










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Ciencias de la Salud
Print version ISSN 1692-7273
Rev. Cienc. Salud vol.10 no.3 Bogotá Sept./Dec. 2012
Correlación genotipo-fenotipo y análisis molecular en pacientes con síndrome Down
Correlation genotype-phenotype and molecular analysis in patients with Down syndrome
Correlação genótipo-fenótipo e análise molecular em pacientes com síndrome de Down
Nora Constanza Contreras Bravo, MSc1, Claudia Tamar Silva Aldana, MSc1, Heidi Eliana Mateus Arbelaez, MD, MSc1
1 Unidad de Genética, Instituto de Ciencias Básicas, Universidad Colegio Mayor del Rosario.
Correspondencia: ncontrer@urosario.edu.co
Recibido: junio 24 de 2011 • Aceptado: octubre 22 de 2012.
Para citar este artículo: Contreras NC, Silva CT, Mateus HE. Correlación genotipo-fenotipo y análisis molecular en pacientes con síndrome Down.
Resumen
El síndrome Down (SD) es la trisomía más común en humanos, presentándose en 1 de cada 745 nacidos vivos y es la causa más frecuente de retardo mental. El origen más observado de la trisomía es una no disyunción meiótica (95%), la cual generalmente es de origen materno, mientras un 5% se debe a errores post-cigóticos mitóticos. Objetivo: identificar el origen parental del cromosoma 21 extra, el momento del error no disyuncional y establecer una correlación entre estos eventos y las manifestaciones fenotípicas de los pacientes afectados. Materiales y métodos: se estudiaron cincuenta familias con un hijo con SD mediante el uso de cinco short tandem repeats (STR) a lo largo de 21q, se construyeron los haplotipos de cada paciente y sus padres, determinando el origen parental y el momento en que surgió el error no disyuncional. Resultados: en 80% de las familias el error fue en meiosis I y 20% en la meiosis II; 98% de los cromosomas adicionales fue de origen materno y 2% paterno. Se encontró correlación genotipo-fenotipo en ocho características estudiadas: cuello corto y ancho, tercera fontanela, labio inferior prominente, paladar estrecho y corto, raíz del hélix cruzando la concha, alopecia, pliegue único palmar y otras anomalías como nevus y xeroderma y eventos de recombinación en 24,5% de las familias analizadas. Conclusiones: la edad materna y la variación en el número de recombinaciones está asociada con no disyunciones meióticas I y II; se encontró correlación entre el momento del error no disyuncional y algunas variables clínicas.
Palabras clave: síndrome Down, trisomía, recombinación, no disyunción genética, PCR.
Abstract
Down Syndrome (DS) is the most common trisomy in human beings. Its incidence is estimated in one of 745 live births. On a global scale, it is the most frequent cause of mental retardation. The origin of this trisomy is due to a meiotic non-disjunction in about 95% of cases and is usually maternal, especially in women above 35 years of age. The remaining 5% is due to errors in post-zygotic mitosis. Objective: identify the parental origin of the extra chromosome 21, when the error is not disyuncional and establish a correlation between these events and phenotypic manifestations of the patients affected. Materials and methods: we studied fifty families with a child with DS, using 5 STRs markers along 21q which allowed identification of the origin of chromosome 21 additional parents, the time when the error occurred and recombination presents. The statistical analysis was done using the package SPSS version 15.0 for Windows. Results: in 80% of households in the error was meiosis I and 20% in meiosis II, 98% of the additional chromosomes was home maternal and paternal 2% similar to those reported by other authors, correlation was found genotype-phenotype characteristics studied at 8, neck short and wide, third fontanel, prominent lower lip, palate narrow and short, crossing hélix root of the shell, alopecia, single palm crease and other anomalies as nevi and xeroderma and recombination events in 24,5% of the families tested. Conclusions: the maternal age and variation in the number of recombination is not associated with disjunctions meiotics I and II genotype phenotype correlation was found, but the sample size should be expanded in order to establish with certainty that the correlations.
Keywords: Down syndrome, trisomy, recombination, nondisjunction genetic, PCR.
Resumo
O síndrome de Down (SD) é a trissomia mais comum em humanos, apresentando-se em 1 de cada 745 nascidos vivos e é a causa mais frequentes de retardo mental. A origem mais observada da trissomia é uma não-disjunção meiótica (95%), a qual geralmente é de origem materna, enquanto um 5% se deve a erros pós-zigoticos mitóticos. Objetivo: identificar a origem parental do cromossoma 21 extra, o momento do erro não-disjuncional e estabelecer uma correlação entre estes eventos e as manifestações fenotípicas dos pacientes afetados. Materiais e métodos: se estudaram cinquenta famíliascom um filho com SD mediante o uso de cinco short tandem repeats (STR) ao longe de 21q, se construíram os haplótipos de cada paciente e seus pais, determinando a origem parental e o momento em que surgiu o erro não-disjuncional. Resultados: em 80% das famílias o erro foi em meiose I e 20% na meiose II; 98% dos cromossomas adicionais foi de origem materno e 2% paterno. Encontrou-se correlação genótipo-fenótipo em oito características estudadas: pescoço curto e amplo, terceira fontanela, lábio inferior proeminente, paladar apertado e curto, raiz da hélix a través da concha, alopecia, prega palmar única e outras anomalias como nevus e xeroderma e eventos de recombinação em 24,5% das famílias analisadas. Conclusões:a idade materna e a variação no número de recombinações está associada com não-disjunções meióticas I e II; encontrou-se correlação entre o momento do erro não disjuncional e algumas variáveis clínicas.
Palavras chave: síndrome de Down, trissomia, recombinação, não-disyunção genética, PCR.
El síndrome Down (SD) es el mejor y más conocido de los desórdenes cromosómicos y es la causa genética más común de retardo mental moderado, representando 10% de todos los casos en Estados Unidos (1-3). La incidencia de esta aneuploidía autosómica se ha estimado en 1 por cada 745 nacimientos. La prevalencia en la población general es de 1:2000 a 1:3300 (1, 4, 5).
En Colombia se han realizado algunos estudios encaminados a determinar la incidencia de la enfermedad, reportando un valor de 1,5 por cada 1.000 nacidos vivos para Cali y de 5 en cada 10.000 nacidos vivos en San Juan de Pasto (6, 7). Un tercer reporte del Estudio Colaborativo Latinoamericano de Malformaciones Congénitas describe una incidencia de 17,4 por cada 10.000 nacidos vivos entre 1998 y 2007 en Colombia (8).
El diagnóstico clínico se hace generalmente en el nacimiento ante los hallazgos fenotípicos clásicos de la entidad, que incluyen hipotonía, talla baja, braquicefalia con occipucio relativamente plano, fisuras parpebrales oblicuas, nariz pequeña con puente nasal deprimido, tendencia a mantener la boca abierta y protruir la lengua, hiperflexibilidad de las articulaciones, cuello corto y con piel redundante, pliegue único palmar y clinodactilia del quinto dedo. Cerca de 40% cursan con cardiopatías congénitas, entre otros hallazgos.
En 95% de los casos el SD es resultado de eventos no disyuncionales durante la meiosis; un 5% restante se debe a traslocaciones entre cromosomas acrocéntricos o a eventos de mosaicismo originados por errores no disyuncionales post-cigóticos mitóticos o fenómenos de corrección de trisomías (9-11).
De los eventos no disyuncionales, 90% de los casos son de origen materno; dentro de este grupo, 78% ocurre en la primera división meiótica. En contraste, 60% de las trisomías 21 de origen paterno son secundarias a no disyunción en la segunda división meiótica (12, 13).
Si la no disyunción ocurrió durante la meiosis I, el probando recibe una copia de cada uno de los cromosomas parentales no disyuncionados. Esto se conoce como heterocigocidad disómica o no reducida, pero si la no disyunción ocurre en la meiosis II, el probando recibe dos copias idénticas del mismo cromosoma parental, lo cual se denomina homocigocidad disómica o reducida (14-16) (figura 1).
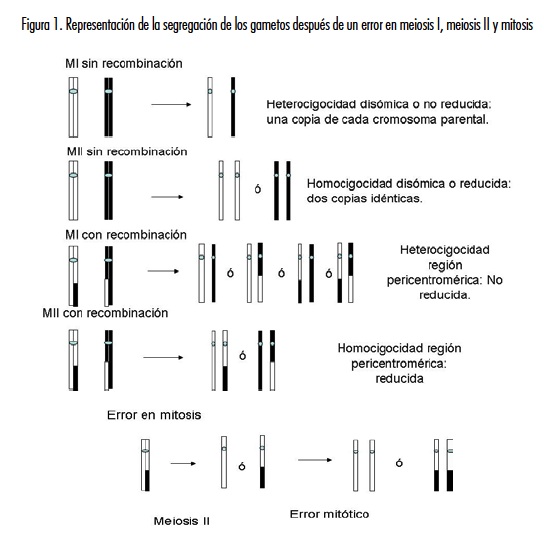
En el cromosoma 21 se han descrito varias repeticiones cortas en tandem o STR (Short Tandem Repeats), específicos para varios loci dentro del cromosoma 21 (17-20). La disponibilidad de estos marcadores ha permitido establecer el origen parental del cromosoma 21 extra y en qué etapa de la meiosis se presentó la no disyunción (12, 21-23).
En el presente estudio, mediante STR se busca identificar el origen parental del cromosoma 21 extra, el momento en el cual surge el error no disyuncional y la correlación entre estos eventos y las manifestaciones fenotípicas de los pacientes afectados.
Materiales y métodos
Se estudiaron cincuenta niños con diagnóstico clínico de SD, confirmado citogenéticamente con trisomía 21 libre y sus padres, atendidos en el servicio de Genética de la Universidad del Rosario durante el lapso de 2004-2007.
Se tomaron muestras de sangre periférica a cada uno de los pacientes y sus padres, previo consentimiento informado aprobado por el Comité de Ética de la Universidad del Rosario. Se realizó la extracción del ADN genómico por la técnica de Salting-Out. Mediante PCR se amplificaron los STR D21S11, D21S1411, D21S1432 D21S1412 y D21S1414, utilizando primers descritos previamente, marcados con CY5, utilizando las siguientes condiciones: 100 ng de ADN; 1,2 U de Taq polimerasa; 1 X de Buffer para PCR (166 mM (NH4)2SO4, 670 mM Tris.HCl, pH 8,8); MgCl2 6 mM; dNTPs 0,6 mM y 5 pmol/ul de cada uno de los primers (17-20) (tabla 1). Los productos de PCR fueron analizados en el secuenciador semiautomático ALFexpress®, el cual, por medio del análisis de curvas y su área, permite determinar la dosis génica de cada uno de los STR en los pacientes y sus padres (24).
Con base en los haplotipos se determinó el estado de monoalélico, dialélico y trialélico para cada uno de los sistemas STR utilizados (figura 2).
La depuración y procesamiento de los datos se hizo utilizando el paquete SPSS versión 15.0 para Windows, la correlación específica entre el genotipo y el fenotipo de las variables discretas se determinó utilizando test exacto de Fisher y para las variables descriptivas se hizo un Anova de un factor.
Resultados
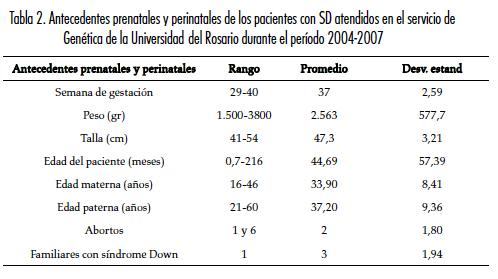
El grupo de pacientes con SD estaba conformado por veintiséis niñas y veinticuatro niños. Las características demográficas y los antecedentes prenatales de las familias analizadas se resumen en la tabla 2.
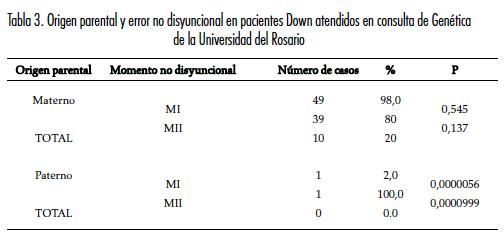
De los cincuenta tríos estudiados, en 98,0% de los casos el cromosoma 21 extra fue de origen materno; de estos, un 80% fue provocado en MI y 20% en MII. El origen fue paterno en solo una de las familias analizadas (2%) y el error se produjo en MI (tabla 3).
En doce familias (24,5%) se presentaron eventos de recombinación, los cuales fueron evaluados con base en los intervalos establecidos sobre el cromosoma 21. El intervalo proximal (IP) está delimitado por los marcadores D21S1432 y D21S11; el intervalo medio (IM) comprendido entre D21S11 y D21S1412 e incluye el STR D21S1414 y el intervalo distal (ID) entre D21S1412 y D21S1411. Del total de recombinaciones observadas, tres ocurrieron en IP en MII, tres en IM en MI, seis en ID, dos en MI y cuatro en MII (figura 3).
Al establecer una correlación entre el fenotipo presente en los pacientes analizados con el momento en que ocurrió la mala segregación de los cromosomas, se encontró que: el cuello corto y ancho era más frecuente en pacientes con error en MI (MI 75,6% vs. MII 44,4%), mientras que otras características como labio inferior prominente (MI 31,7% vs. MII 66,6%), tercera fontanela (MI 2,4% vs. MII 22,2%), paladar estrecho y corto (MI 15% vs. MII 44,4%), raíz del hélix cruzando la concha (MI 0% vs. MII 22,2%), alopecia (MI 0% vs. MII 22,2%), otras anomalías como nevus y xeroderma (MI 2,4% vs. MII 22,2%) y pliegue único palmar (MI 55,2% vs. MII 66,6%) se encontraban asociados con errores en MII (p < 0,1) (tabla 4).
Los valores de heterocigocidad encontrados para cada uno de los STR analizados oscilan entre 73 y 87% (datos no publicados), sin mostrar diferencias con los valores ya reportados en la literatura.
La frecuencia de eventos no disyuncionales en MI va aumentando en forma directamente proporcional a la edad materna. En mujeres menores de 25 años es de 17,1%; entre 35 y 39 años de 26,8% y en mayores de 40 años de 31,7%, mientras los eventos en MII se distribuyen con mayor frecuencia en los extremos de los rangos de edad analizados: mujeres menores de 25 años y mayores de 40, 57,1% y 28,6% respectivamente; sin embargo, estas diferencias no son significativas (p > 0,05).
Discusión
El SD es uno de los desórdenes cromosómicos más comunes y es la causa genética más frecuente de retardo mental. Esta alteración es producida en 95% de los casos por una trisomía 21 libre secundaria a un evento no disyuncional durante la meiosis I o II. Un 5% restante se debe a mosaicismo por errores no disyuncionales después de la segunda división mitótica, a eventos de corrección de aneuploidías o a translocaciones robertsonianas (25, 26).
El análisis del origen del SD en los pacientes examinados indicó que 98% de las no disyunciones fueron de origen materno y 2% de origen paterno. Dentro de las no disyunciones maternas, 80% ocurrió por errores en la MI. Estos hallazgos muestran una distribución similar a los reportes previos de la literatura (27, 28).
La edad materna ha sido considerada como un factor de riesgo para SD. En el presente estudio se observó que 56% de las madres analizadas eran mayores de 35 años y, de estas, 57,1% era mayor de 40 años. Aunque esta asociación entre la edad materna y la frecuencia de concepciones aneuploides ha sido reconocida durante un siglo, la razón del aumento de errores meióticos en los ovocitos dependiente de la edad de la mujer es aún desconocida. Se han postulado diferentes hipótesis para explicar este evento; sin embargo, las más aceptadas se relacionan con la detención del ovocito en la profase I de la meiosis durante el desarrollo embrionario hasta el momento de la ovulación y con ello la acumulación de un sin número de alteraciones que afectan la cohesión de las cromátides hermanas, la recombinación de los cromosomas homólogos y el control que se hace mediante el punto de chequeo de la fase M (29, 30).
La frecuencia de eventos no disyuncionales en MI va aumentando en forma directamente proporcional a la edad materna, mientras los eventos en MII son más frecuentes en las mujeres menores de 25 años y en las mayores de 40 años. A pesar de que estas diferencias no son significativas, esta tendencia ha sido reportada en otros trabajos (28, 31).
Al analizar los haplotipos se encontraron eventos de recombinación en 24,5% de las familias estudiadas. Esta baja tasa de recombinación se correlaciona con estudios previos que relacionan la disminución en la recombinación con el aumento en la frecuencia de eventos no disyuncionales (32). Sin embargo, cabe resaltar que es posible que por las distancias entre los marcadores utilizados algunas recombinaciones no hayan sido detectadas.
Luego de correlacionar el fenotipo con el momento de la no disyunción se encontraron diferencias significativas en ocho rasgos (cuello corto y ancho, tercera fontanela, labio inferior prominente, paladar estrecho y corto, raíz del hélix cruzando la concha, alopecia, otras anomalías de piel y cabello entre las que se encontraban nevus, xeroderma y manchas mongoloides y pliegue único palmar). De estas características, el cuello corto y ancho se encontró asociado con error en MI y los demás con un error en MII (p < 0,1). Es importante recordar que si el error surgió en la primera división el gameto recibe dos cromosomas 21 diferentes (par de homólogos) y si fue en la segunda división recibe cromátidas hermanas; al ser fecundados estos gametos se producirá un individuo con los dos cromosomas homólogos o con dos cromosomas idénticos de uno de sus padres, llevando a que la expresión de los genes en cada uno de estos casos sea diferente, pudiendo esta expresión diferencial afectar el fenotipo del paciente. No obstante, ante la gran diferencia existente entre el número de casos en MI y MII, sería necesario aumentar el tamaño de la muestra para confirmar esta asociación.
En conclusión, las técnicas de biología molecular permiten establecer el origen parental de la no disyunción mediante el uso de STR sobre el cromosoma 21 y el momento cuando se originó la no disyunción meiótica; en la población analizada se encontraron valores similares a los reportados en la literatura. Algunas características clínicas analizadas se relacionaron con el momento del evento no disyuncional; sin embargo, el tamaño de la muestra debe ser ampliado para determinar si existe una asociación real entre estas características.
Agradecimientos
A las Fundaciones Fe y Síndrome Down de las ciudades de Bogotá y Neiva respectivamente por permitirnos estudiar a sus pacientes; a los pacientes y a sus padres por la buena disposición en la realización de este estudio; al profesor Milciades Ibáñez por su ayuda en la parte estadística y a la Universidad del Rosario por su apoyo en la realización de este trabajo.
Descargos de responsabilidad
Los autores declaran que no tienen ningún conflicto de interés.
Bibliografía
1. Fryns JP. Chromosome 21. Trisomy 21. En: Buyse ML (Ed.). Birth defects encyclopedia. Cambridge: Blackwell Scientific Publications; 1990. p. 391-3. [ Links ]
2. Thompson MV, McInnes R, Willard HF. Clinical cytogenetics: general principles and autosomic abnormalities. En: Genetics in Medicine. 5th. ed. Philadelphia: Saunders Elsevier; 1991. p. 209-15. [ Links ]
3. Jorde LB, Carey JC, White RL. Citogenética clínica: bases cromosómicas de la enfermedad del ser humano. Trisomía 21. Barcelona: Genética Médica 1996. [ Links ]
4. De Grouchy J. 21p- maternal en double exemplaire chez un trisomique 21. Ann Genet. 13: 35-42. [ Links ]
5. Korenberg JR, Chen X-N, Schipper R, Sun Z, Gonsky R, Gerwehr S et al. Down Syndrome phenotypes: the consequences of chromosomal imbalance. Proc Natl Acad Sci USA 1994; 91 (11): 4997-5001. [ Links ]
6. Ramírez RE, Isaza C, Gutiérrez MI. La incidencia del síndrome de Down en Cali. Colomb Méd 1996; 27 (3-4): 138-42. [ Links ]
7. Hernández I, Manrique RC. Prevalencia de síndrome de Down en Cehani-ESE, San Juan de Pasto Colombia 1998-2003. Nova 2006; 4 (5): 50-6. [ Links ]
8. Nazer J, Cifuentes L. Malformaciones congénitas en Chile y Latinoamérica: una visión epidemiológica del ECLAMC del período 1995-2008. Rev Med Chil 2011; 139 (1): 72-8. [ Links ]
9. Wolstenholme J, Angell RR. Maternal age and trisomy -a unifying mechanism of formation. Chromosoma 2000; 109 (7): 435-8. Corregido en: Chromosoma 2001; 110 (2): 130. [ Links ]
10. Wilkinson TA, James RS, Crolla JA, Cockwell AE, Temple IK. A case of maternal uniparental disomy of chromosome 9 in association with confined placental mosaicism for trisomy 9. Prenat Diagn 1996; 16 (4): 371-4. [ Links ]
11. Aleyasin A, Mohammad-Ganji S, Ghazanfari M, Jahanshad F. Parental origin of meiotic error of the extra chromosome 21 as indicated by short tandem repeat (STR) polymorphisms in Down sindrome. Arch Iranian Med 2004; 7 (2): 118-121. [ Links ]
12. Antonarakis SE, Petersen MB, McInnis MG, Adelsberg PA, Schinzel AA, Binkert F et al. The meiotic stage of nondisjunction in trisomy 21: determination by using DNA polymorphisms. Am J Hum Genet 1992; 50 (3): 544-50. [ Links ]
13. Sherman SL, Freeman SB, Grantham M, Peters J, Jacobs PA, Kurnit DM et al. Non-disjunction of trisomy 21: comparison of centromere maps resulting from maternal meiosis I and II non-disjunction. Am J Hum Genet 1992; 51 (suppl.): A24. [ Links ]
14. Sherman SL, Takeaesu N, Freeman SB, Grantham M, Phillips C, Blackston RD et al. Trisomy 21: association between reduced recombination and nondisjunction. Am J Hum Genet 1991; 49 (3): 608-20. [ Links ]
15. Pangalos C, Avramopoulos D, Blouin JL, Raoul O, DeBlois MC, Prieur M et al. Understanding the mechanism(s) of Mosaic Trisomy 21 by using DNA polimorphism analysis. Am J Hum Genet 1994; 54 (3): 473-81. [ Links ]
16. Zittergruen MM, Murray JC, Laurer RM, Burns TL, Sheffield VC. Molecular analysis of nondisjunction in Down Syndrome patients with and without atrioventricular septal defects. Circulation 1995; 92 (10): 2803-10. [ Links ]
17. Peterson MB, Frantzen M, Antonarakis SE, Warren AC, Van Broeckhoven C, Chakravarti A et al. Comparative study of microsatellite and cytogenetic markers for detecting the origin of the nondisjoined chromosome 21 in Down syndrome. Am J Hum Genet 1992; 51 (3): 516-25. [ Links ]
18. Pertl B, Weitgasser U, Kopp S, Kroisel PM, Sherlock J, Adinolfi M. Rapid detection of trisomies 21 and 18 and sexing by quantitative fluorescent multiplex PCR. Hum Genet 1996; 98 (1): 55-9. [ Links ]
19. Verma L, McDonald F, Leedham P, McConachie M, Dhanjal S, Hulten M. Rapid and simple prenatal DNA diagnosis of Down's syndrome. Lancet 1998; 352 (9121): 9-12. [ Links ]
20. Sherlock J, Cirigliano V, Petrou M, Tutschek B, Adinolfi M. Assessment of diagnostic quantitative fluorescent multiplex polymerase chain reaction assays performed on single cells. Ann Hum Genet 1998; 62 (Pt 1): 9-23. [ Links ]
21. Lamb NE, Yu K, Shaffer J, Feingold E, Sherman S. Association between maternal age and meiotic recombination for trisomy 21. Am J Hum Genet 2005; 76 (1): 91-9. [ Links ]
22. Antonarakis SE, Avramopoulos D, Blouin JL, Talbot CC Jr, Schinzel AA. Mitotic errors in somatic cells cause trisomy 21 in about 4,5% of cases and are not associated with advanced maternal age. Nat Genet 1993; 3 (2): 146-50. [ Links ]
23. Samura O, Sohda S, Johnson KL, Pertl B, Ralston S, Delli-Bovi L, Bianchi DW. Diagnosis of trisomy 21 in fetal nucleated erythrocytes from maternal blood by use of short tandem repeat sequences. Clin Chem 2001; 47 (9): 1622-6. [ Links ]
24. De Grouchy J, Turleau C. Autosomal disorders. En: Principles and practice of medical genetics. Edimburgo: Churchill Livingstone; 1990. p. 252-6. [ Links ]
25. Kuliev A, Verlinsky Y. Meiotic and mitotic nondisjunction: lessons from preimplantation genetic diagnosis. Hum Reprod Update 2004; 10 (5): 401-7. [ Links ]
26. Stoll C, Alembik Y, Rothemp MP. Study of DS in 238942 consecutive births. Ann Genet 41(1): 44-51. [ Links ]
27. Valle Y, Hernández G, Barajas-Barajas LO, Vázquez-Herrera JA, Ortiz R, Padilla-Gutiérrez JR. Origen meiótico y parental del cromosoma 21 extra en niños con trisomía 21 regular. Rev Med Inst Mex Seguro Soc 2006; 44 (5): 463-8. [ Links ]
28. Ramírez NJ, Belalcázar HM, Yunis JJ, Quintero LN, Arboleda GH, Arboleda H. Parental origin, nondisjunction, and recombination of the extra chromosome 21 in Down syndrome: a study in a sample of the Colombian population. Biomédica 2007; 27 (1): 141-8. [ Links ]
29. Hamatani T, Falco G, Carter MG, Akutsu H, Stagg CA, Sharov AA et al. Age-associated alteration of gene expression patterns in mouse oocytes. Hum Mol Genet 2004; 13 (19): 2263-78. [ Links ]
30. Steuerwald NM, Bermúdez MG, Wells D, Munné S, Cohen J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod Biomed Online 2007; 14 (6): 700-8. [ Links ]
31. Schinzel AA, Adelsberger PA, Binkeert F, Basaran S, Antonarakis SE. No evidence for a paternal interchromosomal effect from analysis of the origin of nondisjunction in Down Syndrome patients with concomitant familial chromosome rearrangements. Am J Hum Genetics 1992; 50 (2):288-93. [ Links ]
32. Warren AC, Chakravarti A, Wong C, Slaugenhaupt SA, Halloran SL, Watkins PC et al. Evidence for reduced recombination on the nondisjoined chromosomes 21 in Down syndrome. Science 1987; 237 (4815): 652-4. [ Links ]
