










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBiotecnología en el Sector Agropecuario y Agroindustrial
Print version ISSN 1692-3561
Rev.Bio.Agro vol.15 no.1 Popayán Jan./June 2017
https://doi.org/10.18684/BSAA(15)85-94
DOI: http://dx.doi.org/10.18684/BSAA(15)85-94
DIVERSIDAD GENÉTICA EN PATRONES DE CÍTRICOS MEDIANTE MICROSATÉLITES AMPLIFICADOS AL AZAR (RAMs)
GENETIC DIVERSITY ON CITRUS ROOTSTOCKS WITH RANDOM AMPLIFIED MICROSATELLITES (RAMs)
DEVERSIDADE GENETICA DE PADROES DE CITROS USANDO MICROSSATELITES AMPLIFICADOS AO ACASO (RAMs)
JEISSON STEEVENS GALLEGO-COLONIA1, AYDA LILIA ENRÍQUEZ-VALENCIA1, ÁLVARO CAICEDO-ARANA2, ANDRÉS MAURICIO POSSO-TERRANOVA3, JAIME EDUARDO MUÑOZ-FLOREZ4.
1 Universidad Nacional de Colombia Sede Palmira, Facultad de Ciencias Agropecuarias. Ingeniera Agrónoma. Palmira, Colombia.
2 Corporación Colombiana de Investigación Agropecuaria (CORPOICA). Ingeniero Agrónomo. Palmira, Colombia.
3 Universidad Nacional de Colombia Sede Palmira, Facultad de Ciencias Agropecuarias. Ph.d. en Biólogia. Palmira, Colombia.
4 Universidad Nacional de Colombia Sede Palmira, Facultad de Ciencias Agropecuarias. Ph.d. Ciencias Agrarias. Palmira, Colombia
Correspondencia: alenriquez.v@hotmail.com.
Recibido para evaluación: 17 de Julio de 2015. Aprobado para publicación: 25 de Julio de 2016.
RESUMEN
Los patrones de cítricos son esenciales en la citricultura debido a que afectan más de veinte características agronómicas de las copas y frutos. La caracterización y documentación de las introducciones de un banco de germoplasma permiten evitar duplicados e identificar introducciones promisorias para procesos de selección, mejoramiento genético o procesos agroindustriales. Se evaluó la diversidad genética de 39 introducciones del banco de germoplasma de Cítricos de CORPOICA Centro de Investigación Palmira. Se utilizaron seis cebadores RAMs los cuales generaron 134 loci polimórficos que se usaron para estimar los parámetros de diversidad genética. La He y porcentaje de loci polimórficos para la colección fue de 0,38 y 92,54%, respectivamente. El análisis de similitud permite apreciar que los materiales se encuentran dispersos en el dendrograma, sin embargo a un nivel de similitud de 0,63 se agrupan las introducciones del género Poncirus y sus híbridos. La especie Clausena lansium fue la más distante del resto de las especies y se diferenció a un nivel de similitud de 0,54. Los resultados indican alta diversidad genética en el banco de germoplasma de patrones de cítricos, que puede ser utilizada en futuros trabajos de investigación.
PALABRAS CLAVE: Citrus, Biodiversidad, Caracterización, Germoplasma, Molecular.
ABSTRACT
Citrus rootstocks are essential in the citrus industry because they affect more than twenty agronomic characteristics of the citrus and fruits. Characterization and documentation of germplasm bank introductions allow to avoid duplicates and to identify promising accessions for selection processes, breeding or agro-industrial processes. It was evaluated the genetic diversity of 39 accessions from the citrus rootstocks germplasm bank of the Colombian Corporation for Agricultural Research (Corpoica-Palmira). Six RAMs primers were used which generated 134 polymorphic loci that were used to estimate genetic diversity parameters. Expected heterogeneity and polymorphic loci percentage for the collection was 0,38 and 92,54%, respectively. Similarity analysis allow to appreciate that the materials are dispersed in the dendrogram, however a similarity level of 0,63 grouped the accessions of the genus Poncirus and their hybrids. The species Clausena lansium was the most distant from other species in the dendrogram and differed at a similarity level of 0,54. The results indicate a high genetic diversity in the citrus rootstocks germplasm bank of Corpoica that can be used in future research programs.
KEYWORDS: Banks, Biodiversity, Characterization, Germplasm, Molecular.
RESUMO
Os porta-enxertos de cítricos são essenciais na cultura de citros porque afetam mais de vinte características agronômicas das cultivares-copa e dos frutos. A caracterização e documentação das introduções de um banco de germoplasma impedem duplicações e permitem identificar acessões promissoras para processos de seleção, melhoramento genético ou processos agroindustriais. Foi avaliada a diversidade genética de 39 acessões do banco de germoplasma de Citrus da Corporación Colombiana de Pesquisa Agropecuária (CORPOICA), centro de pesquisa Palmira. Foram usados seis primers RAMs, os quais geraram 134 loci polimórficos que foram utilizados para estimar os parâmetros de diversidade genética. A heterogeneidade esperada e a percentagem de loci polimórficos para a coleção foi de 0,38 e 92,54%, respectivamente. A análise de similaridade permitiu apreciar que os materiais estão dispersos no dendrograma, no entanto, num nível de similaridade de 0,63 estão agrupadas as acessões do gênero Poncirus e seus híbridos. A espécie Clausena Lansium foi o mais distante das outras espécies no dendrograma e diferiu num nível de similaridade de 0,54. Os resultados mostram alta diversidade genética no banco de germoplasma de padrões de citros de CORPOICA C.I. Palmira, que pode ser utilizada em futuros trabalhos de pesquisa.
PALAVRAS-CHAVE: Bancos, Biodiversidade, Caracterização, Germoplasma, Molecular.
INTRODUCCIÓN
Los cítricos cultivados actualmente se caracterizan por estar formados por la unión de dos individuos por medio del injerto. El patrón constituye el sistema radical y una parte del tronco, sobre el cual se injerta la yema, vareta o púa que forma la copa o parte aérea la cual produce los frutos de interés comercial. El empleo del patrón no sólo ha logrado mejorar los rendimientos y la calidad de los frutos, sino que también ha permitido la siembra en sitios con características adversas para el normal desarrollo de la planta [1].
Los cítricos pertenecen a la familia Rutaceae, formada por siete subfamilias, una de ellas es Aurantioideae, la cual tiene dos tribus, Clauseneae y Citreae. Clauseneae consiste de tres subtribus, entre ellas Clauseninae donde se destaca el género Clausena [2]. Las especies del genero Poncirus hibridan libremente con las especies de cítricos generando grupos de híbridos, que son citranges (Poncirus trifoliata x Citrus sinensis), citrandarinas (Poncirus trifoliata x Citrus reticulata), citremones (Poncirus trifoliata x Citrus limonia) y citrumelos (Poncirus trifoliata x Citrus paradisi), que también reconoce a los tangores como el cruce interespecífico entre mandarina (Citrus reticulata) y naranja (Citrus sinensis) [3].
En los bancos de germoplasma vegetal, la caracterización y documentación de las introducciones permiten evitar duplicados e identificar introducciones promisorias para procesos de selección, mejoramiento genético o procesos agroindustriales [3]. En el banco de germoplasma de cítricos de CORPOICA C.I. (Palmira), se encuentran 40 introducciones de patrones de Citrus y géneros relacionados, establecidos en materas tipo bonsái, muchos de origen desconocido, razón por la cual es necesario hacer estudios moleculares que evalúen la diversidad genética de este banco de germoplasma.
El uso de marcadores moleculares ha sido un instrumento valioso y preciso para la evaluación de la diversidad genética en cítricos y géneros relacionados, incluyendo marcadores isoenzimaticos [4], SRAP [5], RAPD [6], AFLP [7], En Colombia, se evaluó la diversidad genética del banco de germoplasma de cítricos en naranjas Citrus sinensis [8, 9], sin embargo en la colección de patrones de cítricos aún es desconocido. Hasta la fecha los reportes acerca de la medición de la diversidad genética de cítricos y géneros relacionados usados como patrones son escasos.
Los marcadores moleculares RAMs son útiles para medir la diversidad genética en plantas y animales, diferencia entre familias, entre especies y al interior de la especie [10]. El objetivo de este estudio fue determinar el nivel y organización de la diversidad genética de patrones de cítricos del banco de germoplasma de la Corporación Colombiana de Investigación Agropecuaria CORPOICA C.I. Palmira, usando marcadores moleculares RAMs.
MÉTODO
Material vegetal
Se colectó hojas jóvenes de 39 accesiones de patrones de cítricos procedentes del banco de germoplasma de CORPOICA C.I. Palmira (cuadro 1), localizada en Palmira, Valle del Cauca, Colombia, a 1001 msnm, con una precipitación anual de 1.174 mm y temperatura promedio de 25°C. La colección de estudio cuenta con materiales pertenecientes a tres géneros de la familia Rutaceae: Citrus, Poncirus y Clausena.
Extracción de ADN y amplificación con cebadores RAMs
El tejido vegetal se maceró en nitrógeno líquido y se realizó la extracción de ADN siguiendo la metodología [11] con algunas modificaciones. Para la evaluación de la calidad y cantidad del ADN obtenido se realizaron geles de agarosa al 0,8% corridos en buffer TBE 0,5X (Tris-borato 0045 M; EDTA (ácido etildiaminotetraacético 0,001M) a 100 voltios durante 50 minutos, teñidos con bromuro de etidio a una concentración final de 0,5 µg/mL, y visualizados bajo luz ultravioleta.
Las concentraciones se determinaron por comparación con concentraciones conocidas de ADN del bacteriófago lambda. El ADN cuantificado se diluyó en agua HPLC hasta tener la concentración final de 10 ng/µL. El ADN de las accesiones se almacenó en cajas criogénicas a -20°C, con sus respectivas identificaciones, en el Laboratorio de Biología Molecular de la Universidad Nacional de Colombia, Sede Palmira.
Para la caracterización molecular se utilizaron siete cebadores RAMs, teniendo en cuenta su habilidad para revelar polimorfismo, el número de bandas polimórficas y su reproducibilidad (cuadro 2). La concentración de la reacción fue: Tampón Taq 1X, dNTPs 0,2 mM (para cada uno), cebadores 4 µM, MgCl2 2,5 mM, una unidad de Taq polimerasa y 10 ng de ADN en un volumen final de 25 µL. La amplificación se realizó en un termociclador (PTC-100 Programable Termal Controller), con las siguientes condiciones: 95°C por 5 minutos, seguido de 37 ciclos de 95°C por 30 segundos, 50-58°C por 45 segundos (de acuerdo al cebador utilizado (cuadro 2), 72°C por 2 minutos y una extensión final de 72°C por 7 minutos. Los productos amplificados se separaron por electroforesis en geles de poliacrilamida al 6% (37:1 acrilamida - bisacrilamida) en una cámara Biometra, corridos a 150 voltios y 400 mA, durante una hora y 10 minutos. La tinción se realizó utilizando sales de plata (figura 1) [12].
 Cuadro 2
Cuadro 2
Análisis de datos
La lectura de las bandas se realizó considerando como locus polimórfico aquel en el cual el alelo más común tiene una frecuencia menor al 95% [13]. Se generó una matriz binaria de presencia (uno) y ausencia (cero). A partir de esta matriz y usando los programas SIMQUAL del paquete NTSYS-pc (Numerical Taxonomy System for Personal Computer, versión 2.02 PC) y el programa TFPGA (Tools for Population Genetic Analisys) se realizaron los análisis estadísticos.
Para cada cebador se obtuvo el número de bandas totales y bandas polimórficas. La similitud genética se calculó mediante el coeficiente de Dice Nei-Li [14]: S=(2a/2a+b+c), donde a = bandas compartidas por ambos individuos, b = bandas presentes en el individuo (1) pero no en (2), y c = bandas presentes en el individuo (2) pero no en (1). El análisis de agrupamiento se realizó con el programa SAHN de NTSYS-pc (versión 2) utilizando el método UPGMA, método gráfico de agrupamiento por parejas, que usa el promedio aritmético no ponderado. El dendrograma se construyó con el programa TREE de NTSYS-pc. Para estimar la diversidad genética se utilizaron los parámetros de heterocigosidad promedio esperada (He) y el porcentaje de loci polimórficos (P), los cuales se estimaron sobre todos los loci y el promedio de los mismos de acuerdo con la fórmula no sesgada de [15].
RESULTADOS
Marcadores moleculares RAM's
Se seleccionaron seis cebadores RAM's de los siete inicialmente considerados para el estudio. El cebador CGA se descartó del análisis debido a que no amplificó para la mayoría de las muestras. Los seis cebadores generaron 134 bandas que oscilaron entre 17 para el cebador GT y 26 para el cebador CA (cuadro 3), lo cual también coincide con el número de loci polimórficos, con excepción del cebador CCA que género 18 loci polimórficos de un total de 20 loci.
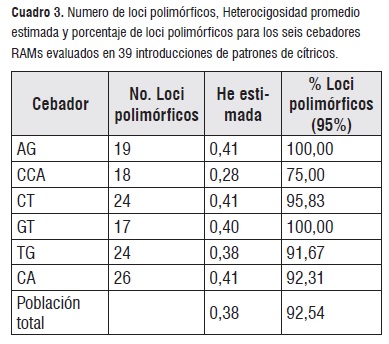 Cuadro 3
Cuadro 3Las condiciones de amplificación para los cebadores RAM's se tomaron en cuenta de acuerdo a lo descrito por [10]. Sin embargo, las condiciones de amplificación pueden variar dependiendo de la especie estudiada.
En este estudio se hizo una variación en la temperatura de hibridación del primer AG (48°C) (figura 1).
Análisis de Diversidad Genética
La heterocigosidad esperada (He) para las 39 introducciones de patrones de Cítricos fue de 0,38, indicando un alto nivel de diversidad genética en la colección (cuadro 3). Teniendo en cuenta que 0 = He = 0,3 [16], este valor puede estar asociado a que se evaluaron tres géneros diferentes, Citrus, Poncirus y Clausena, y al interior de ellos diferentes especies y cruzamientos (híbridos). En la caracterización molecular con RAM's del grupo de naranjas del banco de germoplasma de cítricos de CORPOICA, se encontró un valor de He de 0,31 [8], el cual también es alto y similar a lo encontrado en este estudio para patrones de cítricos.
El porcentaje de loci polimórfico encontrado para la colección fue de 92,54%, indica alta diversidad genética, lo cual se debe posiblemente a que la colección está formada por introducciones de tres géneros diferentes y además la mayoría de estos materiales son híbridos intergenéricos e interespecíficos. Los valores de He y porcentaje de loci polimórficos para cada uno de los cebadores RAM's fueron altos. Los valores de He variaron entre 0,41 para AG, y 0,28 para CCA, y valores intermedios importantes fueron registrados en CT y CA: 0,41, GT: 0,40, TG: 0,38. El porcentaje de loci polimórficos osciló entre 75% para CCA, y 100% para AG y GT; de igual forma se registraron valores importantes en los cebadores CT: 95,83%, CA: 92,31%, TG: 91,67%. Los cebadores con secuencias repetidas de dos pares de bases fueron más polimórficos que CCA con tres pares de bases. De manera general hubo correlación entre He y polimorfismo en los cebadores evaluados. Los resultados indican que los seis cebadores evaluados realizaron un aporte significativo al estudio de la diversidad genética de patrones de cítricos constituyéndose en marcadores moleculares útiles para futuros trabajos de investigación.
La colección está compuesta por especies e híbridos de tres géneros diferentes, por lo tanto ésta se organizó en tres grupos: el grupo uno corresponde a materiales del género Poncirus y sus híbridos con el género Citrus; el grupo dos compuesto por introducciones del género Citrus e híbridos interespecíficos (tangores); y el grupo tres formado por un material del género Clausena.
Debe aclararse que este último no es un grupo como tal debido a que está formado por un solo material y por lo anterior no registró valores de heterocigosidad y porcentaje de loci polimórficos. Estos valores para el grupo uno fueron de 0,34 y 82,09%, respectivamente; mientras que para la población dos, fueron de 0,37 y 90,30%, respectivamente. Estos resultados indican que el grupo dos tiene la mayor diversidad genética de la colección. Lo anterior puede ser explicado debido a que este grupo se compone de mandarinas, naranjas, limones, pomelos, híbridos, y otros materiales que no están incluidos dentro de los grupos anteriores, por ejemplo Citrus lycopersicae.
En mandarina Citrus reticulata Blanco, se encontró alta heterogeneidad genética (86% polimorfismo) utilizando AFLP a pesar de considerar introducciones altamente relacionadas [17]. Utilizando otras técnicas moleculares, isoenzimas [18], RFLP, RAPD [19] y microsatélites [20] se ha concluido que este grupo reporta mayor diversidad genética dentro del género Citrus.
En naranja Citrus sinensis L. Osbeck, se desarrollaron y caracterizaron marcadores moleculares microsatélites polimórficos, encontraron valores de He promedio de 0,50, y de heterocigosidad observada (Ho) promedio de 0,99 [21], que son valores altos que justifican la diversidad genética del género Citrus. Sin embargo, en otros estudios con RAM's y microsatélites, se encontraron valores de He de 0,22 y 0,25, respectivamente, y porcentaje de loci polimórficos de 49% y 64,8%, respectivamente [8, 9], los cuales son bajos y contrastan con el presente estudio debido a que las introducciones evaluadas son cultivares clonados de naranjas no híbridas, mientras que en este estudio de la diversidad genética de patrones de cítricos la colección cuenta con materiales híbridos (dos tangores y siete citranges) y una accesión de naranja no híbrida (Salerma).
Los pomelos Citrus paradisi, se caracterizan por presentan baja diversidad entre ellos, sin embargo, al ser comparados con otras especies del género Citrus, como es el caso del presente estudio, se obtiene alta diversidad. Se evaluaron la variabilidad genética de 38 introducciones de Citrus paradisi usando RAPD y SSR encontrando bajo polimorfismo entre los materiales (49%) [22]. Esto sugiere que el polimorfismo morfológico observado entre los pomelos debe estar asociado con mutaciones somáticas, las cuales no son detectadas por los marcadores moleculares utilizados.
A pesar que la mayor diversidad se encontró en el grupo de Citrus (He 0,37 y 90,30% loci polimórficos) se debe tener en cuenta que la diversidad del grupo uno (Poncirus y sus híbridos con Citrus) también es alta (He 0,34 y 82,09% loci polimórficos). Lo anterior puede atribuirse a que éste grupo incluye citranges, citremones, citrumelos, citrandarinas, que son materiales provenientes del cruce entre dos géneros (Cuadro 1).
Distancia Genética
Los valores de distancia genética calculados con el índice de Nei [14] mostraron que los grupos 1 y 2, Poncirus y Citrus, respectivamente, son más cercanos entre ellos (0,05), al ser comparadas con Clausena lansium (0,28) (figura 2). Lo anterior puede ser explicado debido a que los géneros Poncirus y Citrus pertenecen a la misma subtribu (Citrinae), dentro de la tribu Citreae, mientras que el género Clausena se incluye en la subtribu Clauseninae, dentro de la tribu Clauseneae, ubicándose lejos taxonómicamente.
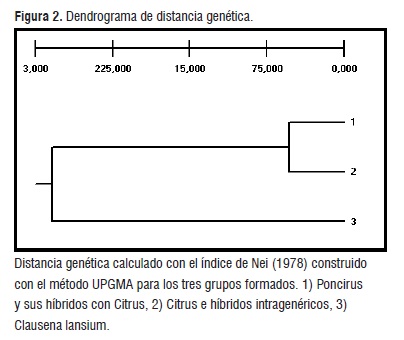 Figura 2
Figura 2Análisis de Similitud Genética
El análisis mediante el coeficiente de Dice Nei-Li permite apreciar que los materiales del banco de germoplasma se encuentran dispersos en el dendrograma (figura 3). La falta de agrupamientos puede deberse a que en este estudio no se evaluó una sola especie, sino varias especies e híbridos pertenecientes a tres géneros diferentes. Además, este banco, está compuesto por introducciones de diferentes partes del mundo, en particular de Riverside - California, Florida (E.U), Valencia (España), Embrapa (Brasil), entre otros centros de investigación, universidades y fincas de agricultores lo cual hace que la diversidad de la colección sea más amplia.
Sin embargo, a un nivel de similitud de 0,63 se agrupan todos los materiales trifoliados, los cuales corresponden a Poncirus y sus híbridos. La influencia de Poncirus trifoliata es fuerte en los citranges evidenciado por la naturaleza trifoliada de sus hojas, así como la acidez y amargura de sus frutos y la resistencia al frio [23]. Este nivel de similitud es consistente con el estudio de (5) donde utilizaron marcadores SRAP con el fin de detectar polimorfismo entre 86 materiales de Citrus y especies cercanas en la subfamilia Aurantioidea el grupo de Poncirus y sus híbridos se agrupó a un nivel de similitud de 0,55. 'Pomelo Africa' (Citrus paradisi) fue la única accesión del género Citrus que se agrupó con los materiales trifoliados. Lo anterior puede ser explicado debido a que 'Pomelo Africa' puede estar compartiendo características en común con los materiales de este grupo, el cual posee híbridos de Citrus paradisi con Poncirus trifoliata.
Dentro del grupo de los materiales trifoliados, a un nivel de similitud del 0,66 se diferencia un subgrupo el cual lo conforman tres materiales Poncirus trifoliata ('English large', 'Kryder 15-3' y 'Pomeroy'). Otro subgrupo al mismo nivel de similitud reúne materiales Poncirus trifoliata e híbridos. Dentro de este subgrupo están dos citremones 1449 (Poncirus trifoliata x Citrus limon), los cuales inicialmente se pensaba que se trataba del mismo material en el banco de germoplasma, pero la técnica RAM's permitió la diferenciación de estos materiales. Estos citremones tienen el nivel de similitud más alto del análisis (0,92).
Los citranges 'Troyer' y 'Carrizo' (Citrus sinensis x Poncirus trifoliata), importantes patrones en la citricultura, se agrupan a un nivel de similitud de 0,83, lo que demuestra que estos materiales son cercanos como lo revela la técnica RAMs en este estudio. Con marcadores SRAP, se encontró que 'Troyer' y 'Carrizo' fueron casi idénticos (0,99) [5]. 'Troyer' y 'Carrizo' son en realidad un clon único el cual se originó como una plántula zigótica (CPB 4-5019) proveniente del cruce de naranja 'Washington navel' Citrus sinensis y 'naranja trifoliada' Poncirus trifoliata, en lugar de las dos plantas hermanas como se había supuesto [24].
Aunque morfológicamente son casi idénticos, el citrange 'Carrizo' tiene ventajas agronómicas que han desplazado el uso del citrange 'Troyer' [25].
Las citrandarinas 'Sunky x Jacobson' y 'Sunky x English' (Citrus sunky x Poncirus trifoliata), se agruparon a un nivel de similitud de 0,84. Este resultado se esperaba, considerando que ambas introducciones tienen un parental en común (Citrus sunky) y dos líneas hermanas: Jacobson e English (Poncirus trifoliata).
Fuera de este gran grupo de materiales trifoliados (Poncirus y sus híbridos), los demás materiales pertenecientes a Citrus forman agrupaciones interesantes. Las limas y limones no forman una agrupación clara, no obstante hay algunas tendencias. La lima 'Rangpur' y el limón 'Milan' tienden a agruparse a un nivel de similitud de 0,62 con el grupo que reúne mandarina, naranja, pomelo, e híbridos, descrito anteriormente.
La lima 'Rangpur', es bastante diferente morfológicamente y genotípicamente de las limas y fue considerada como Citrus reticulata [21]. El origen y los parentales de los rangpurs son inciertos, pero generalmente han sido clasificados con las mandarinas en estudios previos [5]. 'Rangpur' es un híbrido de citrón y mandarina (Citrus medica x Citrus reticulata) [26].
Naranjo agrio Citrus aurantium, fue distante de los demás materiales (0,57), lo cual puede explicarse dado que es una de las especies más primitivas de los cítricos reportados en la historia después del cidro (Citrus medica). Su proceso de evolución y domesticación no ha estado tan marcado como el naranjo dulce y la naranja trifoliada [7].
Clausena lansium, única especie representativa del género Clausena, fue la especie más distante del resto de las especies en el dendrograma, con un nivel de similitud de 0,54. Esto puede ser debido a que Clausena lansium pertenece a la tribu Clauseneae, una de las dos tribus de la subfamilia Aurantioideae, la cual también incluye la tribu Citreae. La tribu Citreae incluye los géneros Citrus y Poncirus [2]. Con marcadores SRAP, separaron a un nivel de similitud de 0,32 a Clausena lansium de los géneros de la subtribu Citrineae, entre ellos, Poncirus y Citrus [5], y por lo tanto, los resultados obtenidos están de acuerdo a lo reportado anteriormente.
CONCLUSIONES
Los marcadores moleculares RAMs son útiles para la evaluación de la diversidad genética en Citrus y géneros relacionados, debido a que mostraron altos niveles de polimorfismo, además, pudieron diferenciar el género Citrus de sus parientes distantes. La Técnica RAMs fue eficiente en la discriminación de los tres géneros que componen a los patrones de cítricos del banco de germoplasma CORPOICA (Citrus, Poncirus y Clausena). La distancia genética entre Citrus y Poncirus fue menor en comparación con el género Clausena, debido a que se incluyen taxonómicamente en tribus diferentes.
Se esperaba que el grupo uno (Poncirus y sus híbridos) tuviese la mayor diversidad genética porque incluye materiales provenientes del cruce de dos géneros, sin embargo se encontró que Citrus tenía la mayor diversidad genética debido a que este grupo se compone de mandarinas, naranjas, limones, pomelos, híbridos, y otros materiales que no están incluidos dentro de los grupos anteriores. El análisis de similitud permite apreciar que los materiales del banco de germoplasma se encuentran dispersos en el dendrograma, no obstante las introducciones trifoliadas se agrupan y algunas de Citrus tienden a formar agrupaciones. La técnica RAM's tiene potencial en futuros estudios de distancia genética entre los patrones de cítricos estudiados y las principales copas usadas comercialmente, los cuales permitirán entender varios fenómenos como la compatibilidad y afinidad copa-patrón, y el impacto que esto tiene en el desarrollo del árbol.
REFERENCIAS
[1] ORDUZ, R.J.O y MATEUS, C.D.M. Generalidades de los cítricos y recomendaciones agronómicas para su cultivo en Colombia [online]. 2012. Disponible: http://hdl.handle.net/10567/561 [citado 4 de Agosto de 2015] [ Links ].
[2] HYNNIEWTA, M., KUMAR, M.S. and RAMA, R.S. Genetic diversity and phylogenetic analysis of Citrus (L) from north-east India as revealed by meiosis, and molecular analysis of internal transcribed spacer region of rDNA. Meta Gene, 2, 2014, p. 237-251. [ Links ]
[3] GANOPOULOS, I., XANTHOPOULOU, A., MASTROGIANNI, A., DROUZAS, A., KALIVAS, A., BLETSOS, F., KROMMYDAS, K., RALLI, P., TSAFTARIS, A. and MADESIS, P. High Resolution Melting (HRM) analysis in eggplant (Solanum melongena L.): A tool for microsatellite genotyping and molecular characterization of a Greek Genebank collection. Biochemical Systematics and Ecology, 58, 2015, p. 64-71. [ Links ]
[4] POONDLA, A.V., BANDIKARI, A.R., SUBRAMANYAM, B.R.A. and SARATHI, O. V.R. Low temperature active pectinases production by Saccharomyces cerevisiae isolate and their characterization. Biocatalysis and Agricultural Biotechnology, 4, 2015, p. 70-76. [ Links ]
[5] UZUN, A., YESILOGLU, T., AKA-KACAR, Y., TUZCU, O. and GULSEN, O. Genetic diversity and relationships within Citrus and related genera based on sequence related amplified polymorphism markers (SRAPs). Scientia Horticulturae, 121, 2009, p. 306-312. [ Links ]
[6] HYNNIEWTA, M., KUMAR, M.S. and RAMA, R.S. Genetic diversity and phylogenetic analysis of Citrus (L) from north-east India as revealed by meiosis, and molecular analysis of internal transcribed spacer region of rDNA. Meta Gene, 2, 2014, p. 237-251. [ Links ]
[7] LEÓN, J. Botánica de los cultivos tropicales. 3 ed. San José (Costa Rica): Iica/Catie, 1987, p. 84. [ Links ]
[8] LI, Y., CHEN, W., MA, D. and WU, Y. cDNA-AFLP. Ana lysis revealed genes potentially implicated in Catharanthus roseus flowers during wheat blue dwarf phytoplasma infection. Physiological and molecular plant pathology, 84, 2013, p. 1-9. [ Links ]
[9] MORILLO, A.C., MORILLO, Y., CHAGÜEZA, Y., CAICEDO, A. y MUÑOZ, J.E. Caracterización de la diversidad genética en naranja y comparación del polimorfismo de microsatélites amplificados al azar (RAMs) usando electroforesis de poliacrilamida y agarosa. Acta Agronómica, 58(4), 2009, p. 234-244. [ Links ]
[10] MORILLO, Y., MORILLO, A.C., CHAGÜEZA, Y., CAICEDO, A. y MUÑOZ, J.E. Caracterización mediante microsatélites de 34 introducciones de naranja Citrus sinensis L. Osbeck del Banco de Germoplasma de Corpoica-Palmira. Acta Agronómica, 59(1), 2010, p. 9-19. [ Links ]
[11] MUÑOZ, J.E., MORILLO, A.C. y MORILLO, Y. Microsatélites amplificados al azar (RAM) en estudios de diversidad genética vegetal. Acta agronómica, 57(4), 2008, p. 219-226. [ Links ]
[12] DELLAPORTA, S.L., WOOD, J. and HICKS, J.B. A plant DNA minipreparation: Versión II. Plant. Molecular Biology Reports, 14, 1983, p. 19-21. [ Links ]
[13] MAURYE, P., BASU, A. and GUPTA, A. Simple and cost effective apparatus for silver staining of polyacrylamide gels with sequential reagents addition and real time monitoring. Bioscience and Bioengineering, 117, 2014, p. 769-774. [ Links ]
[14] BERG, E. and HAMRICK, J. Quantification of genetic diversity at allozyme loci. Canadian of Forest Research, 27, 1997, p. 415-424. [ Links ]
[15] NEI, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89, 1978, p. 583-590. [ Links ]
[16] NEI, M. Analysis of genetic diversity in subdivided populations. Proceedings of the National Academy of Sciences, 70, 1973, p. 3321-3326. [ Links ]
[17] MEDINA, F.H.P., BORDIGNON, R. and SIQUEIRA, W.J. Segregações gaméticas de locos isoenzímicos em porta-enxertos de citros e suas contribuições alélicas na formação de híbridos. Bragantia, 62, 2013, p. 357-367. [ Links ]
[18] YAHMEDA, J.B.B., NOVILLO, B.P., GARCIA, B.A., TALONB, M. and MORILLON, R. Salt tolerance traits revealed in mandarins (Citrus reticulata Blanco) are mainly related to root-to-shoot Cl- translocation limitation and leaf detoxification processes. Scientia Horticulturae, 191, 2015, p. 90-100. [ Links ]
[19] SANGA, W., HUANGB, Z.R., YANGA, L.T., GUOA, P. and CHENA L.S. An investigation of boron-toxicity in leaves of two citrus species differing in boron-tolerance using comparative proteomics. Proteomics, 123, 2015, p. 128-146. [ Links ]
[20] WANG, L.Q., YANG, L.T., GUO, P., CHEN, E.J. and CHEN, L.S. Leaf cDNA-AFLP analysis reveals novel mechanisms for boron-induced alleviation of aluminum-toxicity in Citrus grandis seedlings. Ecotoxicology and environmental safety, 120, 2015, p. 349-359. [ Links ]
[21] MAHJBI, A., BARAKET, G., OUESLATI, A. and SALHI-HANNACHI, A. Start Codon Targeted (SCoT) markers provide new insights into the genetic diversity analysis and characterization of Tunisian Citrus species. Biochemical Systematics and Ecology, 61, 2015, p. 390-398. [ Links ]
[22] WANG, J., HAO, H., LIU, R., MA, Q., CHENG, Y. and DENG, X. Comparative analysis of surface wax in mature fruits between Satsuma mandarin (Citrus unshiu) and 'Newhall' navel orange (Citrus sinensis) from the perspective of crystal morphology, chemical composition and key gene expression. Food Chemistry, 153, 2014, p. 177- 185. [ Links ]
[23] BASTOS, P.M.D., RAYNAUD, M., VENANCIO, A.L., BASSANEZI, R.B. and REDON, R. Identification of citrus varieties using laser-induced fluorescence spectroscopy (LIFS). Computers and Electronics in Agriculture, 95, 2013, p. 11-18. [ Links ]
[24] HALL, D.G., JUSTIN, G. and STEPHEN, L.L. Further investigations on colonization of Poncirus trifoliata by the Asian citrus psyllid. Crop Protection, 72, 2015, p. 112-118. [ Links ]
[25] DISTEFANO, G., CIRILLI, M., THOMAS, B. and MULEO, R. Ectopic expression of Arabidopsis phytochrome B in Troyer citrange affects photosynthesis and plant morphology. Scientia Horticulturae, 159, 2013, p. 1-7. [ Links ]
[26] ALBRECHT, U., GREG, M.C. and BOWMAN, K.D. Influence of rootstock variety on Huanglongbing disease development in field-grown sweet orange (Citrus sinensis [L.] Osbeck). Scientia Horticulturae, 138, 2012, p. 210-220. [ Links ]
[27] RODRIGUEZ, M.J., ALQUÉZAR, B., ALÓS, E. and ZACARÍAS, L. Biochemical bases and molecular regulation of pigmentation in the peel of Citrus fruit. Scientia Horticulturae, 163, 2013, p. 46-62. [ Links ]

