










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiotecnología en el Sector Agropecuario y Agroindustrial
Print version ISSN 1692-3561
Rev.Bio.Agro vol.10 no.2 Popayán July/Dec. 2012
VARIABILIDAD GENÉTICA DEL ROBLE COMÚN (Quercus humboldtii BONPL.) EN LA REGIÓN DEL MACIZO COLOMBIANO
VARIABILIDADE GENÉTICA DO ROBLE COMUM (Quercus humboldtii BONPL.) NA REGIÃO DO MACIZO COLOMBIANO
GENETIC VARIABILITY OF COMMON OAK (Quercus humboldtii BONPL.) IN THE MACIZO COLOMBIANO REGION
GISELA MABEL PAZ P.1
1 Magíster. Universidad del Cauca.
Correspondencia: gmabel@unicauca.edu.co
Recibido para evaluación: 13/09/2011 Aprobado para publicación: 18/06/2012
RESUMEN
Los impactos genéticos de la fragmentación de las poblaciones dependen, entre otros factores, del flujo de genes. Con un flujo restringido de genes, la fragmentación aumenta la probabilidad de homocigosis y la pérdida de la diversidad genética. Estas poblaciones involucran algunas especies como el Quercus humboldtii (roble). Para estimar los niveles de diversidad genética en 120 individuos de cuatro poblaciones en el Macizo Colombiano (Andes Colombianos) se utilizaron seis cebadores de marcadores RAPDs. Se obtuvo un total de 123 loci en todas las poblaciones, 122 de los cuales fueron polimórficos. La diversidad genética fue de 0,4583 y el valor del estadístico de estructura genética (GST) fue de 0,1616 con un valor de flujo genético de 2,58. Basados en los resultados, se indica que existe un grado de estructuración genética entre las poblaciones y que para el mantenimiento y la conservación del germoplasma se recomienda: si el objetivo es conservar una unidad panmíctica, se requiere generar corredores biológicos; pero si se quiere mantener la actual diferenciación no habría necesidad de la creación de los mismos pero si de planes de reforestación locales.
PALABRAS CLAVE: Diversidad genética, Estructura genética, Genética de la conservación.
ABSTRACT
Genetics impacts of fragmentation in populations depend, among other factors, of gene flow. With a restricted gene flow, fragmentation increases the probability of homozygosity and the loss of genetic diversity. These populations involve some species such as Quercus humboldtii (oak). To measure the levels of genetic diversity in 120 samples from 4 different populations in the Colombian Andean region (Macizo Colombiano), six primers of RAPDs markers were used. A total of 123 loci were obtained in all the populations, 122 of them were found to be polimorphic. The genetic diversity was of 0.4583 and the statistic value of genetic structure (GST) was of 0.1616 with a genetic flow value of 2.58. Based on the results, it was underlined that there is a level of genetic structuration among the populations and for its sustenance and preservation of germplasm, it is reccomended: if the objective is to keep a panmictic unit, it is required to generate biologic corridors; but if it is required to maintain the current differentiation it wont be necessary the creation of these corridors but the creation of plans of local reforestation.
KEY WORDS: Genetic diversity, Genetic structure, Conservation genetics.
RESUMO
Os impactos genéticos da fragmentação das populações dependem, entre outros fatores, do fluxo de genes. Com um fluxo de genes restringido, a fragmentação aumenta a probabilidade de homozigose e a perda da diversidade genética. Estas populações envolvem algumas espécies como o Quercus humboldtii (carvalho). Para estimar os níveis de diversidade genética em 120 indivíduos de quatro populações no Macizo Colombiano (Andes Colombianos), se empregaram seis primers marcadores RAPDs. Obteve-se um total de 123 loci em todas as populações, 122 das quais foram polimórficas. A diversidade genética foi de 0,4583 e o valor do estatístico de estrutura genética (GST) foi de 0,1616 com um valor de fluxo genético de 2,58. Baseado nos resultados sugere-se que existe um grau de estruturação genética entre as populações, e para manter e conservar o germoplasma recomenda-se conservar uma unidade panmítica, o qual requer a geração de corredores biológicos ou pelo menos para manter a atual diferenciação, precisa-se de planos de conservação de áreas naturais locais.
PALAVRAS-CHAVE: Diversidade genética, Estrutura genética, Conservação genética.
INTRODUCCIÓN
El Macizo Colombiano es una extensa región del sur occidente de Colombia que por su ubicación estratégica reúne una serie de condiciones biológicas, hídricas y geográficas que la hacen importante. En términos biológicos, es una zona reconocida por la diversidad de ecosistemas y la gran diversidad a nivel de especies representada en diferentes grupos biológicos. En términos hídricos, en ella se encuentra la estrella hidrográfica donde nacen ríos tan importantes como Magdalena, Cauca, Caquetá, Putumayo y Patía y por último, en términos geográficos, es precisamente en esta zona donde la cordillera de los Andes después de atravesar todo Suramérica se divide en tres ramales, situación que genera aislamiento y este a su vez innumerables consecuencias biológicas. Por estas razones, el Macizo Colombiano es una extensa zona estratégica que necesita ser estudiada para garantizar su conservación.
Quercus humboldtii (roble), pertenece a la familia de las Fagáceas, su polinización es 100% por el viento y sus semillas son dispersadas por roedores, la gravedad o aves como el pájaro carpintero [1]. El roble, presenta una amplia distribución en la zona andina, donde dominan las vertientes internas de las tres cordilleras entre los 1.600 y los 3.000 m.s.n.m [2,3,4,5,6,7,8]. Se desarrolla a una temperatura entre 10 y 17°C, con una precipitación de 1.300 a 3.000 mm/año. Su amplia distribución altitudinal trae como consecuencia que la especie muestre evidentes cambios morfológicos. Esta gran variabilidad morfológica es responsable de que en el pasado se hayan descrito 7 especies del género Quercus en Colombia (Q. humboldtii H & B; Q. tolimensis H & B; Q. almaguerensis H & B; Q. lindenii De Candolle; Eritrobalanus (Quercus) duqueana Schwartz; Q. colombiana Cuatrecasas y Q. boyacensis Cuatrecasas). Sin embargo, Müller [9] en la revisión del género Quercus reconoce para Colombia solo la especie Quercus humboldtii, agrupando en ésta las anteriormente descritas. Posteriormente, esta agrupación fue ratificada por estudios morfológicos y moleculares [10].
Q. humboldtii, es una especie endémica de Colombia y está catalogada en el libro rojo de plantas maderables de Colombia, como vulnerable (VU) [11]; esta categoría se asigna porque la especie tiene una extensión nacional inferior a 20.000 km2, un área de ocupación inferior a 2.000 Km., una población de individuos maduros menor a 1000, está presente únicamente en 10 localidades y presenta una reducción obvia (observada, estimada, inferida o sospechada) en los últimos 10 años o de tres generaciones, mayor al 50%. En nuestro estudio realizado en los municipios de Almaguer y la Vega, considerada una localidad tipo, en razón a la determinación del Q. almaguerensis encontrado en esta región del Macizo colombiano, se realizó un censo en nueve (9) veredas dentro de las cuales se encontraron cuatro (4) poblaciones de roble y se evidenció la fragmentación de su hábitat por la deforestación para la utilización de la madera y las actividades agrícolas de sus pobladores.
El conocimiento de la cantidad y distribución de la variación genética dentro y entre poblaciones es fundamental para la conservación de los recursos genéticos por varias razones: primero, el cambio del medio ambiente es un proceso continuo y se espera que especies con una alta diversidad genética, puedan adaptarse mejor a las condiciones adversas y segundo, conociendo la diversidad genética de las especies, se pueden realizar procesos de repoblamiento antes que desaparezcan. De otro lado, la diversidad genética está en función de la distancia geográfica, es decir, las poblaciones ampliamente separadas, estarán más diferenciadas que las poblaciones más cercanas geográficamente y en poblaciones fragmentadas, la diversidad genética será menor dentro de los fragmentos, como se ha demostrado en varias especies tropicales [12]. Existen otros factores que inciden en el comportamiento genético de las especies, como por ejemplo: el sistema reproductivo que controla como los gametos se unen para formar la progenie, la dispersión de esta progenie, la variación en el conjunto de genes, la organización de esta variación en genotipos, la distribución espacial de estos genotipos, eventos al azar, y el proceso de crecimiento, mortalidad y reemplazo que van a originar poblaciones futuras [1,13].
Los marcadores RAPD (Polimorfismo de ADN amplificados al azar) [14], han sido utilizados para conocer la diversidad genética de las especies, porque pueden originar información de un gran número de loci. Estudios anteriores en otras poblaciones de Q. humboldtii en áreas protegidas utilizando esta técnica molecular han revelado niveles de diversidad y estructura genética similares o superiores a otras especies de roble de climas templados [15]. Fernández y Sork [16,17], utilizando marcadores microsatélites, encontraron que como consecuencia de la fragmentación de los bosques de roble se altera el flujo de polen, razón por la cual se puede ver afectada la diversidad genética al cabo de pocas generaciones.
En el presente estudio, marcadores genéticos RAPDs, se utilizaron para determinar la variabilidad y la estructura genética de cuatro poblaciones naturales de Q. humboldtii ubicados en la región del Macizo Colombiano. Los resultados pueden ser utilizados para proveer algunas recomendaciones en la elaboración de estrategias de conservación y cultivo en esta especie.
MÉTODO
La región de estudio comprende nueve veredas de la región del Macizo Colombiano: La Zanja, Dominguillo, Guambial, Riñonada, Buenavista, El Jordán, Julián, Ledezma y Chorrillos donde se encontraron cuatro poblaciones de Q. humboldtii en bosques fragmentados, ubicados en las veredas de la Riñonada, Buenavista, la Zanja y el Jordán.
Se seleccionaron cuarenta árboles de cada población, los cuales fueron marcados con placas de aluminio, se cortó una hoja joven de cada árbol y se enfrió en un contenedor de nitrógeno líquido para transportarlas al laboratorio, donde se realizaron los estudios genéticos.
Aislamiento de ADN: El ADN total fue extraído de cada individuo macerando la muestra del tejido en nitrógeno liquido, siguiendo las instrucciones del Kit de Quiagen (Dneasy plant Mini Kit, cat No.69104) para la extracción de ADN de plantas, con la cual se obtuvo el ADN observado en un gel al 0,8% de agarosa, teñido con bromuro de etidio, luego se llevó a una concentración final de 5 ng/ul para la reacción de PCR (Reacción en Cadena de la Polimerasa) y se guardó a –80°C para el análisis posterior de variabilidad genética.
Técnica RAPDs: Las reacciones de PCR (reacción en cadena de la polimerasa) se llevaron a cabo utilizando 6 cebadores reportados como polimórficos para Q. humboldtii [15], con las siguientes condiciones: un volumen final de 25 µl con 2,5 µl de buffer para PCR 10X que contenía 2mM de MgCl2, 0,25 mM de dNTPs, 0,8 µM de cebador, 1 U de Taq polimerasa y 5 µl de ADN.
La PCR se realizó en un termociclador PTC-100 (MJ Research Inc) siguiendo el siguiente programa: desnaturalización inicial a 94°C durante 3 minutos, alineamiento a 38°C durante 30 segundos, extensión a 72°C durante 1 minuto, seguido por 40 ciclos de desnaturalización a 94°C por 30 segundos, una extensión final a 72°C por 5 minutos, seguido por un enfriamiento a 4°C.
Los productos de la amplificación fueron ubicados en geles de agarosa al 1,5% teñidos con bromuro de etidio y para determinar el tamaño de los loci se utilizó un marcador de peso molecular Generuler DNA ladder Mix de Fermentas. Los geles fueron visualizados en un transiluminador con luz ultravioleta y fotografiados para su lectura posterior.
Análisis de los datos
A partir de los geles, se realizó una matriz de presencia (1) y ausencia (0), con la cual se calculó la diversidad genética en términos de Índice de Shannon, estructura genética en términos de GST, identidad genética y distancia genética de Nei [18], con el método UPGMA, estructura poblacional en términos de Θ, diversidad genética [19], análisis multidimensional de correspondencia múltiple (AMC), utilizando para ello el programa TFPGA [20].
Para complementar los análisis de estructura genética se realizaron análisis de agrupamiento a partir de matrices de similitud. Con la matriz de datos presencia/ausencia se calculó el índice de similitud desarrollado por Dice [21] y adaptado por Nei y Li, [22], para datos moleculares. Las matrices y los dendrogramas de similitud fueron calculados utilizando los programas Tools for population genetic analyses (TFPGA) 1.3. [20].
RESULTADOS
Con los seis cebadores evaluados se obtuvo 122 loci con un rango de tamaños entre 300 a 3000 pb y un número promedio de bandas por cebador de 20,5. El porcentaje promedio de loci polimórficos para las cuatro poblaciones de Q. humboldtii fue de 99,18% (Cuadro 1).
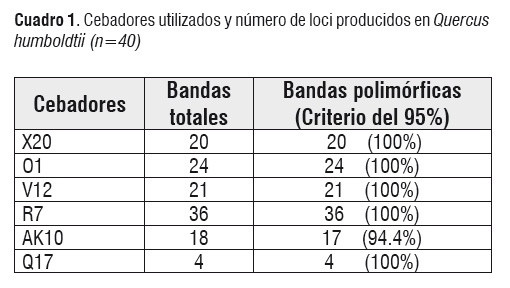
La diversidad genética total para todos los loci considerando como una sola unidad panmíctica a las cuatro poblaciones fue de 0,4583. Para cada población, los valores de diversidad genética oscilaron entre 0,3876 para la población de la Riñonada, y 0,3776 para la población de la Zanja (Cuadro 2). Se observa que la diversidad genética total es mayor que el rango en que esta la diversidad por población. Esto puede ser un indicador de estructura poblacional, es decir a pesar de que los valores de diversidad por población son similares, existen diferencias en las frecuencias de los alelos de los loci estudiados.
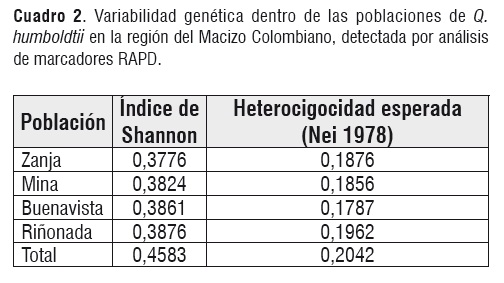 Cuadro 2
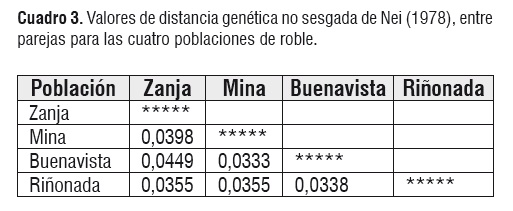
Cuadro 2Para confirmar lo anterior, se hizo un análisis de estructura poblacional, estimando el valor de valor de Θ como 0,1373 ± 0,0202. Debido a que el error estándar es bajo se espera que los valores de Θ por parejas oscilen alrededor de este valor. Considerando las cuatro poblaciones, el valor estimado del GST fue de 0,1616, y el número estimado del flujo de génico fue de 2,58. Estos valores intermedios indican que efectivamente existe un grado significativo de estructuración y por lo tanto, no se pueden considerar las cuatro poblaciones como una sola unidad poblacional (Cuadro 3).
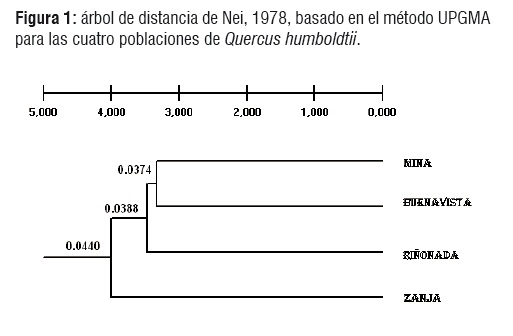
Se construyó un dendrograma con el método UPGMA, basado sobre la distancia genética de Nei (Figura 1), en el cual se reporta una mayor similitud entre las poblaciones de Mina y Buenavista, quedando organizadas en un mismo grupo y estas dos a su vez, se separan de la población de la riñonada en un nuevo nodo y en el último nodo aparece la población de la Zanja mas diferenciada. Esta diferencia posiblemente se debe a que la población de la Zanja esta mas aislada geográficamente de las demás poblaciones.
El análisis multidimensional de correspondencia múltiple, muestra las relaciones existentes entre los individuos de las diferentes poblaciones (Figura 2).
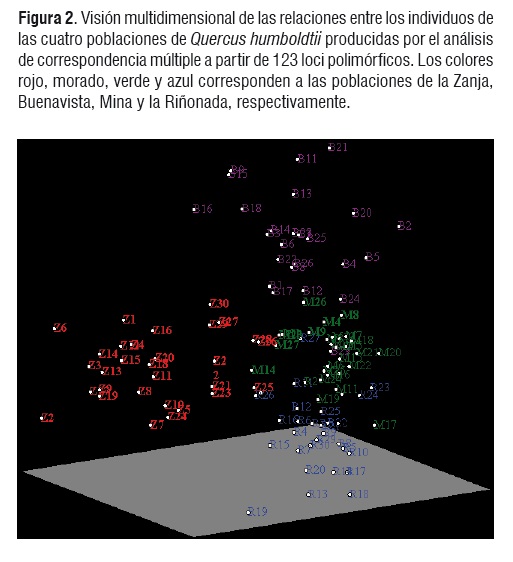
Claramente se observa que los individuos de la Zanja y de Buenavista, se separan del resto de las poblaciones, con poca superposición de individuos, lo cual también soporta la subdivisión poblacional mencionada anteriormente.
Discusión
Variación genética. Las cuatro poblaciones, muestran niveles similares de variación genética con un rango muy estrecho de variación (Cuadro 2).
Las plantas de la población de la Riñonada, exhibieron los más altos niveles de variación genética (I=0,3876), en contraste, la población de la Zanja, presenta los menores niveles de diversidad (I=0,3776). Estos resultados indican que la diversidad genética de los bosques fragmentados tropicales de roble, es menor que la de bosques continuos. (Ho=0,813, Φ=0,015). [16].
Palacio, 2005 [15] en trabajos de diversidad genética en tres poblaciones de robles, también encontró estimaciones de diversidad genética menores que la reportada para bosques continuos (H=0,5379) y valores de estructura genética similares a los estimados en esta investigación (ΦST=0,205).
Se considera que existe un grado intermedio de estructuración genética entre las cuatro poblaciones de roble encontradas en la región de Macizo Colombiano, esto se confirma con el flujo de genes de 2,58 el cual, a largo plazo seguramente podría aumentar la deriva genética o procesos de autopolinización entre los individuos. En futuros estudios se podría analizar las características reproductivas de las poblaciones, como por ejemplo el grado de autopolinización y el número de donadores de polen, ya que estos cambios afectan directamente a las oportunidades de apareamiento entre los individuos.
En el presente estudio, se utilizaron marcadores genéticos RAPDs, para determinar la variabilidad genética y la estructura genética de cuatro poblaciones naturales de Q. humboldtii ubicados en la región del Macizo Colombiano para la elaboración de estrategias de conservación y cultivo para esta especie. Según los resultados de investigación se debe tener en cuenta que el repoblamiento debe ser local, debido a la diferenciación genética existente entre las poblaciones. Los valores de diferenciación encontrados parecen indicar que esta separación es relativamente reciente ocasionada posiblemente por la fragmentación del hábitat (figura 3), por lo tanto, según este estudio si el objetivo es conservar una unidad panmíctica se requiere generar corredores biológicos, pero si se quiere mantener la actual diferenciación no habría necesidad de la creación de los mismos pero si de planes de reforestación locales.

AGRADECIMIENTOS
A la Universidad del Cauca, Al Fondo para la Acción Ambiental, por el apoyo financiero en el proyecto, al Dr. Eiber Cardenas, por su apoyo en el análisis de los datos, A Juan Diego Palacio por el préstamo de los equipos y experiencia en el laboratorio de Biotecnología del Instituto de investigación de recursos biológicos Alexander von Humboldt.
REFERENCIAS
[1] KATTAN, G. Food habits and social organization of acorn woodpeckers in Colombia. Condor 90(1): 100-106. 1988. [ Links ]
[2] CUATRECASAS, J. Aspectos de la vegetación natural de Colombia. Revista de la Academia Colombiana de Ciencias 1958. 10: 221-268. [ Links ]
[3] ESPINAL, L. S. Algunos aspectos de la vegetación del oriente antioqueño. Bogotá, Instituto Geografico Agustín Codazzi. 1964. [ Links ]
[4] ESPINAL, L. S. Geografía ecológica de Antioquia. Medellín, Universidad Nacional de Colombia. 1992. [ Links ]
[5] ESPINAL, L. S. Regiones de Colombia. Medellín, Universidad Nacional de Colombia. 1993. [ Links ]
[6] PACHECO, R. A. and C. A. PINZÓN. Notas divulgativas: El Roble (Quercus humboldtii Bonpland). Santa Fe de Bogotá, Jardín Botánico de Bogotá José Celestino Mutis. 1997. [ Links ]
[7] RANGEL, J. O. Colombian Biome Meeting. Vegetation of Colombia: reconstruction implications in the global and local changes, Bogotá. 2000. [ Links ]
[8] CALDERÓN, E. Plantas colombianas en peligro, extintas o en duda. Instituto Humboldt. 2001. [ Links ]
[9] MÜLLER, C. H. The Central America species of Quercus. U.S. Depto. Agric. Misc. Publ. 477: 1-216. 1942. [ Links ]
[10] CAVELIER, J., T. AIDE, et al. Especiación del género Quercus (robles) en Colombia: Un siglo y medio de incertidumbre. Fondo FEN: 39. Bogotá, D. C., 1995. [ Links ]
[11] CARDENAS L.,D., SALINAS, N. R., (ed.) Libro rojo de plantas de Colombia, Volumen 4, especies maderables amenazadas, primera parte. Instituto Amazónico de Investigaciones Científicas SINCHI. – Ministerio de Ambiente, Vivienda y Desarrollo Territorial. 2007. 232 pp. [ Links ]
[12] PAUTASSO, M. Geographical genetics and the conservation of forest trees. Perspectives in Plant Ecology, Evolution and Systematics 11:157–189. 2009. [ Links ]
[13] LEVIN,D.A. Some genetic consequences of being a plant. In: Brussard, P., (ed.), Ecological Genetics: The inerface. Pp. 189-212. Springer-Verlag, N,Y. 1978. [ Links ]
[14] WILLIAMS, J. G. K., A. R. KUBELIK, y col. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research 18(22): 6531-6535. 1990. [ Links ]
[15] PALACIO-MEJÍA, J. D. Estudio comparativo de la diversidad genética y divergencia evolutiva entre la especie vulnerable Colombobalanus excelsa (Lozano et al.) Nixon & Crepet y el roble común Quercus humboldtii Bondpl.: implicaciones para la biología de la conservación. Escuela de posgrados. Palmira, Universidad Nacional de Colombia, Sede Palmira: 108. 2005. [ Links ]
[16] FERNANDEZ-M, J.F.AND V.L. SORK. Genetic variation in fragmented forest stands of the Andean oak Quercus humboldtii Bonpl. (Fagaceae) Biotropica 39: 72-78.2007. [ Links ]
[17] FERNÁNDEZ-M., J. F. AND V. L. SORK. Mating Patterns of a Subdivided Population of the Andean Oak (Quercus humboldtii Bonpl., Fagaceae). Journal of Heredity 96(6): 635-643. 2005. [ Links ]
[18] NEI, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89: 583-590. 1978. [ Links ]
[19] NEI M. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences, USA 70: 3321-3323. 1973. [ Links ]
[20] MILLER, P. M. Tools for population genetic analyses (TFPGA) 1.3. A Windows program for the analysis of allozyme and molecular population genetic data. Computer software distributed by author. 1997. [ Links ]
[21] DICE, L. R. Measures of the amount of ecological association betweem species. Ecology 26: 297-302.1945. [ Links ]
[22] NEI, M. and W.-H. LI. Mathematical Model for Studying Genetic Variation in Terms of Restriction Endo Nucleases. Proceedings of the National Academy of Sciences of the United States of America 76(10): 5269-5273. 1979. [ Links ]
