










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiotecnología en el Sector Agropecuario y Agroindustrial
Print version ISSN 1692-3561
Rev.Bio.Agro vol.10 no.2 Popayán July/Dec. 2012
AISLAMIENTO E IDENTIFICACIÓN DE BACTERIAS SOLUBILIZADORAS DE FOSFATOS, HABITANTES DE LA RIZÓSFERA DE CHONTADURO (B. gassipaes Kunth)
ISOLATION AND IDENTIFICATION OF PHOSPHATE-SOLUBILIZING BACTERIA INHABITING THE RIZHOSPHERE OF PEACH PALM (B. gassipaes Kunth)
ISOLAMENTO E IDENTIFICAÇÃO DE BACTÉRIAS SOLUBILIZADORAS DA FOSFATO, HABITANTES DA RIZÓSFERA DE CHONTADURO (B. gassipaes Kunth)
1 Ph.D. Docente auxiliar ECAPMA – Universidad Nacional Abierta y a Distancia UNAD
2 Ph. D. Profesora titular Universidad Nacional de Colombia sede Palmira.
Correspondencia: cpatinot@yahoo.com
Recibido para evaluación: 25/01/2012. Aprobado para publicación: 29/07/2012
RESUMEN
El chontaduro o pejiyabe (Bactris gasipaes Kunth) esuna especie nativa de las selvas húmedas del pacífico colombiano, adaptada a condiciones de alta acidez y deficiencia de nutrientes, especialmente fósforo (P), características dominantes en los suelos de esta zona tropical. Los microorganismos rizosféricos solubilizadores de P, entre ellos las bacterias (BSP), contribuyen a mejorar la disponibilidad de este nutriente, por ello, la necesidad de aislarlos, reconocerlos e identificarlos en la rizosfera, como uno de los mecanismos de adaptación de esta especie. Una vez aisladas y purificadas en medio libre de fuentes de fósforo orgánico e inorgánico solubles, las poblaciones bacterianas se identificaron mediante la utilizacióndel análisis de las secuencias DNAr 16S y la técnica BOX-PCR, estableciendo que en el sitio de estudio, las poblaciones dominantes fueron Burkholderia ambifaria y Burkholderia sp. 383, seguidas por Pseudomonas putida, especies reconocidas globalmente por su actividad solubilizadora, y como rizobacterias promotoras del crecimiento vegetal (PGPR).
PALABRAS CLAVE: Solubilización de fosfatos, Fósforo del suelo, PGPR, Biofertilizantes.
ABSTRACT
The chontaduro or pejiyabe (Bactris gasipaes Kunth), is a species native of rainforest of Colombian's Pacific Coast, adapted to soils with high acidity and depleted in nutrients, especially phosphorus.The phosphate solubilizing microorganisms, including bacteria (BSP), improve the availability of this nutrient, therefore, the need to isolate, recognize and identify those in the rhizosphere, as one of the mechanisms of adaptation of this species. Once isolated and purified in medium free of sources of soluble phosphorus, bacterial populations were identified using sequence analysis of 16S rDNA and molecular genotyping by BOX-PCR, stating that in the study site, the dominant populations were Burkholderia ambifaria and B. sp. 383, followed by Pseudomonas putida, species recognized globally for its solubilizing activity, and as plant growth promoting rhizobacteria (PGPR).
KEY WORDS: Phosphate solubilization, Soil phosphorus, Rhizosphere.
RESUMO
Do chontaduro oupejiyabe (Bactris gasipaes Kunth) é uma espécie nativa da florestas úmidas do pacífico colombiano, adaptados às condições de alta acidez e deficiência de nutrientes, especialmente fósforo (P), características dominantes no solo de esta zona tropical. Microrganismos do solo comcapacidade de solubilizar fosfatos naturais, incluindo as bactérias, contribuir para melhorar a disponibilidade de nutrientes presente, por conseguinte, não existe necessidade de isolar as, reconhecer e identificar-los na rizosfera, como um dos mecanismos de adaptação do presente espécies. Uma vez que tenham sido isoladas e purificadas emmeiolivre de fontes de P orgânicos e inorgânicos solúveis, populações bacterianas foram identificadas pela utilização da análise das sequências do extrachromosome DNAr 16S e a técnica BOX-PCR, que estabelece que a área de estudo, a populações dominante foram: Burkholderia ambifaria e B. sp. 383, seguido por Pseudomonas putida, espécies reconhecidas mundialmente para suaatividade solubilizadora, e como rizobactérias promotoras de crescimento de plantas (RPCP).
PALAVRAS-CHAVE: Solubilização de fosfatos, Fósforo do solo, RPCP, Biofertilizantes.
INTRODUCCIÓN
En el mundo, los suelos ácidos ocupan cerca de 3.95 billones de hectáreas, correspondientes al 30% de la superficie terrestre [1, 2], mientras en Colombia, representan más del 80% del territorio [3]. En general, estos suelos son deficientes en fósforo (P) disponible para los cultivos, debido a su elevada capacidad de fijación, determinada por: a) los altos contenidos de hierro y aluminio, los cuales al combinarse con el P precipitan como sales insolubles, y b) por la presencia de hidróxidos y sesquióxidos, sobre cuyas superficies se adsorbe el elemento.
La baja disponibilidad de P en el suelo conlleva una mala nutrición de las plantas, por cuanto el nutrientese suple normalmente con la aplicación de fertilizantes de síntesis química y roca fosfórica. Sin embargo, los impactos ambientales deésta práctica y el agotamiento inminente de las reservas globales de la roca fosfórica [4, 5], son razones que impulsan la búsqueda de alternativas sostenibles que permitan satisfacer los requerimientos de P de los cultivos en la agricultura moderna.
En el trópico, a pesar de la baja disponibilidad del P, los ecosistemas naturales mantienen una enorme y diversa biomasa vegetal, que sugiere la existencia de mecanismos capaces de regular el ciclaje biogeoquímico de los nutrientes, en particular el P. Tales mecanismos incluyen las funciones metabólicas de los icroorganismos del suelo y/o de las mismas plantas, a través de interacciones complejas que ocurren en la rizósfera [6, 7, 8]. De hecho, es amplio el registro de la diversidad de bacterias [9], incluidasactinobacterias [10], y de hongos [11], asociados o no a las raíces de las plantas, que hacen disponibles las diferentes formas químicas del P del suelo mediante procesos de solubilización de las fuentes minerales y/o mineralización de las formas orgánicas [11, 12].
Desde 1903, cuando se registró que algunos microorganismos tenían capacidad de solubilizar el P en el suelo y se sugirió que ellos podrían jugar papel importante en la nutrición fosfórica de las plantas, se han hecho numerosos avances en la comprensión de los mecanismos responsables del proceso y en el conocimiento de la biodiversidad microbiana con tal capacidad [1, 11, 12]. Actualmente, se considera la solubilización de fosfatos como una característica clave de los microorganismos promotores del crecimiento vegetal (PGPR).
En relación con los mecanismos, lamineralización del P orgánico depende de la secreción de enzimas fosfatasas (principalmente fitasas), mientras la solubilización a partir de fuentes minerales como fosfatos mono y dibásicos (especialmente del tipo de las apatitas), está ligada a la producción de ácidos inorgánicos, ácidos orgánicos y/o a la liberación de protones, productos del metabolismo primario del organismo solubilizador [1, 7].
Como tecnología aplicada, las primeras aplicaciones masivas en campo de inoculantes con capacidad solubilizadora de P se efectuaron en la antigua Unión Soviética en los años cincuenta, con resultados sorprendentes. La aplicación de kaolinita impregnada con esporas de Bacillus megaterium var.phosphaticum llevó a incrementos de hasta 70% en la producción de los cultivos. Con la aplicación de fosfobactericina, nombre dado a este bioinoculante, también se obtuvieron resultados positivos en suelos de la India deficientes en P, pero fueron negativos en los Estados Unidos [1]. En Cuba, Fosforina® es un bioinoculante a base de Pseudomonas fluorescens aplicado principalmente en tomate [13]. En Colombia, actualmente se comercializa Fosfosol®, producto de la investigación que sobre microorganismos solubilizadores de P (MSP) se hace en la Universidad Nacional de Colombia sede Bogotá, cuyo ingrediente activo es Penicillium janthinellum. Está dirigido especialmente al cultivo del arroz, produciendo incrementos del rendimiento entre el 5 y el 38% con respecto a cultivos no inoculados [14].
No obstante, y a pesar de las potencialidades ambientales y económicas implícitas enel uso de los MSP como biofertilizantes, aún son escasos los estudios básicos que respalden la tecnología, especialmente en el trópico [1, 13]. Tampoco es amplia su adopción por parte de los productores, campesinos y técnicos.
El presente trabajo busca contribuir al conocimiento de la diversidad bacteriana solubilizadora de P asociada a la rizósfera del chontaduro, cultivo de importancia en la región tropical del Pacífico colombiano. Las posibilidades de establecimiento de un proceso biotecnológico a partir del conocimiento de las BSP, requiere que se genere información acerca de la eficiencia, estabilidadgenotípica de los microorganismos objeto de estudio y su identificación filogenética. Por ello, la evaluación de estas tres variables, bajo condiciones in vitro, constituyen el objetivo de este trabajo, como paso previo a la estimación de su eficacia en condiciones de invernadero y campo.
MÉTODO
A partir de las raíces de cuatro plantas de chontaduro (B. gassipaes Kunth)en edad productiva, seleccionadas aleatoriamente y crecidas bajo condiciones de bosque natural en un suelo del corregimiento del Bajo Calima, en el municipio de Buenaventura (Valle del Cauca), se tomaron submuestras de suelo rizosférico (partículas adheridas al rizoplano), las cuales se mezclaron y homogenizaron para conformar una muestra de 200 g de suelo, recogida en bolsas plásticas, almacenada en nevera de icopory transportada al Laboratorio de Microbiología de la Universidad Nacional de Colombia sede Palmira.
En agua destilada estéril se efectuaron diluciones seriales de la muestra de suelo hasta 10-5. En medio de cultivo Pikovskaya (PVK) (en g L-1: glucosa, 10; Ca3(PO4)2, 5; (NH4)2SO4, 0,5; NaCl, 0,2; MgSO4,7H2O, 0,1; KCl, 0,2; extracto de levadura, 0,5; MnSO4,H2O, 0,002; FeSO4,7H2O, 0,002; agar, 15), a partir de las diluciones 10-3 a 10-5s e hicieron aislamientos por estría sobre 4 placas de Petri/dilución.El halo transparente que se formó alrededor de las colonias, en la medida que ocurrió la solubilización, constituyó el indicador de actividad. Los aislamientos obtenidos se purificaron y conservaron sobre el mismo medio, para análisis posteriores.En todos los casos, los cultivos microbianos se mantuvieron en incubadora a 28°C y luz artificial.
Índice de eficiencia solubilizadora de fosfatos (ESF)
Esta variable hace referencia a la capacidad relativa de los microorganismosde utilizar como sustrato las fuentes insolubles de este nutriente, tornándolas disponibles en el medio. Para ello,los aislamientos obtenidos, se evaluaron a los cinco días después de sembrados. La medida del índice ESF se hizo sobre 4 replicaciones por cada aislamiento. El índice ESF se calculó por la relación (11) (Ec. 1):

Estabilidad de la capacidad solubilizadora
Para verificar la permanencia en el tiempo de la capacidad solubilizadora, cada uno de los aislamientos obtenidos, se sometió a cinco resiembras consecutivas en PVK y evaluación de ESF en cada etapa (11). Los tratamientos generados a partir de cada uno de los aislados, se dispusieron en un diseño completamente al azar. El ANAVA se efectuó en el programa IBM SPSSStatistics 19. Para la comparación de medias se utilizó la prueba Tukey (α=0,05).
Identificación filogenética de los aislamientos
Se hizo a través del análisis de las secuencias del gen ADNr 16S: Para el aislamiento y purificación del ADN genómico de los aislados se utilizó el Wizard® Genomic DNA Purification Kit (Promega,USA), utilizando los protocolos sugeridos por el fabricante. El gen ADNr 16S en cada uno de los aislados se amplificó con los primer bacterianos universales fD1 y rD1 (15):fD1: (5'-CCGAATTCGTCGACAACAGAGTTTGATCCTGGCTCAG-3'), y, rD1: (5'-CCCGGGATCCAAGCTTAAGGAGGTGATCCAGCC-3').
En todos los casos, la PCR fue efectuada en un volumen de reacción de 15 µl, conteniendo: 1 µl de ADN genómico (como molde en concentración de 10 ng/ µl), 7,5 µl de GoTaq® Green Master Mix (Promega), 1 µl de cada primer (a una concentración de 10 pmoles/ µl cada uno) y 4,5 µl de agua libre de nucleasas. La reacción de termociclaje consistió de un paso de denaturalización de 95°C (2 min), seguido por 30 ciclos de 95°C (30 s), 50°C (aislados 1, 2, 4 y 6) o 65°C (aislados 3, 5 y 7) (30 s), 72°C (4 min), y un paso final de elongación de 72°C (5 min). Las temperaturas de los anillajes se determinaron como las más adecuadas por ensayos previos.
Para verificar la calidad del amplición, los productos de amplificación se corrieron en un gel de agarosa (0,8%) durante 40 min y un voltaje de 65 V. Una vez hecha la electroforesis los geles se visualizaron con ayuda de un transiluminador. Los amplicones obtenidos de cada uno de los siete aislados se secuenciaron en los dos sentidos 5' a 3' y 3' a 5' y las secuencias se ensamblaron utilizando el software Geneious ® v.4,8.4. La secuencia ensamblada se analizó utilizando la herramienta BLAST (Blastn, Megablastn) en varias bases de datos disponibles en Internet (Database 16S ribosomal RNA sequences (Bacteria and Archaea), GenBank; 16S rRNA gene database, Greengenes; Pseudomonas Genome Database; Burkholderia Genome Database) (cuadro 2).
Perfilamiento molecular de las cepas
Con el objetivo de diferenciar las cepas adscritas a una misma clasificación filogenética, se efectuó un perfilamiento molecular a través de la técnica BOX-PCR [16].
Para obtener el ADN genómico de las diferentes cepas se utilizó el protocolo ya descrito. La reacción de amplificación del elemento BOX se hizo con la misma mezcla de reacción usada para la amplificación del gen RNA 16S, excepto que el cebador utilizado fue el BOX A1R (16): 5'-CTACGGCAAGGCGACGCTGACG-3'. Las condiciones y tiempos del termociclaje fueron descritas por Seo and Tsuchiya (17). Los ampliconesBOX se corrieron en un gel de agarosa (1,5%), durante 90 minutos y un voltaje de 75 V. Los patrones electroforéticos se visualizaron en un transiluminador, y se analizaron con ayuda del software TotalLab ® TL120 1D. Para comprobar la robustez y reproducibilidad de la técnica, la corrida de los amplicones BOX se efectuó dos veces, en días distintos, bajo las mismas condiciones experimentales.
RESULTADOS
Aislamiento de BSFy estimación de la eficiencia de solubilización
Se obtuvieron 22 aislamientos bacterianos con capacidad para hacer disponible el P a partir de fosfato tricálcico. Al establecer la estabilidad fenotípica de su capacidad solubilizadora se encontró que, a la quinta resiembra, sólo siete aislados conservaron esta cualidad, los cuales se escogieron para adelantar los ensayos de eficiencia solubilizadora. Estos se denominaron UNBS1 a UNBS7.
Los siete aislamientos presentaron diferencias estadísticas significativas en su valor de ESF y la prueba de Tukey permitió discriminar tres grupos (Cuadro 1, Figura 1). Los aislados UNBS7, UNBS5 y UNBS2 no se diferenciaron estadísticamente y presentaron los mayores valores deESF, y por tanto mayor eficiencia solubilizadora, mientras la cepa UNBS1 fue la de menor halo de solubilización.Los aislamientos UNBS3 y UNBS4no se diferenciaron estadísticamente en su valor ESF del aislado UNBS1.
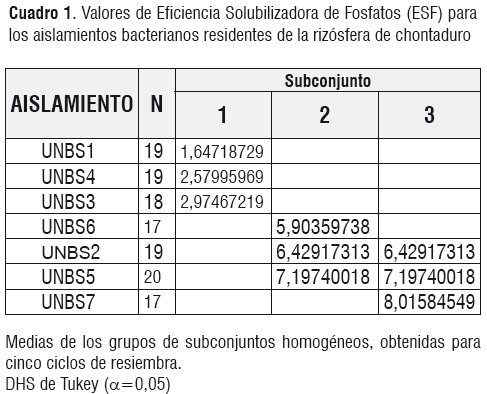
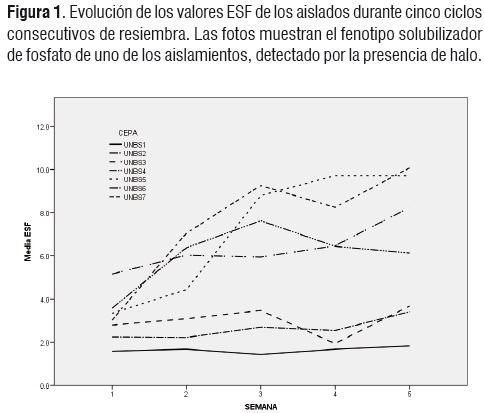
Se encontró que la resiembra sucesiva de los aislados durante cinco semanas llevó a un aumento en el valor de ESF en algunos de ellos (Figura 1), especialmente en UNBS7, UNBS5 y UNBS2.
Análisis de secuencias rDNA 16S de las BSF
El análisis de las secuencias de ADNr 16S amplificadas (Figura 2) demostró muy alta similitudde secuencia entre los aislados UNBS2, UNBS5, UNBS6 y UNBS7 y las secuencias de ADNr 16s de Burkholderia ambifaria MC60-4 (>99%)(Cuadro 2),(Números de accesión CP001025.1, CP001026.1 y CP001027.1 de la base de datos de secuencias del NCBI, cromosomas 1, 2 y 3, secuencias completas), lo que indica que estos aislados se corresponden contal especie, conclusión que se refuerza con los perfiles BOX-PCR (Figura 3), los cuales agrupan los cuatro aislados dentro de un mismo cluster.
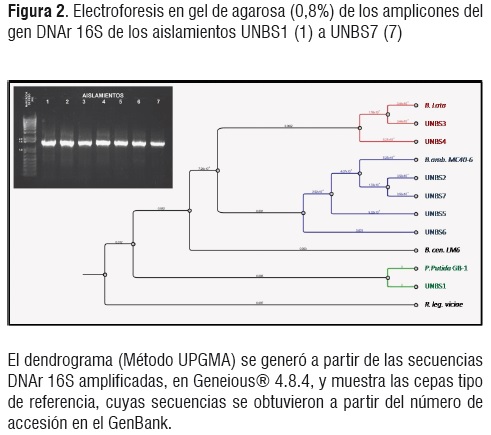
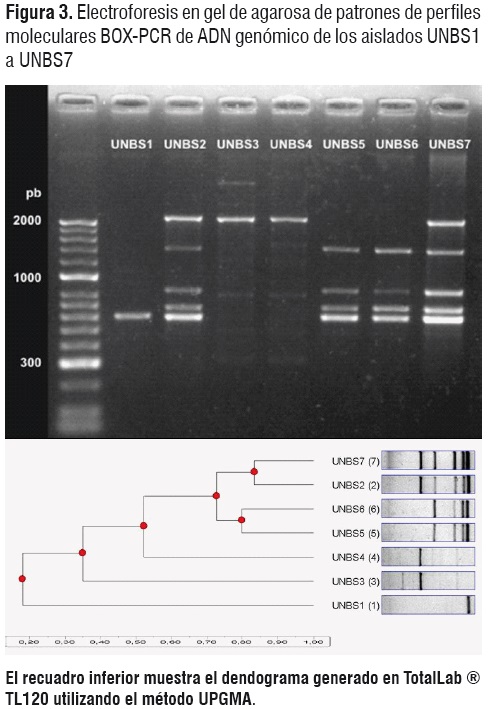
Las secuencias ADNr 16S de los aislamientos UNBS3 y UNBS4 mostraron alta homología a las secuencias correspondientes de Burkholderia sp. 383 (>99%) (Números de accesión CP00152,1, y CP00151,1 de la base de datos de secuencias del NCBI, cromosomas 2 y 1, secuencias completas). Los perfiles BOX-PCR de estos mismos aislados apoyan también su identidad como taxón distinto de las cepas de B. ambifaria MC60-4.Por su parte, el aislamiento UNBS1 mostró mayor homología de secuencia con Pseudomonas putida cepa GB-1 (?96%) (Número de accesión CP000926,1 de la base de datos de secuencias del NCBI, secuencia completa), y el perfil BOX-PCR demostró igualmentemayor nivel de divergencia taxonómica en relación con los demás aislados.
Aunque el análisis de la secuencia ADNr 16S no permitió distinguir como diferentes los aislados UNBS2, UNBS5, UNBS6 y UNBS7, el perfil BOX-PCR (Figura 3) demostró que los aislados UNBS2 y UNBS7 pertenecen a una misma cepa, mientras los aislados UNBS5 y UNBS6 corresponden a otra distinta, dentro de la especie B. ambifaria (Figura 3). Igual situación se presentó para los aislados UNBS3 y UNBS4, los cuales demostraron ser cepas diferentes de la especie Burkholderia sp. 383.
A pesar de que la prueba de siembra en medio PVK permitió obtener 22 aislamientos en una primera oportunidad, 15 (quince) no fueron estables en su capacidad solubilizadora, como se demostró por la desaparición del halo de solubilización en las resiembras consecutivas. Este hecho corrobora la importancia,cuando se seleccionan cepas, de efectuar la confirmación de la estabilidad del fenotipo solubilizador por varios ciclos de siembra consecutivos, que garanticen una base genética que se exprese en el fenotipo.
Al evaluar la ESF de los siete aislados restantes, estables en su capacidad solubilizadora, se encontró un aumento en el índice de eficiencia de la solubilización a través de cinco ciclos de resiembra, lo que indicaría, probablemente,la selección positiva de una característica determinada genéticamente.
Analizados filogenéticamente, los siete aislamientos presentaron alta similaridad con Burkholderia (excepto UNBS1) y cuatro de ellos (UNBS2, UNBS5, UNBS6 y UNBS7) con Burkholderia ambifaria. UNBS1 mostró mayor homología de secuencia con Pseudomonas putida cepa GB-1.
Burkholderia es un género con alta versatilidad metabólica, que agrupa 58 especies reconocidas formalmente (18, 19, 20). En el presente estudio, Burkholderia ambifaria se registra por primera vez en la zona pacífica de Colombia, como habitante de la rizósfera de plantas silvestres de chontaduro, crecidas sin ningún tipo de manejo agronómico. Previamente, Marino et al., [23], aislaron Bacillus sp. y Gordonia sp. como microorganismos BSP, a partir de las raíces de este cultivo, en un suelo de Brasil.
Los diferentes miembros del género Burkholderia registrados como organismos solubilizadores/mineralizadores de P en el suelo y la rizósfera de muchas plantas, deben su capacidad a distintos mecanismos. Caballero-Mellado et al. [21] señalaron a la producción de ácidos orgánicos por parte de varias cepas diazotróficas de Burkholderia, aisladas de la rizósfera y el rizoplano de tomate, como la causade la solubilización del fosforo mineral en ensayos in vitro. En el caso de B. tropica, los mismos investigadores presumen la existencia de un mecanismo desconocido, adicional a la producción de ácidos orgánicos, como responsable de la solubilización. Delvasto et al. [24], demostraron que Burkholderia caribensis FeGL03 produjo ácido glucónico como principal ácido orgánico en medios de cultivo que contenían fosfato tricálcico como única fuente de P, igualmente, Songet al. [25], identificaron ácido glucónico en un cultivo de Burkholderia cepacia, que contenía mineral triturado de hierro, proveniente de una mina, como fuente de P.
Además de su capacidad solubilizadora, el complejo B. cepacia tiene otras características que hacen de sus miembros alternativas promisorias como agentes PGPR. Actúan eficientemente como agentes de biocontrol, tienen alta capacidad degradativa de compuestos xenobióticos, son productores de compuestos reguladores del crecimiento vegetal, particularmente giberelinas y AIA [26, 27] y producen ACC deaminasa, enzima implicada en la hidrólisis del ACC, compuesto que sirve como precursor inmediato de la síntesis del etileno por las plantas [21, 27].
De especial relevancia para aplicaciones agrobiotecnológicas, es el hecho que varias especies de Burkholderia no incluidas [21, 28] y también pertenecientes [22] al complejo B. cepacia, tienen la capacidad de fijar N2. Además, los miembros de Burkholderia compiten ventajosamente por los exudados radicales y de la semilla [22], lo cual les confiere ventajas competitivas en éste nicho, característica clave para que una cepa pueda considerarse un inoculante efectivo.
Al igual que Burkholderia, Pseudomonas ha sido aislado en todas partes del mundo, a partir de diferentes ambientes [29]. Presenta alta gama de capacidades metabólicas, que le hacen que compita con éxito en todos los nichos en que habita. En su interacción con las plantas, Pseudomonas puede tener efectos perjudiciales, pero en la mayoría de los casos, su acción propicia el crecimiento y desarrollo vegetal [30]. Debido a que los exudados radicales y la superficie radical determinan un nicho que favorece las poblaciones altamente competitivas de Pseudomonas, la raíz de las plantas es un reservorio natural de este género [31].
Hay revisiones intensivas sobre los efectos de fitoestimulación, biocontrol y biofertilización, entre otros, que ejercen varias especies de Pseudomonas sobre diferentes cultivos [30,31, 32, 33], así como sus aplicaciones ambientales [34, 35]. Actividades registradas para miembros de las pseudomonadas fluorescentes son la producción de AIA, de ACC deaminasa, de sideróforos y de compuestos con actividad antifúngica, además de la secreción de quitinasas, celulasas y proteasas [36].
En términos de capacidad solubilizadora de fosfatos, Pseudomonas es uno de los géneros bacterianos más eficientes. Naiket al. [36], en un extenso estudio sobre la capacidad solubilizadora de fosfatos de 443 aislamientos de pseudomonadas fluorescentes, obtenidos a partir de la rizosfera de banano y arroz, encontraron que 80 fueron eficientes solubilizadores de P, incluyendo: Pseudomonas monteilii, P. putida, P. plecoglossicida, P. fluorescens, P. fulva, P. mosselii y P. aeruginosa. Dentro de éstas, 12 (15%) se clasificaron como P. putida. El análisis de BOX-PCR determinó además, amplia diversidad intraespecífica entre los aislamientos. En un estudio similar, Naiket al. [37] no encontraron a P. putida en la rizosfera de banano, lo cual asociaron a posibles variaciones condicionadas por el genotipo del huésped. Browneet al. [38], encontraron mayor densidad de pseudomonadas fluorescentes en trigo en comparación con cebada, aunque las poblaciones de BSP no variaron significativamente entre los dos cultivos. En ambas plantas, la mayor tasa de solubilización estuvo asociada a aislados de la especie P. fluorescens.
Con relación a los posibles mecanismos de solubilización de fosfatos, varios estudios confirman que en Pseudomonas, incluyendo P. putida, están asociados a la producción de ácidos orgánicos, especialmente a través de la oxidación de glucosa a ácido glucónicopor la ruta de oxidación directa, la que estáa su vez relacionada directamente con la acidificación del medio de cultivo [36].
CONCLUSIONES
A partir de rizosfera de chontaduro crecido naturalmente en la Costa Pacífica del Valle del Cauca, se obtuvieron 22 aislamientos bacterianos con capacidad temporal de solubilización de P (BSP). Siete mostraron estabilidad en la característica solubilizadora.
El género bacteriano con mayor frecuencia dentro de las BSP aisladas correspondió a Burkholderia, incluyendo 3 cepas de B. ambifaria y dos de Burkholderia sp. 383, registradas en la literatura como eficientes PGPR, con actividades benéficas relacionadas con: control de fitopatógenos, producción de sustancias con acción hormonal, antibiótica, diazotrófica y enzimática. También se aisló una cepa de Pseudomonas putida.
El perfil molecular BOX-PCR de las cepas evidenció la ocurrencia de diversidad intraespecífica entre las cepas de B. ambifaria, indetectable con el análisis de las secuencias del ADNr 16S. Se considera necesario el uso complementario de las dos técnicas en análisis de biodiversidad microbiana asociada a la rizosfera de los cultivos.
AGRADECIMIENTOS
Los autores manifiestan su agradecimiento a la Dra. Karina López y al profesor Jaime Muñoz, por su asesoría y por facilitar las instalaciones y recursos de los laboratorios de Microbiología y Biología Molecular, respectivamente, en la Universidad Nacional de Colombia, sede Palmira.
Agradecemos igualmente a la Vicerrectoría de Investigaciones de la misma universidad, por el apoyo financiero logrado por el proyecto como uno de los ganadores de la Convocatoria Nacional de Investigación, del año 2009.
REFERENCIAS
[1] Yarzabal, L. A. En: Soil biology and agriculture in the Tropics. Soil Biology, vol. 21. Berlin (Germany): Springer-Verlag, 2010, p. 209 - 233. [ Links ]
[2] Fageria, N. K.; Baligar, V. C. Ameliorating soil acidity of tropical Oxisols by liming for sustainable crop production. Adv. Agron., 99, 2008, p. 345-399. [ Links ]
[3] Malagón, D. Suelos de Colombia. Santafé de Bogotá D.C. (Colombia): Instituto Geográfico Agustín Codazzi, 1995, 632 p. [ Links ]
[4] Cordell, D.; Drangert, J. A. and White, S. The story of phosphorus: Global food security and food for thought. Global Environment Change, 19, 2009: p. 292 -305. [ Links ]
[5] Vaccari, D. A. Phosphorus: A looming. Scientific American, June, 2009: p. 55 – 59. [ Links ]
[6] Marschner, P. Marschner's Mineral Nutrition of Higher Plants, Third Edition. Academic Press, 2011, 672 p. [ Links ]
[7] Marschner, P. En: Nutrient cycling in terrestrial ecosystems. Berlin (Germany): Springer-Verlag, 2007, p. 159 - 181. [ Links ]
[8] Bünemann, E. K.; Condron, L. M. En: Nutrient cycling in terrestrial ecosystems. Berlin (Germany): Springer-Verlag, 2007, p. 65-92. [ Links ]
[9] Rodriguez, H. and Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol Adv., 17, 1999: p. 319-339. [ Links ]
[10] Hamdali, H.; Hafidi, M.; Virolle, M.J.; Ouhdouch, Y. Rock phosphate-solubilizing Actinomycetes: screening for plant growth promoting activities. World J Microbiol Biotechnol., 24, 2008: p. 2565–2575. [ Links ]
[11] Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi – current perspective. Arch Agron Soil Scien. 56(1), 2010: 73–98. [ Links ]
[12] Zaidi, A.; Khan, M. S.; Ahemad, M.; Oves, M.; Wani, P. A. En: Microbial strategies for crop improvement. Berlin (Germany): Springer-Verlag, 2009: p. 23-50. [ Links ]
[13] Uribe, D., Sánchez-Nieves, J. et al.En: Soil Biology and Agriculture in the Tropics. Berlin (Germany): Springer-Verlag, 2010:235-250. [ Links ]
[14] Moreno Sarmiento, N.; Uribe Vélez, D. y Moreno L. En: Biofertilizantes en Iberoamérica: visión técnica, científica y empresarial. En: Uruguay, BIOFAG, 2007: p. 38 – 45. [ Links ]
[15] Weisburg, W. G., Barns, S. M., Pelletier, D. A. and Lane, D. J. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol., 173, 1991: p. 697-703. [ Links ]
[16] Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based Polymerase Chain Reaction. Methods Mol Cell Biol. 5, 1994: 25-40. [ Links ]
[17] Seo, S. T. and Tsuchiya, K. Genotypic characterization of Burkholderiacenocepacia strains by rep-PCR and PCR–RFLP of the fliC gene. FEMS Microbiology Letters, 245, 2005: p. 19–24. [ Links ]
[18] Wong-Villarreal, A.; Caballero-Mellado, J. Rapid identification of nitrogen-fixing and legume-nodulating Burkholderia species based on PCR 16S rRNA species-specific oligonucleotides. SystApplMicrobiol. 33, 2010: p. 35–43. [ Links ]
[19] Compant, S.; Nowak, J.; Coenye, T.; Clément, C.; Barka, E. A. Diversity and occurrence of Burkholderiaspp. in the natural environment. FEMS Microbiol Rev. 32, 2008: p. 607-626. [ Links ]
[20] Chiarini, L.; Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Visca, P. Burkholderiacepacia complex species: health hazards and biotechnological potential. Trends Microbiol. 14(6), 2006: p. 277-286. [ Links ]
[21] Caballero-Mellado, J.; Onofre L., J.; Estrada de los Santos, P.; Martínez Aguilar, L. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Appl Environ Microbiol. 73(16), 2007: p. 5308-5319. [ Links ]
[22] Tabacchioni, S.; Bevivino, A.; Dalmastri, C.; Chiarini, L. Burkholderiacepacia complex in the rhizosphere: a minireview. Annals Microbiol. 52, 2002: p. 103-117. [ Links ]
[23] Marino, W.; Marschner, P.; Gasparotto, L. Phosphate Solubilizing Bacteria from the Rhizosphere of Theobroma grandiflorum (Willd. ex Spreng.) Schum. and Bactris gasipaes H.B.K.: Potential for Plant Growth Promotion. German-Brazilian Workshop on Neotropical Ecosystems – Achievements and Prospects of Cooperative Research, Hamburg, September 3-8, 2000. Recuperation of Degraded Areas – Summaries of Lectures, Session 5. [ Links ]
[24] Delvasto, P.; Ballester, A.; Muñoz, J.A.; González, F.; Blásquez, M. L.; Igual, J. M.; Valverde, A. Mobilization of phosphorus from iron ore by the bacterium Burkholderiacaribensis FeGLO3. Miner Eng. 22, 2009: p. 1-9. [ Links ]
[25] Song, O. K.; Lee, S. J.; Lee, Y. S.; Lee, S. C.; Kim, K. K.; Choi, Y. L. Solubilization of insoluble inorganic phosphate by Burkholderia cepacia DA23 isolated from cultivated soil. Braz J of Microbiol. 39, 2008: p. 151-156. [ Links ]
[26] Ludovic, V.; Groleau, M. C.; Dekimpe, V.; Deziel, E. Burkholderia diversity and versatility: an inventory of the extracellular products. J Microbiol Biotechnol. 17(9), 2007: p. 1407-1429. [ Links ]
[27] Sun, Y.; Cheng, Z.; Glick, B. R. The presence of a 1-aminocyclopropane-1-carboxylate (ACC) deaminase deletion mutation alters the physiology of the endophytic plant growth-promoting bacterium Burkholderia phytofirmans Ps JN. FEMS MicrobiolLett. 296, 2009: p. 131-136. [ Links ]
[28] Sprent, J. I. 60Ma of legume nodulation. What's new? What's changing? J Exp Bot. 59(5), 2008: p. 1081-1084. [ Links ]
[29] Peix, A.; Ramírez Bahena, M.; Velásquez, E. Historical evolution and current status of the taxonomy of genus Pseudomonas. Infec Genet Evol. 9, 2009: p. 1132–1147. [ Links ]
[30] Miller, S. H.; Mark, G. L.; Franks, A.; O'gara, F. En: Pseudomonas. Model Organism, Pathogen, Cell Factory. Wiley – VCH Verlag GmbH & Co. KGaA, Weinheim, 2008: p. 353 – 376 pp. [ Links ]
[31] Mercado Blanco, J.; Bakker, P. A. H. Interactions between plants and beneficial Pseudomonas spp.: exploiting bacterial traits for crop protection. Antonie van Leeuwenhoek. 92, 2007: p. 367–389. [ Links ]
[32] Höfte, M.; Altier, N. Fluorescent pseudomonads as biocontrol agents for sustainable agricultural systems, Res Microbiol. 2010. doi: 10.1016/j.resmic.2010.04.007. [ Links ]
[33] Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat Rev Microbiol. 3, 2005: p.307-319. [ Links ]
[34] Rehm, B. H. A. En: Pseudomonas. Model Organism, Pathogen, Cell Factory. Wiley – VCH Verlag GmbH & Co. KGaA, Weinheim., 2008: p. 377 – 395. [ Links ]
[35] Jiménez, J. I.; Nogales, J.; García, J. L.; Díaz, E. En: Handbook of Hydrocarbon and Lipid Microbiology, Springer-Verlag, Berlin, 2010: p. 1297-1325. [ Links ]
[36] Naik, P. R.; Raman, G.; Narayanan, K. B.; Sakthivel, N. Assessment of genetic and functional diversity of phosphate solubilizing fluorescent pseudomonads isolated from rhizospheric soil. BMC Microbiol.8, 2008: p. 230. [ Links ]
[37] Naik, P. R.; Sahoo, N.; Goswami, D.; Ayyadurai, N.; Sakthivel, N. Genetic and functional diversity among fluorescent pseudomonads isolated from the rhizosphere of banana. Microb Ecol. 56, 2008: 492–504. [ Links ]
[38] Browne, P.; Rice, O.; Miller, S. H.; Burke, J.; Dowling, D. N.; Morrissey, J. P.; O'gara, F. Superior inorganic phosphate solubilization is linked to phylogeny within the Pseudomonas fluorescens complex. Appl Soil Ecol. 43(1), 2009: p. 131-138. [ Links ]
