de las series de microinjertación y la edad del brote, esta situación es similar para el contenido de poliaminas totales. En esta investigación se encontraron varias correlaciones entre el desarrollo vegetal 'In Vitro' de Annona muricata y las poliaminas confirmando su función como marcadores fisiológicos.
PALABRAS CLAVES: Microinjertación, Putrescina, Espermidina, Espermina.
ABSTRACT
Annona muricata is promising for crop because of its importance like food and exquisite fruit. The recalcitrant nature of the vegetative buds of the specie has dificulted the micropropagation of plant material mature that previously it was selected for its high productivity. This study aimed to identify the relationship between plant development 'In Vitro' of Annona muricata and endogenous levels of polyamines. Plant development 'In Vitro' was promoted by serial micrografting and the endogenous levels of polyamines were quantified by High-Performance Liquid Chromatography (HPLC). The results indicate that spermine is the most abundant in shoots (54,5%), followed by putrescine (27,6%) and spermidine (17,9%). The spermine decreases with increasing series of micrografting and age of the shoot, this situation is similar for the total content of polyamines. This research found several correlations between plant development 'in Vitro' of Annona muricata and polyamines, reaffirming their role as physiological markers.
KEY WORDS: Micrografting, Putrescine, Spermidine, Spermine.
RESUMO
Annona muricata é promissor para o cultivo de alimentos para a sua importância e delicadeza de seus frutos. A natureza recalcitrante da gemas vegetativas das espécies tem dificultado a micropropagação de plantas cultivadas previamente selecionados para alta produtividade. Este estudo teve como objetivo identificar a relação entre o desenvolvimento das plantas 'In Vitro' de Annona muricata e nos teores endógenos de poliaminas. Desenvolvimento das plantas 'In Vitro' foi promovido por microenxertia em série e os níveis endógenos de poliaminas livres foram medidos por cromatografia líquida de alta eficiencia (CLAE). Os resultados indicam que as poliaminas espermina é o mais abundante na parte aérea (54,5%), seguida da putrecina (27,6 %) e espermidina (17,9%). Espermina diminui com o aumento da microenxertia série e idade da epidemia, a situação é semelhante ao teor total de poliaminas. Nesta pesquisa, encontramos uma série de correlações entre o desenvolvimento das plantas 'in Vitro' de mudas de gravioleira e poliaminas confirmando seu papel como marcadores fisiológicos.
PALAVRAS-CHAVE: Microenxertia, Putrescina, Espermina, Espermidina.
INTRODUCCIÓN
Las poliaminas (PAs) son compuestos nitrogenados que se encuentran en todos los organismos y son esenciales para la viabilidad de las células. Son moléculas alifáticas de bajo peso molecular que en los tejidos se encuentran en forma libre o conjugada con otras biomoléculas [1-3].
Las PAs se consideran reguladoras de los procesos celulares y fisiológicos en las plantas, ya que, están implicadas en la división celular, en la síntesis de proteínas, en la organogénesis, la embriogénesis, en el desarrollo de flores, frutos, polen, nudos, raíces, en la germinación, fotosíntesis y respuestas al estrés [4, 10].
La Putrescina (Put), la Espermidina (Spd) y la Espermina (Spm) son las principales PAs encontradas en las plantas y en general, los tejidos vegetales juveniles contienen más PAs que los tejidos adultos [11]. En las plantas superiores los altos niveles de PAs son característicos de tejidos meristemáticos y en crecimiento activo; cada órgano de la planta presenta un patrón específico y está correlacionado con los procesos de división o expansión celular [12-14].
Las PAs como indicadores de la condición fisiológica de las plantas sirven para evaluar su desarrollo cuando existen evidencias de problemas de propagación 'in Vitro' como es el caso de Annona Muricata L. Esta especie se caracteriza porque produce la fruta más grande dentro de la familia de las Annonáceas y su importancia alimenticia como fuente de calcio, fósforo, fibra y vitaminas, ha generado un interés por tecnificar comercialmente su cultivo [15]. No obstante, los estudios de propagación 'in Vitro' de A. muricata evidencian problemas de oxidación, latencia, fenolización de explantes y problemas con la formación de raíces [16-19].
Dado que las PAs están involucradas en importantes procesos biológicos, esta investigación tuvo como objetivo identificar la relación entre los niveles endógenos de poliaminas libres (Put, Spd y Spm) y el desarrollo vegetal 'in Vitro' de Annona muricata L. En la revisión bibliográfica realizada no se encontraron estudios que relacionen a las PAs con el desarrollo vegetal de esta especie.
MÉTODO
Material vegetal: en el año 2009 se realizó un experimento de revigorización de material adulto de Annona muricata L. por medio de la microinjertación en serie con el objetivo de recuperar las propiedades organogénicas del tejido y facilitar su propagación 'In Vitro' [20]. En el diseño experimental se probaron tres tratamientos de microinjertación en serie y un control así: M1 fueron estacas microinjertadas en un portainjerto, M2 fueron estacas microinjertadas en dos veces, es decir, permanecieron en dos portainjertos diferentes con un lapso de un mes entre cada traspaso, M3 fueron estacas microinjertadas tres veces, es decir, permanecieron en tres portainjertos diferentes con un lapso de un mes entre cada traspaso y el Control fueron estacas sin microinjertar extraídas directamente de la planta madre adulta. Al finalizar los tratamientos se tomaron 300 mg de material vegetal seleccionado al azar (brotes en M1, M2 y M3, y yemas en el control, dado que en este ultimo tratamiento no hubo desarrollo de brotes). Este material fue liofilizado (en un Liofilizador Telstar- Italia, Ref. LyoQuest-55) y almacenado en un congelador (Challenger – Colombia, Ref. IBDC-216) a -20°C para su posterior análisis de PAs, por CLAE (Cromatografía líquida de alta resolución). Se realizaron tres réplicas por tratamiento.
Cuantificación de poliaminas: a) Extracción. El contenido de PAs libres en Annona muricata L. fue analizado modificando el método de Aldesuquy [21], para ello, el material vegetal liofilizado (300 mg) fue macerado y homogenizado en 2mL de TCA (50 g/L-1 de Ácido tricloroacético, Sigma Aldrich-USA, Referencia T6399-250G), la mezcla se centrifugó (centrífuga Hettich – Alemania. Ref. Universal 32R) a 11500 rpm por 15 min. a 4°C y se recuperó el sobrenadante en tubos de vidrio que permanecieron en un recipiente con hielo por 30 minutos. El extracto se mezcló con 5mL de dietil-eter (Sigma Aldrich - USA, Ref. 309966-1L), se agitó en un vortex (IKA – USA. Minishaker MS1S1), se dejo reposar por 3 minutos y se descartó la fase superior; este procedimiento se repitió dos veces más. El extracto se deshidrató por burbujeo con nitrógeno y se diluyó con 200 µL de H2O. b) Dansilación: 100 µL de cada muestra fue dansilada con 200 µL de cloruro de dansilo (30 mg/mL en acetona. Sigma-Aldrich – USA, Referencia 39220-5G-F) y NaHCO3 (50 mg). Luego de 12 horas en oscuridad, se adicionaron 100 µL de prolina (150 mg/mL, Sigma Aldrich - USA, Ref. P0380-100G) durante 30 min., el extracto fue secado por burbujeo con nitrógeno y diluido en 100 µL de H2O. Para extraer las aminas dansiladas se adicionó 1mL de benceno (Sigma Aldrich - USA, Ref. 270709-100ML), se agitó en vortex por 15 segundos y se centrifugó a 5.800 rpm. por un minuto. La capa de benceno (0,8mL) fue cuidadosamente removida y transferida a nuevos tubos eppenforf y las aminas dansiladas fueron conservadas a -20°C hasta su posterior análisis por CLAE. Para la curva de calibración, las poliaminas Put, Spd y Spm (Sigma Aldrich - USA, Ref. S3256-1G, S2626-1G, P7505-25G) se utilizaron para realizar soluciones patrón de 10-150 nmol/mL diluidas en metanol (Grado HPLC, Merck – Ref. MX0475-1) y 100 µL de cada solución patrón fue dansilada de manera similar a las muestras. c) Cromatografía. 100 µL de los derivados dansilados fueron ubicados en tubos eppendorf, se evaporó el benceno con la ayuda de nitrógeno y los derivados dansilados fueron diluidos en 500µL de metanol. Un volumen de 50 µL fué inyectado en el Cromatógrafo Líquido (Agilent- Canada, HP Series 1100), con una columna de fase reversa C-18 (Phenomenex – USA, Ref. 00B-4017-B0 de 50x2mm). La lectura con detector UV se realizó a los 254 nm, se usó una elusión en gradiente de metanol/agua del 65% con incrementos de metanol hasta el 85% hasta los 8 minutos, una velocidad de flujo de 0,8 mL/min. y un tiempo de corrida de 17 min.
Las variables bioquímicas fueron el contenido de Put, Spd, Spm y PAs totales y las relaciones de PAs: Put/Spd+Spm, Put/Spd, Put/Spm y Spd/Spm. Las variables morfológicas fueron el número de brotes, nudos y hojas, y la longitud promedio del brote.
Análisis estadístico: Los datos fueron analizados y graficados utilizando los programas Statistica versión 6,0 y Excel. Se realizaron pruebas de normalidad y homogeneidad de varianza, un análisis de varianza no paramétrico (Kruskal-Wallis), una prueba de Dunn para comparar los tratamientos y un análisis de correlación (coeficiente de Pearson) entre las variables bioquímicas y morfológicas. Para el análisis de correlación no se tuvo en cuenta el control pues este corresponde a yemas que no contienen tejido diferenciado.
RESULTADOS
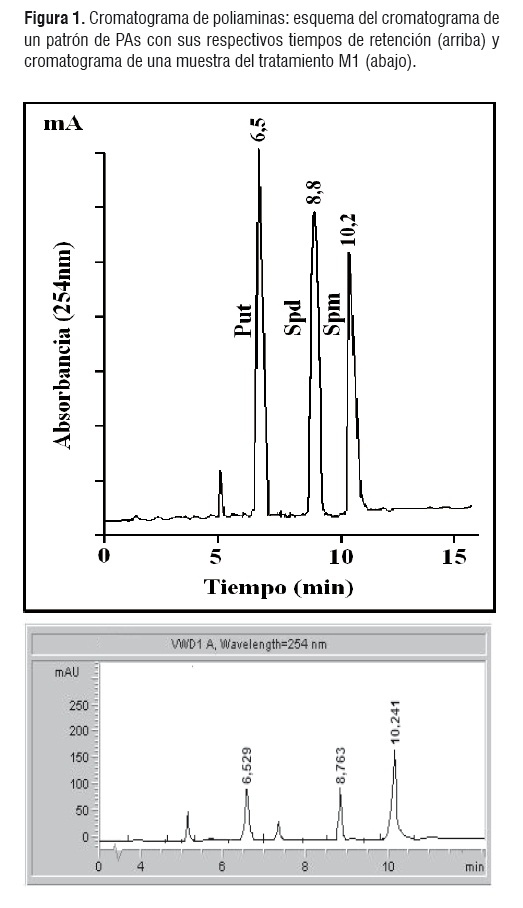
El orden en el que aparecen las PAs y sus tiempos de retención se indican en la figura 1. No se presentaron sustancias adicionales que interfirieran con los picos de las poliaminas estudiadas. El tiempo de retención de las poliaminas fue igual en muestras y patrones y su aparición en el cromatograma concuerda con su peso molecular, es decir, Put (88,15 g/mol) aparece primero porque tiene la menor masa molecular, seguido por Spd (145,2 g/mol) y Spm (202,34 g/mol).
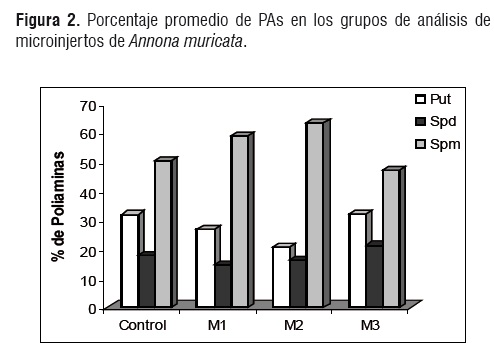
El porcentaje promedio de cada poliamina en los tratamientos se indica en la figura 2. En general, la Spm es la PA de mayor abundancia en los tejidos analizados de Annona muricata L. seguida de Put y Spd. Cuando el porcentaje de Spm aumenta, el porcentaje de Put baja indicando una relación inversa entre ellas; esta situación es explicada por la ruta biosintética de las poliaminas ya que, mediante la adición de grupos aminopropilo (provenientes de la metionina transformada) la Put se transforma en Spd y esta a su vez en Spm con la ayuda de enzimas [3,10]. Así, la producción de Spm implica un gasto de Put y la reducción en la síntesis de Spm conlleva a una acumulación de Put. Es probable que Spd funcione como puente metabólico entre la putrescina y la espermina y que su biosíntesis esté regulada por la actividad enzimática que la produce más que por la relación fuente-demanda entre poliaminas.
El análisis de varianza del contenido de poliaminas y sus relaciones en los tratamientos indicó una diferencia estadísticamente significativa (p<0.05) para casi todas las variables bioquímicas (cuadro 1). Lo anterior es una evidencia de que los tratamientos de microinjertación provocaron un cambio fisiológico en los tejidos, causado principalmente por el paso de nutrientes, reguladores de crecimiento o mensajeros secundarios desde el portainjerto, que es un material joven, hacia la estaca, desencadenando un proceso de organogénesis en los tejidos. La variable Put/Spd no presentó diferencias significativas en los tratamientos y no se incluyó en el cuadro 1.
Se puede generalizar que en las plantas superiores los niveles altos de poliaminas son característicos de tejidos meristemáticos y en crecimiento activo, y que cada órgano de la planta presenta un patrón específico, en algunas ocasiones correlacionado con procesos de división celular y en otros de expansión celular [12]. Entre los tratamientos aplicados en este estudio, el Control presenta los niveles más altos de poliaminas; esto era de esperarse porque el material vegetal analizado fueron yemas que en su interior contenían tejido meristemático, reconocido por sus altos niveles de poliaminas comparados con los de tejidos diferenciados o senescentes. Así, el tejido meristemático ubicado en las yemas de material adulto de Annona muricata L., se encuentra en un proceso de división celular, evidenciado por sus altos niveles de poliaminas.
La Spm es importante para el desarrollo vegetal de Annona muricata L., porque se encuentra en mayor concentración en todas las condiciones ensayadas. Las series de microinjertación (M1, M2 y M3) reducen significativamente el contenido de Spm y de Pas totales en Annona muricata L. Al parecer, la síntesis de Spm está relacionada con la edad del explante, siendo mayor la concentración de Spm cuando menor es la edad del brote microinjertado. Esta información corrobora el hecho de que la concentración de PAs es mayor en plantas jóvenes no senescentes y disminuye a medida que los órganos se hacen viejos [3].
Las relaciones entre las poliaminas pueden explicar en parte fenómenos de activación o inhibición de su biosíntesis. Dado que la PA precursora (Put) se encuentra en el numerador de la relación, los valores altos implican la acumulación de Put, mientras que los valores bajos implican la biosíntesis de Spm. En este estudio, se evidencia una relación de fuente-demanda entre Put y Spm respectivamente; además, las relaciones que involucran a Spm son menores de 1, indicando que no hay acumulación de Putrescina y que la síntesis de Spm es constante. La relación de fuente-demanda entre Put y Spm también fue observada por Romero y Norato [22-23].
El análisis de correlación entre las variables morfológicas y bioquímicas se indica en el cuadro 2. Las correlaciones mas altas se presentan en las variables que relacionan las poliaminas, más que en las poliaminas individuales como tal.
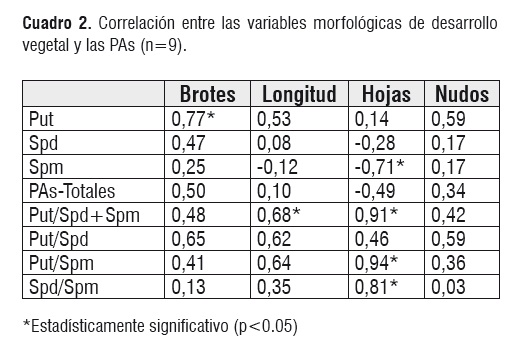
La correlación positiva entre el número de hojas y las relaciones que involucran a Spm (Put/Spd+Spm, Put/Spm y Spd/Spm) permite proponer que la dinámica metabólica de las PAs participa en la organogénesis de la hoja en Annona muricata L. La correlación negativa entre el número de hojas y los niveles de Spm está indicando que los brotes con más hojas contienen menos Spm.
El número de brotes se relaciona positivamente con los niveles de Put y sucede lo mismo entre la longitud del brote y la relación Put/Spd+Spm. Este resultado concuerda con los resultados obtenidos por otros autores sobre la importancia que Put tiene en el desarrollo vegetal 'In Vitro' de otras plantas. Rey y colaboradores investigaron el contenido endógeno de Pas en hojas de plantas jóvenes y maduras de Corulus avellana L. revigorizadas in Vitro" mediante subcultivos [24]. Encontraron que Put libre se encuentra en mayor cantidad en el tejido joven y que el tejido adulto iguala los niveles de PAs cuando se analizaron 14 subcultivos; luego de los cuales las PAs decrecen. La relación Put/Spd+Spm es mayor en el tejido joven y micro propagado que en el adulto y podría reflejar un balance entre el crecimiento vegetativo y procesos reproductivos.
Huh encontró que la Put en Dendranthema grandiflorum (crisantemo) aumenta con el incremento en la formación de yemas axilares estimulado por la temperatura. La transformación de Put a Spd y Spm podría estar relacionada en la formación de yemas axilares [25]. La revigorización de Pinus radiata investigada por Fraga esta correlacionada positivamente con el contenido de Put libre y con la relación Put/spd+spm [26]. La relación Put/spd+spm es mayor en yemas apicales de plantas jóvenes de Pinus radiata y Pinus pérsica al compararlas con las yemas de plantas adultas y la Put decrece con la edad en ambas especies [27].
En este trabajo se encontraron varias correlaciones que implican a las PAs como marcadores fisiológicos de desarrollo vegetal 'In Vitro' de Annona muricata L. La correlación de la longitud del brote con la relación Put/Spd+Spm soporta la idea de que la dinámica metabólica de las PAs, más que de sus concentraciones individuales, podría participar en la elongación celular y crecimiento de la planta.
CONCLUSIONES
La edad del material vegetal de Annona muricata L., está asociada al contenido endógeno de poliaminas libres principalmente de Spm, que es la PA más abundante en los tejidos y representa mas de la mitad del contenido total de PAs. El contenido de PAs difiere con el tipo de tejido vegetal u órgano en esta especie, así, el tejido meristemático encontrado en las yemas contiene los niveles mas altos de PAs, mientras que las hojas contienen menos Spm. En este trabajo se encontraron varias correlaciones que implican a las PAs como marcadores fisiológicos de desarrollo vegetal 'In Vitro' de Annona muricata L., entre ellos la división celular, la elongación celular y la organogénesis.
AGRADECIMIENTOS
Los autores agradecen la colaboración del Postgrado en Ciencias Biología y de Los Departamentos de Biología y Química de La Universidad del Valle; a Alba Marina Torres Ph.D. y Hernán Mauricio Romero Ph.D. por sus aportes en la investigación. Ricardo Benítez agradece a la Universidad del Cauca por su apoyo para el trabajo.
REFERENCIAS
[1] KOVÁCS, Z., SIMON-SARKADI, L., SZUCS, A. and KOCSY, G. Differential effects of cold, osmotic stress and abscisic acid on polyamine accumulation in wheat. Amino Acids., 38, 2010, p. 623–631. [ Links ]
[2] GEMICI, M., UNAL, D., AZERI, F., TAN, K. Correlation between polyamines and growth regulators. J. Food Sci, 29, 2006, p. 13-23. [ Links ]
[3] MENDOZA, C. y ROCHA, P., PAs: reguladores del crecimiento con múltiples efectos en las plantas. Palmas., 23(4), 2002, p. 39-46. [ Links ]
[4] GALIBA, G., VAGUJFALVI, A., LI CH., SOLTES Z, A. and DUBCOVSKY, J. Regulatory genes involved in the determination of frost tolerance in temperate cereals. Plant Sci., 176, 2009, p. 12–19. [ Links ]
[5] GROPPA, M.D., and BENAVIDES, M.P. Polyamines and abiotic stress: recent advances. Amino Acids., 34, 2008, p. 35–45. [ Links ]
[6] BARON, K. and STASOLLA, C., The role of polyamines during in Vivo and 'In Vitro' development. 'In Vitro' Cell. Dev. Bio. Plant., 44, 2008, p. 384-395. [ Links ]
[7] ALCÁZAR, R., MARCO, F., CUEVAS, J.C., PATRON, M., CARRASCO, F., et al. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 28, 2006, p. 1867–1876. [ Links ]
[8] TIBURCIO, A. ALTABELLA, T. and MASGRAU, C., Polyamines In: New Developments in Plant Hormone Research. Bisseling T and Schell J (Ed). Springer-Verlag, New York, 2002, p. 38-85 [ Links ]
[9] BAIS, H. and RAVISHANKAR, G., Role of polyamines in the ontogeny of plants and their biotechnological applications. Plant Cell, Tiss. and Org. Cult., 69, 2002, p. 1-34. [ Links ]
[10] EVANS, P and MALMBERG, R., Do polyamines have roles in plant development?. Plant Physiol., Annu. Rev. Plant Mol. Biol., 40, 1989, p. 235 – 269. [ Links ]
[11] RAVINDAR, K., et al. Polyamines in plants: An overview. Halic University, Printed in Turkey. Journal of Cell and Molecular Biology., 2, 2003, p. 1-12. [ Links ]
[12] FEIXA, X., PAs y Tolerancia al Aluminio en variedades tolerantes y sensibles de Zea maiz. (Tesis Doctoral). Barcelona (España): Universidad Autónoma de Barcelona, 2001, p. 20 45. [ Links ]
[13] LIU, J., et al. Production of polyamines is enhanced by endogenous abscisic acid in maize seedlings subjected to salt stress. J. Integr. Plant Biol., 47, 2005, p. 1326–1334. [ Links ]
[14] ALCÁZAR, R., et al. Polyamines: molecules with regulatory functions in plant abiotic stress tolerance. Planta., 231, 2010, p. 1237–1249. [ Links ]
[15] TROPICAL PLANT DATABASE. Graviola (Annona muricata). Raintree Nutrition. [on line]. Available: http://www.rain-tree.com/graviola.htm [Citado 14 de Diciembre de 2010] [ Links ]
[16] ÁLZATE, A., Optimización de la Metodología de propagación 'In Vitro' del guanábano desarrollada por Corporación Biotec y CIAT, como mecanismo para asegurar cultivos productivos y con frutos de óptima calidad sanitaria y organoléptica. [Informe final de pasantía Joven Investigador]. Santiago de Cali (Colombia): Convenio Colciencias – CIAT, 2002, p. 5 -19 [ Links ]
[17] PERDOMO AGUIRRE, G., Evaluación de una tecnología de microinjertación en clones promisorios de guanábana (Annona muricata L.). [Tesis]. Cali (Colombia): Universidad del Valle, 2001, p. 22 - 43 [ Links ]
[18] BRIDG, H., Micropropagation and determination of the 'In Vitro' stability of Annona cherimoya Mill. and Annona muricata L. Berlin, Humboldt-University. Landwirtschaftlich-Gärtnerische Fakultät, Diss. (on line). Available: http://edoc.hu-berlin.de/dissertacionen/bridg-hannia. [Citado 9 de Diciembre de 2010] [ Links ]
[19] RINCÓN, A., et al. Establecimiento aséptico de brotes laterales de Annona spp. Rev. Fac. Agron. (LUZ)., 16 (1), 1999, p. 76-81. [ Links ]
[20] ACOSTA, A.M., PEÑA E.J., y CASTRO D. Evaluación de medios de cultivo para la producción in vitro de Annona muricata mediante la técnica de microinjertación seriada. Acta Agronómica Vol 60, No 2 (2011). [ Links ]
[21] ALDESUQUY, H., ABDEL-FATTAH, G., and BAKA Z., Changes in chlorophyll, polyamines and chloroplast ultrastructure of Puccinia striiformis induced green islands on detached leaves of Triticum aestivum. Plant Physiol. Biochem., 38, 2000, p. 613-620. [ Links ]
[22] ROMERO, H y NORATO, J., Estudios fisiopatológicos en palma de aceite: I. Contenido de PAs libres y su relación con la pudrición del cogollo., Revista Comalfi, 26(1,3), 1999, p. 1-7. [ Links ]
[23] NORATO, J y ROMERO, H ¿En las plantas que sufren los efectos de las heladas las PAs conforman algún mecanismo que les permita sobrevivir?. Revista Comalfi., 22 (3), 1995, p. 19-23. [ Links ]
[24] REY, M., DÍAZ-SALA, C., y RODRIGUEZ, R., Free polyamine content in leaves and buds of hazelnut (Corylus avellana L. cv. Negret) trees subjected to repeated severe pruning. Scientia Horticulturae., 76, 1998, p. 115-121. [ Links ]
[25] HUH, Y., et al., Effect of Temperature on Axillary Bud Formation and Polyamine Contents of Nonbranching Chrysanthemum. Hort Science., 40, 2005, p. 993-1147. [ Links ]
[26] FRAGA, M., RODRÍGUEZ, R., and CAÑAL, M., Reinvigoration of Pinus radiata is associated with partial recovery of juvenile-like polyamine concentrations. Tree Physiol. 23(3), 2003, p. 205-9. [ Links ]
[27] FRAGA, M., et al., Changes in polyamine concentration associated with aging in Pinus radiata and Prunus persica. Tree Physiol., 24(11), 2004, p. 1221-6. [ Links ]
