










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Ciencias de la Salud
Print version ISSN 1692-7273
Rev. Cienc. Salud vol.12 no.3 Bogotá Sept./Dec. 2014
https://doi.org/dx.doi.org/10.12804/revsalud12.03.2014.01
Doi: dx.doi.org/10.12804/revsalud12.03.2014.01
Defectos en la expresión de cadena zeta (ζ) en un grupo de pacientes con lupus, escleroderma y artritis de inicio tardío, Colombia 2014
Defects in the Zeta Chain Expression (ζ) in a Group of Patients with SLE, Scleroderma and Late-onset Arthritis, Colombia 2014
Defeitos na expressão de cadeia zeta (ζ) em um grupo de pacientes com Lúpus, Esclerodermia e artrite de início tardio, Colômbia 2014
Heber Siachoque-Montañez1 Milcíades Ibáñez-Pinilla2, Antonio Iglesias-Gamarra2
1 Escuela de Medicina y Ciencias de la Salud, Universidad del Rosario, Bogotá D. C., Colombia. Correspondencia: heber.siachoque@urosario.edu.co
2 Facultad de Medicina, Universidad Nacional de Colombia. Bogotá D. C., Colombia.
Recibido 7 de octubre de 2013 • Aceptado: 21 de julio de 2014
Para citar este artículo: Siachoque-Montañez H, Ibáñez-Pinilla M, Iglesias-Gamarra A. Defectos en la expresión de cadena zeta (ζ) en un grupo de pacientes con lupus, escleroderma y artritis de inicio tardío, Colombia 2014. Rev Cienc Salud. 2014;12(3): 303-18. doi: dx.doi.org/10.12804/revsalud12.03.2014.01
Resumen
Introducción: El lupus eritematoso sistémico (LES), la escleroderma y la artritis de inicio tardío son enfermedades inflamatorias autoinmunes (EIA) caracterizadas por la producción de autoanticuerpos y presencia de células T anormales, las cuales generan respuesta inmune defectuosa. La expresión anormal de moléculas clave en señalización y la función defectuosa de los linfocitos T cumplen un papel significativo en la patogénesis de la enfermedad autoinmune. Las células T muestran numerosas anormalidades en la señalización del complejo receptor de célula T cadena zeta (TCRζ) estas aberraciones resultan en la alteración de la expresión de citoquinas y algunos eventos bioquímicos involucrados en la expresión de moléculas de superficie. Los defectos en el complejo TCRζpueden estar asociados al uso de corticoides, utilizados en pacientes con enfermedad autoinmune, debido a su potente actividad antiinflamatoria y propiedades inmunosupresoras. Los corticoides sintéticos como la dexametasona inhiben la actividad trascripcional de algunos factores como NFkB y AP-1 los cuales regulan la síntesis de algunas citoquinas y podrían estar involucrados en la síntesis de TCRζ. Objetivos: Evaluar la asociación de los defectos de la expresión de cadena zeta (ζ), en células T de pacientes con enfermedad autoinmune sin tratamiento con corticoides. Los defectos en la expresión defectuosa de zeta, proteína asociada al receptor TCR, altera la activación normal del linfocito T. Materiales y métodos: Estudio analítico de casos y controles, con una relación 1:1 (13:13). Los casos fueron pacientes con enfermedad autoinmune activa (6 pacientes con LES, 5 pacientes con escleroderma y 2 pacientes con artritis de inicio tardío), y que no hubieran iniciado tratamiento con corticoides, los controles fueron pacientes sin diagnóstico de enfermedad autoinmune. El diagnóstico se hizo a través de los criterios establecidos por el Colegio Americano de Reumatología para pacientes con LES, escleroderma y artritis de inicio tardío. Se realizó venopunción extrayendo 10ml de sangre total, se extrajo RNA total y se hizo RT PCR, se amplificó utilizando un juego de primers que flanquean una región de 138 pares de bases que involucraban los exones 2, 3 y 4 de cadena ζ. Resultados: En los valores de amplificación de cadena Z se encontraron diferencias significativas en pacientes con enfermedad autoinmune (0,8214±0,1787, med=0,7368) comparada con el grupo control (0,9225±0,1272, med=0,9830) (p=0,045, Test no-paramétrico, exacto de Mann Whitney a una cola). Conclusión: La expresión de cadena Z se encontró disminuida en células T de pacientes con enfermedad autoinmune, sin uso de corticoides, la disminución o una falta de expresión de la cadena Z puede causar alteraciones en la respuesta inmune.
Palabras clave: Corticoides cadena ζ, células T, splicing, lupus eritematoso sistémico (LES), receptor de la célula T (TCR), fracción cristalizable de IgG(Fcγ), citoquinas, autoinmunidad.
Abstract
Introduction: Systemic Lupus Erythematosus (SLE), Scleroderma and late-onset arthritis are autoimmune inflammatory diseases (EIA) characterized by autoantibody production and presence of abnormal T cells which generate defective immune response. The abnormal expression of key signaling molecules in the defective function of T-lymphocytes plays a significant role in the pathogenesis of autoimmune disease.The T-cells exhibit numerous abnormalities TCRζ1 signaling complex, these aberrations result in altered expression of cytokines and some biochemical events involved in the expression of surface molecules. Defects in the complex may be associated TCR ζ to steroids used in autoimmune disease patients due to their powerful anti- inflammatory activity and immunosuppressive properties. The synthetic corticosteroids such as dexamethasone inhibit the transcriptional activity of some factors such as NFKB and AP-1, which regulate the synthesis of certain cytokines and could be involved in the TCRζ synthesis. Material and Methods: A case-control study, with a 1:1 ratio of cases and controls (13:13). Cases were patients with active autoimmune disease (6 patients with SLE, 5 patients with scleroderma and 2 patients with lateonset arthritis), who have not started treatment with corticosteroids. Controls were patients with no autoimmune disease. The diagnosis was made by the criteria established by the American College of Rheumatology for patients with SLE, scleroderma and late-onset arthritis.A 10 mL sample was obtained by venipuncture whole blood. Total RNA was extracted and RT-PCR was performed using a set of primers flanking a region of 138 base pairs involving exons 2, 3 and 4 of the ζchain. Results: The values of Z chain amplification showed significant differences in patients with autoimmune disease (0.8214 ± 0.1787, med = 0.7368) compared with the control group (0.9225 ± 0.1272, med = 0.9830) (p = 0.045, Mann-Withney non-parametric one tailed exact test). Conclusion: We observed a reduced level of in the zeta chain expression in T cells in patients with autoimmune disease without use of corticosteroids.
Key Words: ζ Chain Corticosteroids, T-Cells, Splicing, Lupus Erythematosus (SLE), T-Cell Receptor (TCR), IgG Fraction Crystallizable (Fcγ), Cytokines, Autoimmunity.
Resumo
Introdução: O Lúpus Eritematoso Sistêmico (LES), Esclerodermia e Artite de início tardio são doenças inflamatórias autoimunes caracterizadas por produção de autoanticorpos e presença de células T anormais as quais geram resposta imune defeituosa.A expressão anormal de moléculas chave em sinalização e a função defeituosa dos linfócitos T cumprem um papel significativo na patogênese da doença autoimune.As células T mostram numerosas anormalidades na sinalização do Complexo Receptor de célula T cadeia zeta (TCRζ)1, estas aberrações resultam na alteração da expressão de citocinas e alguns eventos bioquímicos envolvidos na expressão de moléculas de superfície. Os defeitos no complexo TCRζ podem estar associados ao uso de corticoides, utilizados em pacientes com doença autoimune devido a seu potente atividade auto-inflamatória e propriedades imunossupressoras. Os corticoides sintéticos como a dexametasona inibem a atividade transcricional de alguns fatores como NFkB e AP-1 os quais regulam a síntese de algumas citocinas e poderiam estar envolvidas na síntese de TCRζ2, 3. Objetivos: Avaliar a associação dos defeitos da expressão de cadeia zeta (ζ), em células T de pacientes com doença autoimune sem tratamento com corticoides. Os defeitos na expressão defeituosa de zeta, proteína associada ao receptor TCR altera a ativação normal do linfócito T. Materiais e métodos: Estudo analítico de casos e controles, com uma relação 1:1 (13:13). Os casos foram pacientes com doença autoimune ativa (6 pacientes com LES, 5 pacientes com esclerodermia e 2 pacientes com artrite de início tardio), e que não tenham iniciado tratamento com corticoides e os controles foram pacientes sem diagnóstico de doença autoimune. O diagnóstico se fez através dos critérios estabelecidos pelo Colegio Americano de Reumatología para pacientes com LES, esclerodermia e artrite de início tardio. Se realizou venopunção extraindo 10mls de sangue total, se extraiu RNA total e se fez RT PCR, se amplificou utilizando um jogo de primers que flanqueiam uma região de 138 pares de bases que envolviam os exões 2, 3 e 4 de cadeia ζ. Resultados: Nos valores de amplificação de cadeia Z se encontraram diferenças significativas em pacientes com doença autoimune (0.8214±0.1787, med=0.7368) comparada com o grupo controle (0.9225±0.1272, med=0.9830) (p=0.045, Teste não paramétrico, exato de Mann Whitney) Conclusão: A expressão de cadeia Z encontrou-se diminuída em células T de pacientes com doença autoimune, sem uso de corticoides, a diminuição ou uma falta de expressão da cadeia Z pode causar alterações na resposta imune.
Palavras-chave: Corticoides Cadeia ζ, células T, Splicing, Lúpus Eritematoso Sistêmico (LES), Receptor da célula T (TCR), Fração cristalizável de IgG (Fcg), citocinas, autoimunidade.
Introducción
Los mecanismos patológicos precisos desencadenantes de la función anormal de las células T en pacientes con enfermedad autoinmune (EA) no han sido esclarecidos totalmente; la identificación de eventos genéticos fundamentales, así como el estudio de los mecanismos bioquímicos involucrados en señalización intracelular, pueden contribuir al entendimiento de la etiopatogénesis del lupus eritematoso sistémico (LES), además de proveer nuevos blancos terapéuticos para una futura intervención farmacológica (1-3). En vista de este significativo esfuerzo por descubrir la alteración en la expresión de algunos genes que pueden predisponer la aberrante respuesta a autoantígenos y la causa patológica de la producción de auto anticuerpos, se evidenció que la cadena ζ asociada al receptor de la célula T es una molécula clave y vital en la señalización, es crítica para el desarrollo y la función de la célula T; su defecto, en la gran mayoría de pacientes con LES y en otras enfermedades autoinmunes, la podrían ubicar como la causa más importante del desencadenamiento de la enfermedad (4).
Los estudios genómicos realizados recientemente en los que se evidencia pérdida de exones han reportado cierta asociación con la susceptibilidad al desarrollo de enfermedades autoinmunes. Las células T de pacientes con LES expresan además una marcada disminución de transcritos mRNA para cadena ζ y, por consiguiente, en la proteína expresada; sin embargo, los eventos de señalización iniciados por ζTCR/CD3 se incrementan notablemente en células T de pacientes con EA comparados con células T de personas normales (3,4).
La identificación de la deficiencia de cadena ζ en pacientes fue realizada inicialmente por análisis de fosforilación de tirosinas en TCR/ CD3, los cuales revelaron disminución en la expresión de la proteína en células T de pacientes con LES. El déficit en la expresión de cadena ζha sido reportado por varios grupos de investigación, sugiriendo que no está restringido a un grupo poblacional (5-7). La prevalencia de la deficiencia de cadena ζen pacientes con LES entre individuos de diferentes grupos raciales, con diferentes niveles de actividad de la enfermedad determinados por el índice de MEX-SLEDAI, muestra una prevalencia del 78 %, siendo de gran relevancia en la inmunopatogénesis del LES (7).
Aspectos moleculares relacionados con disminución de la activación de los linfocitos están asociados con alteración en las proteínas involucradas en las cascadas de señalización intracelular. El complejo receptor T TCR/CD3 juega un papel crítico en proliferación y síntesis de citoquinas, orquestando en gran parte la respuesta celular y humoral a través de la activación inicial TCR-péptido. El entrecruzamiento del complejo receptor lleva a un incremento de los niveles de calcio intracitoplasmático y fosforilación de tirosinas presentes en proteínas citosólicas (8); esta sobreexcitabilidad de las células T de pacientes con LES puede ser atribuida a la pérdida de la expresión de cadena TCR ζ, la cual es reemplazada por la cadena FcγR en algunos eventos con agregación en las balsas lipídicas en la membrana celular(9).
Los receptores FcγR hacen parte de una familia de proteínas llamadas receptores inmunes reconocedores de cadenas múltiples (MIRR), los cuales se presentan en forma de complejos hetero-oligoméricos de una cadena alfa que unen la inmunoglobulina y otra cadena de señalización. Los dominios citoplasmáticos de los MIRR son de tipo ITAM, es decir, de activación. Los genes que codifican para Fcγ, un locus candidato en la susceptibilidad genética al desarrollo de enfermedades autoinmunes, están localizados en la misma región del gen que codifica para cadena ζ (5). Esta zona se ha caracterizado por desarrollar entrecruzamientos desiguales y rearreglos alternativos, lo que ha dado lugar a duplicaciones y deleciones.
El reemplazo de la cadena ζcausa aberraciones en la cascada de señalización, lo cual puede generar defectos en la síntesis de IL-2; es claro que la disminución en la expresión de ζ es el evento central más relevante en el desencadenamiento de la enfermedad (7).
Papel de los corticoides en la expresión de algunas proteínas
Un aspecto de gran relevancia y que puede estar asociado a la disminución en la expresión de cadena zeta tiene que ver con la utilización de corticoides, los cuales son utilizados en pacientes con EA con el fin de disminuir la reacción inflamatoria. Los glucocorticoides (GC) son hormonas que cumplen múltiples funciones y juegan un papel relevante en la respuesta inmune con efectos tanto reguladores como supresores que actúan sobre poblaciones de linfocitos T y B. Las glándulas suprarrenales son la principal fuente (corticosterona en roedores y cortisol en los seres humanos), son liberados en respuesta a la hormona corticotropina (ACTH) producida por la glándula pituitaria. Los GC son lipofílicos y en sangre se unen a trasportadores séricos como la proteína de unión a corticoides (CBG) y la albúmina (10). Sus efectos inmunosupresores y anti-proliferativos en gran medida son dados a través de la modulación de la transcripción génica (11).A la luz de los nuevos conocimientos, los conceptos sobre los efectos represivos y activadores de los GC dependientes principalmente de ‘transrepresión y de ‘transactivación han cambiado, estudios recientes han demostrado que las acciones antiinflamatorias más relevantes de los GC son causadas por la activación o represión de genes reguladores de citoquinas antiinflamorias (12, 13). Tradicionalmente, las terapias antiinflamatorias se han centrado en el diseño de estrategias dirigidas a disminuir o neutralizar el nivel de mediadores pro inflamatorios e inhibir el reclutamiento de leucocitos y su activación. Estos tratamientos incluyen fármacos antiinflamatorios no esteroideos (AINE), glucocorticoides (GC), receptores agonistas (GCs sintéticos) y anticuerpos o inhibidores dirigidos a citoquinas pro inflamatorias específicas, tales como el factor de necrosis tumoral (TNF-α) y la interleucina-1 (IL-1) (14).Algunos corticoides sintéticos como la dexametasona, potencian la actividad inhibidora del sistema inmune, inhibiendo o activando la actividad de algunos factores de trascripción, interfiriendo en la expresión de algunas proteínas.Aún no son claros los mecanismos que subyacen estos complejos eventos regulatorios (15).
La fracción proteica libre de GC pasa fácilmente a través de la membrana celular hacia el citoplasma en donde se une a receptores de glucocorticoides tipo II. Debido a su efecto inmunosupresor, antiinflamatorio y antialérgico los glucocorticoides sintéticos han sido ampliamente utilizados en el tratamiento de múltiples patologías entre ellas enfermedades de tipo autoinmune, inhibiendo la producción y secreción de algunas citoquinas proinflamatorias (16).
Los GC pueden modular tanto componentes innatos del sistema inmune como adaptativos. Los componentes innatos funcionan como una primera línea de defensa contra los patógenos invasores y los adaptativos son instruidos por eventos inmunes que ya han ocurrido y que responden con la liberación de anticuerpos de alta afinidad dirigidos contra material extraño. Mediante la activación de la apoptosis en ambas células ‘innatas' dendríticas (DC) y ‘adaptativas' linfocitos T, los GR afectan simultáneamente la activación y las funciones efectoras de las células inmunes a través de la manipulación de sus vías transcripcionales (17).
Los efectos de los GC en el desarrollo de otras poblaciones celulares como lo son las células Th17 son desconocidas, en parte debido a que los eventos que conducen a la diferenciación de Th17 no son completamente claros; sin embargo, la evidencia disponible sugiere que indirectamente los GC como la prednisolona que inhibe la polarización de células T naive hacia linfocitos Th17 como resultado de una notable disminución en la expresión de IL-23 por parte de las CDs (18-20). De igual forma, varios grupos han demostrado que la inducción de IL-6 por patógenos se encuentra suprimida por acción de los GC, al igual que los niveles de TGFβ en las articulaciones de ratones artríticos (21).
El papel de las CDs es crítico en la activación de las células Th y cualquier evento que involucre el uso de GC o inmunosupresores puede alterar la diferenciación celular. Se han propuesto poblaciones de CDs de distintos orígenes para regular diferencialmente los subconjuntos de Th. Por ejemplo, células CDs criadas en un entorno con IL-10, IL-6, y TNFα impiden la activación de células Th1 (22).
Cuatro mecanismos diferentes se consideran para mediar los efectos terapéuticos de los GC: los mecanismos genómicos clásicos causados por la activación de los receptores citosólicos GC (cGCR); mecanismos no genómicos secundarios provocados por la activación de los receptores citosólicos GC; efectos no genómicos de los receptores de glucocorticoides unidos a membrana (mGCR), y efectos no genómicos, causados por la interacción con las membranas celulares, incluyendo membranas mitocondriales(23-27). Nuevos conocimientos se han adquirido en relación a las acciones genómicas clásicas (18); estos son los mecanismos más importantes de acción de los GC y dan lugar a cambios en la expresión de los genes. Estas acciones son causadas por el paso de moléculas de GC a través de la membrana plasmática con una alta afinidad de unión al cGCR inactivo, con formación y traslocación del complejo GC/cGCR dentro del núcleo. En la ‘transactivación' el complejo dimerizado GCR-proteína se une al promotor de los genes regulados por GC, conduciendo finalmente a la síntesis de ciertas proteínas reguladas por la activación transcripcional. La ‘transrepresión' monómerica de GCR interfiere con la actividad pro-inflamatoria de factores de transcripción tales como la proteína activadora 1 (AP-1) y el factor nuclear kB (NF-kB), lo que lleva a la baja regulación de la síntesis de proteínas pro inflamatorias. En contraste con los efectos no genómicos, que se producen rápidamente, en cuestión de segundos o minutos, algunos procesos genómicos requieren por lo menos de 30 minutos antes de que se puedan generar cambios significativos en las concentraciones de proteínas reguladoras. Por lo general, se tarda horas o días para que se produzcan estos cambios en células, tejido su órganos(12, 27, 28).
Hasta hace poco, se creía que los efectos represivos y antiinflamatorios de los GC dependían principalmente de ‘transrepresión', mientras que el desarrollo de las enfermedades autoinmunes y las acciones metabólicas de la terapia con GC dependían de la ‘transactivación' (18). Este punto de vista ha sido revisado recientemente, con la sugerencia de que las acciones antiinflamatorias claves de GCs son causadas por la activación de genes (18-20). Esta clase de genes incluye: la proteína fosfatasa dual específica 1 (DUSP1), la proteína quinasa activada por mitógenos fosfatasa1 (MAPK-1), el gen antiinflamatorio interleucina-10 (IL-10), la citoquina antiinflamatoria inducida por GC, con motivo estructural de cremallera de leucina (GILZ), una proteína de interacción de NFkB la cual inhibe la función de NFkB y AP-1, un fosfolípido-dependiente de calcio (Anexina-1). (12-13, 28). Las observaciones de los efectos inmunomoduladores de GILZ en la enfermedad autoinmune sobre linfocitos T y sinoviocitos en la artritis reumatoide (AR) en un modelo in vivo de AR, apoyan la hipótesis de que GILZ es un regulador clave de inflamación por GC en la enfermedad autoinmune (29).
Materiales y métodos
Se realizó un estudio analítico de casos y controles. La edad de los pacientes estuvo en el rango entre 18 a 65 años, en su mayoría de género femenino. Los criterios de inclusión de los casos fueron pacientes con diagnóstico de enfermedad autoinmune activa sin tratamiento (corticoides e inmunosupresores), utilizando como criterio el índice de MEX-SLEDAI (índice de activa lúpica) con un puntaje mayor a 3 y menor de 12 y en los controles fueron personas sin enfermedad autoinmune, con resultados serológicos negativos para este tipo de enfermedad.Anticuerpos anti DNA, anti-nucleares negativos, antígenos extractables negativo, RA test negativo, PCR normal, niveles séricos de complemento C1q, C2 y C4 normales, sin sintomatología asociada a la enfermedad; y como criterios de exclusión, sujetos que presentaran infecciones virales y bacterianas evaluadas clínicamente y pacientes con enfermedades infecciosas crónicas como HCV/HIV.
El tamaño de muestra fue de 26 pacientes con una relación de caso y control 1:1(13:13), para una diferencia mínimo de 0,10 de expresión de cadena Z, entre los grupos caso y control, con una confiabilidad del 95 % (error tipo I= 5%) y poder del 80% (error tipo II=20%).
Las variables de estudio fueron la presencia o no de enfermedad autoinmune, que fue la variable dependiente; y como independiente, la expresión de cadena Z medida con la técnica de RT-PCR y densitometría; y las variables de confusión fueron la edad y género del sujeto.
En todos los individuos evaluados (controles y pacientes), se realizó la amplificación de la β-actina, como control intrínseco y con base en ello se normalizaron los datos obtenidos con el fin de determinar el porcentaje relativo de expresión de la cadena zeta. Este proyecto de investigación fue un estudio observacional analítico sin ninguna intervención para el paciente y de acuerdo con lo establecido en la Resolución 008430 de 1993 de Colombia (Normas científicas, técnicas y administrativas para la investigación en salud),aunque se realizaron pruebas serológicas para diagnóstico de enfermedad autoinmune, siguiendo todas las normas de bioseguridad dada por la Secretaría de Salud de Bogotá, en los laboratorios de Bioquímica de la Universidad Nacional. El trabajo fue evaluado y aprobado por el comité de ética de la Sociedad Colombiana de Reumatología.
Toma demuestra
A cada paciente se le realizó una extracción de 10 ml de sangre total, se utilizaron tubos heparinizados, la muestra fue centrifugada a 3500 revoluciones por 5 minutos, el sobrenadante fue descartado y el precipitado (pellet) fue resuspendido adicionando 10 ml de solución salina. En un tubo falcón se adicionaron 3 ml de Ficoll-Hypaque (Ficoll Hypaque S.G. 1077; Amersham Pharmacia Biotech; Upsala, Sweden) lentamente por la pared del tubo. Después, las muestras fueron centrifugadas a 1200 revoluciones por 12 minutos, la capa de mononucleares fue separada y la interface de linfocitos extraída por succión.
Separación de linfocitos
Se obtuvieron células mononucleares de sangre periférica (PBMCs) de las cuales los linfocitos T fueron separados por depleción, utilizando un coctel de anticuerpos conjugados los cuales marcan células no-T y las retienen adhiriéndolas a perlas magnéticas (Dynal; Dynal Biotech; Lake Success, NY).El porcentaje de células obtenido por esta técnica fue del 98 %.
Extracción de RNA
La extracción de RNA se realizó adicionando 400 µl de cloroformo a cada muestra (células T obtenidas de sangre periférica y separadas por el método de ficoll), las muestras fueron mezcladas y agitadas (vortéx) durante 5 minutos, centrifugadas a 4 grados por 10 minutos a 12000 revoluciones, se les adicionó 1 ml de isopropanol, se dejaron a -20°C durante 20 min, posteriormente se centrifugaron a 12000 revoluciones durante 15 minutos a 5°C descartando el isopropanol y conservando el pellet.
El pellet se resuspendió con etanol, manteniéndose a una temperatura de -20°C toda la noche, las muestras se centrifugaron a 12000 revoluciones por 15 minutos a 4°C y se removió el etanol. El pellet fue resuspendido en 20 µls de agua DEPC y homogenizado. Las muestras fueron sometidas a choque térmico por cerca de 7 minutos, dando vortex y nuevamente colocadas en el termobloque a 60°C y guardadas a -20°C.
RT-PCR
El RNA total fue cuantificado, utilizándose50ng para obtención de cDNA(kit utilizado RT-PCR; promega), para la amplificación se utilizaron primers sintetizados por Sigma Genosys y se empleó un termociclador Smart.
Primers utilizados para la amplificación por RT-PCR
5'- TCTCACTGCCTTGTTCCTGA -3' and 5'- CGGTGCAGAGAACAGGTTT los cuales flanquean una región de 138 pares de bases, involucra exones 2, 3 y 4 del nucleótido 259 al 399 (figura 1).
Se realizó denaturación inicial a 94 °C por 6 min, 33 ciclos a 94 °C por 1 min; 67 °C por1 min; 72 °C por 2 min y una extensión final a 72 °C por7 min. Los productos de la PCR fueron separados utilizando un gel de agarosa al 1,5%.

Densitometría
Los productos de la RT-PCR fueron sometidos a electroforesis en geles de agarosa al 1,5 % y teñidos con bromuro de etidio. Los valores numéricos de la cantidad de amplificado fueron obtenidos sometiendo dichos geles a densitometría mediante el uso del equipo de análisis y foto documentación de geles Ref 05-CG9/ DZPC mediante un software (documentador digital). La figura 2 muestra los resultados obtenidos utilizando las células de los normales y los pacientes con enfermedad autoinmune.
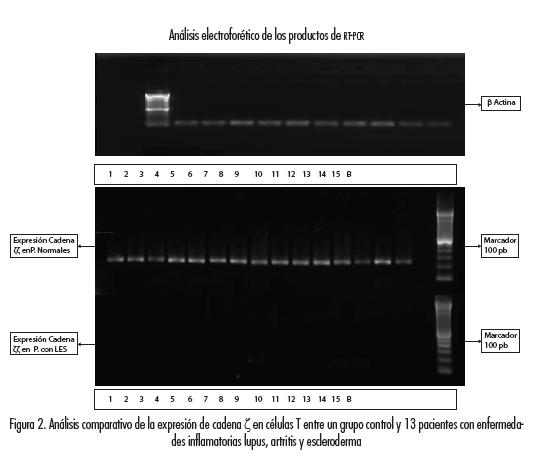
El análisis de los cambios relativos de la expresión génica para cadena zeta de los experimentos realizados mediante la técnica de PCR tiempo real se realizó utilizando el método 2-ΔΔCT, para el análisis cuantitativo en expresión de cadena zeta se utilizó como control interno β actina (gen constitutivo).
Análisis estadístico Se sistematizaron los datos del estudio en el paquete estadístico SPSS versión 20.0.Se evaluó la normalidad de las variables numéricas (densitometria y expresión de cadena Z) con la prueba de Shapiro Wilk y la homogeneidad de varianzas con el test de Levene. Se determinó la diferencia entre los promedios con la prueba T-student para grupos independientes o de medianas con la prueba no-paramétrica de Mann Whitney exacta. Para las comparaciones con los tipos de patología se usó un Anova paramétrico a una vía y posteriormente la prueba de comparaciones múltiples T2 de Thamane para varianzas heterogéneas, en caso de distribuciones diferentes a normal se tuvo en cuenta el Anova no-paramétrico exacta de Kruskall-Wallis. Las pruebas estadísticas se evaluaron a un nivel de significancia del 5 % (p<0,05).
Resultados
El grupo con la enfermedad (casos) lo conformaron 13 pacientes (6 con LES, 5 escleroderma y 2 con artritis de inicio tardío) y 13 sujetos como grupo control. El género más frecuente fue el femenino, en su distribución no mostró diferencias significativas entre los casos (69,2%, n=9)y controles (92,3 %, n=12) (p=0,322, test exacto de Fisher). El promedio de edad de los casos fue 48,62±12,47 años y en los controles 50,46 ±9,41años, sin mostrar diferencias significativas (p=0,674, T-student para varianzas homogéneas).
Expresión de cadena zeta
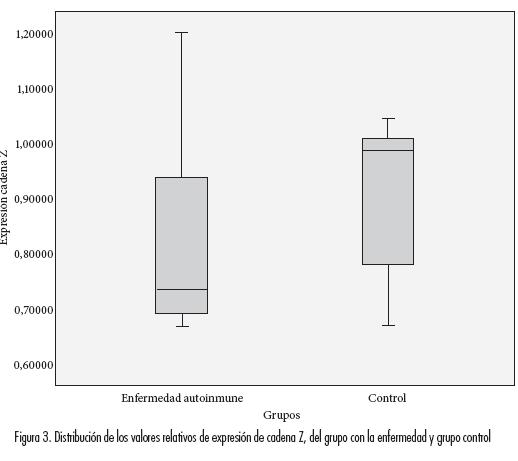
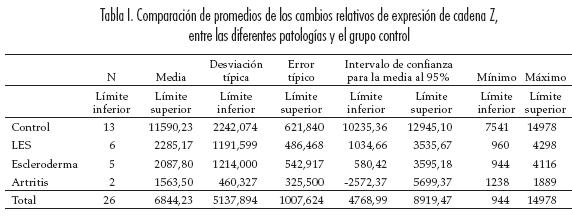
Utilizando la técnica de RT-PCR se evaluó semicuantitativamente la cantidad de mRNA de cadena zeta (ζ) presente en células T de pacientes con LES, muestras obtenidas de sangre periférica. Los valores relativos de expresión de cadena Z mostraron una distribución diferente a normal en los dos grupos, con asimetría a la derecha (p=0.006, test de Shapiro Wilk). Al comparar se encontraron diferencias significativas en las distribuciones, siendo menor la expresión en los pacientes con enfermedad autoinmune (0,8214±0,1787, med=0,7368) comparada con el grupo control (0,9225±0,12727, med=0,9830) (p=0,045, test no-paramétrico, exacto de Mann Whitney a una cola) (figura 3).
En la densitometría se encontró menor la expresión significativamente en los casos (2098,23±1079,69, med=1843,67) que en los controles (11590,23±2242,07, med=12308,67) (p<0,001, T-Student para varianzas heterogéneas) (figura 4).
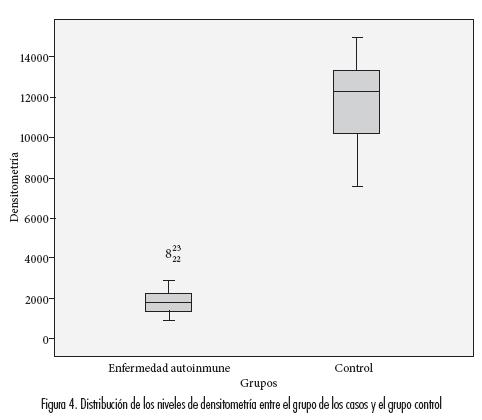
Al comparar con las patologías específicas de comparaciones múltiples de Tamanhe), con se encontraron diferencias significativas entre escleroderma (p<0, 001, test de comparaciones los promedios (p<0,001) y fueron mayores múltiples de Tamanhe) y con artritis (p<0,001, los valores de la densitometría al comparar test de comparaciones múltiples de Tamanhe) entre el grupo control con LES (p<0, 001, test (tabla 1).
Discusión
Aunque los mecanismos moleculares envueltos en la expresión defectuosa de ζTCR en pacientes con enfermedad autoinmune no son claros, una disminución o una falta de expresión de las cadenas pueden causar alteraciones en la respuesta inmune. La importancia en la evaluación de la expresión de zeta se debe a que un mal funcionamiento de esta proteína afecta la activación de factores de trascripción como NFκB, factor nuclear de activación del linfocito (NFAT), AP1, involucrados en la activación y síntesis de citoquinas por parte de los linfocitos T. Los defectos en zeta pueden alterar la respuesta conllevando a una sobre activación de linfocitos B con un aumento en la síntesis de anticuerpos, siendo uno de los eventos más importantes en el desarrollo de la enfermedad autoinmune (30).
Los estudios realizados por Tenbrock y Tsokos evidenciaron un cambio en la expresión de cadena zeta por el receptor Fcγ (FcγR) en células T de pacientes con LES, lo cual altera la vía de señalización, que termina con la expresión defectuosa de interleucina 2 (IL-2). Una alteración en la síntesis de IL-2 por parte de los linfocitos T, causa disminución en el número de células T efectoras y Treg en los pacientes. Los estudios de Tsokos han demostrado que la activación del linfocito T se incrementa considerablemente al presentarse la conmutación de estas proteínas (11, 30, 31). La explicación a dicho evento está asociada con el estímulo generado por la fosforilación del dominio intracitoplasmático de FcγR por la quinasa Syk en pacientes con LES, lo cual induce un incremento en la concentración de calcio (Ca2+) citoplasmático, lo cual activa la actividad fosfatasa de la calcineurina sobre el NFAT. Cuando el factor nuclear se transloca al núcleo, se induce la expresión de CD154 (CD40L) en el linfocito T, siendo en gran parte el responsable de la activación policlonal de células B y de la sobreproducción de anticuerpos por un mayor contacto entre linfocitos T y linfocitos B (11).
Los corticoides sintéticos utilizados en el tratamiento de enfermedades autoinmunes pueden alterar los eventos fisiológicos de la activación de células T, interfiriendo en los factores de trascripción que regulan principalmente la síntesis de citoquinas (32).La compleja función de señalización iniciada con la unión del antígeno al receptor de células T (TCR) es considerablemente dependiente de la expresión de la cadena Z TCR. La expresión anormal de esta proteína y la función defectuosa en los linfocitos T juegan un papel relevante en la patogenia de las enfermedades autoinmunes. Se ha sugerido que los defectos en su expresión contribuyen a la activación policlonal del linfocito B y al desarrollo de enfermedades autoinmunes como lupus, artritis, escleroderma, enfermedad de Graves y otros procesos patológicos (32,33). El dominio citoplasmático de la cadena Z TCR tiene tres motivos activadores del inmunorreceptor de tirosina (ITAM) (34,35).Tras la activación de TCR, dos residuos de tirosina dentro de cada ITAM se fosforilan, dando lugar al reclutamiento de moléculas de señalización corriente abajo. La fosforilación de varios residuos de tirosina en estas ITAM es crucial para el ensamblaje del complejo de señalización. Los eventos de señalización dirigen diversas funciones efectoras bajo diferentes condiciones de activación del receptor, cualquier modificación postrasduccional inducida por fármacos inmunosupresores o corticoides puede alterar la actividad fisiológica de los linfocitos (36-38).
El efecto de algunos corticoides sobre la expresión de la cadena Z TCR en células T estimuladas por antígenos induce alteración en la señalización, algunos defectos involucran modificaciones en los patrones de fosforilación de regiones intracitoplasmáticas de moléculas reguladoras, incrementando los niveles de calcio intracelular con activación de sustratos proteicos como el factor nuclear NFAT. Algunos estudios han demostrado que los corticoides regulan la baja la expresión de proteínas involucradas en la señalización de linfocitos T como la cadena Z de TCR (39), mientras que otros investigadores afirman que los corticoides sobre regulan la expresión de algunas isoformas de la cadena Z TCR con efectos inhibitorios en la síntesis de citoquinas (40,41). Trabajos recientes han demostrado que un gran número de glucocorticoides utilizados en el control de la enfermedad autoinmune son los responsables de la disminución de la trascripción génica (42-44). En virtud de los diversos puntos de vista citados anteriormente, sobre los efectos de los corticoides en la inhibición de la trascripción génica, se estudió la expresión de cadena zeta en pacientes con enfermedad autoinmune, que no recibieron corticoides, con el fin de determinar si los defectos en la expresión eran inducidos por la utilización de los mismos.
Los resultados obtenidos muestran una marcada disminución en la expresión de la cadena zeta en pacientes con diferentes enfermedades autoinmunes lo que nos permite inferir que no es un evento exclusivo, sino que la causa desencadenante podría ser un evento común en la enfermedad autoinmune.
¿Qué genera esos cambios en la expresión de la cadena zeta? Posiblemente, la presencia de la enfermedad autoinmune produzca factores que modifiquen su expresión.
En consonancia con los cambios en la expresión de la cadena zeta, los trabajos de Boldizsar en 2013 muestran un aumento en la expresión de la cadena Z de TCR, en pacientes tratados con bajas dosis de corticoides, con un aumento en la fosforilación de las proteínas tirosinas quinasas como ZAP-70 corriente abajo, potenciando además intermediarios de señalización como la proteína LAT en respuesta a una exposición antigénica (45, 46). La baja concentración de dexametasona (10 nM) aumenta la actividad transcripcional del promotor de TCR Z, el factor de trascripción elf-1en linfocitos T humanos (35). A una mayor dosis (100nM), tiene un efecto contrario suprimiendo su expresión, potenciando la actividad inhibidora del sistema inmune, inhibiendo la actividad de algunos factores de trascripción que interfieren con la expresión de algunas proteínas y modulando diferencialmente la expresión de cadena ζ en células T humanas estimuladas con antígenos.
Aún no son claros los mecanismos que sub-yacen estos complejos eventos regulatorios (47), el tratamiento con dexametasona y otros inmunosupresores.
En el presente trabajo se observó baja estabilidad del RNA ζTCR en células T de pacientes con enfermedad autoinmune: lupus, escleroderma y artritis de inicio tardío comparadas con los sujetos normales, los defectos en la expresión de zeta probablemente son dependientes de fallas en los mecanismos postranscripcionales como lo son los defectos en los factores de transcripción elf-1, ubiquitinación y proteólisis independientes del uso de corticoides (32). Los primers utilizados para este ensayo flanquean una región que involucra los exones 2,3 y 4 de ζ TCR y van del aminoácido 259 al 399, esto permitió evaluar la expresión de la cadena sin evaluar la actividad de ζ TCR (30).
Son muchas las preguntas que surgen sobre su regulación, sobre la modulación de los factores de trascripción que regulan su producción, así como las modificaciones postrascripcionales asociadas con defectos en la activación celular. Los defectos en expresión en la fosforilación están asociados con el desarrollo de enfermedades autoinmunes, sin embargo, aún no es claro si todas las subpoblaciones de linfocitos T de pacientes con enfermedad autoinmune presentan defectos en las proteínas involucradas en las vías de señalización o es exclusivo de una subpoblación de linfocitos T que regula la respuesta inmune.
Actualmente trabajamos en el análisis de expresión proteica de zeta y otras proteínas involucradas en señalización en subpoblaciones de células T reguladoras, así como en las modificaciones postrasduccionales asociadas al estadio de la enfermedad y a las dosis de corticoides utilizadas. Pretendemos hacer un análisis de la posible conmutación de ζ porFcγen pacientes con EA asociada al uso de corticoides e inmunosupresores.
Una mejor comprensión de la biología de zeta y de las proteínas adaptadoras involucradas en la señalización en células T de pacientes con enfermedad autoinmune contribuirá no solo a dilucidar los eventos de señalización intracelular en linfocitos T, sino también a generar herramientas terapéuticas que ayuden a controlar la enfermedad.
Bibliografía
1. Juang CM, Hung CF, Yeh JY, Horng HC, Twu NF, Cheng MH, et al. Regulatory T cells: potential target in anticancer immunotherapy. Taiwan J Obstet Gynecol. 2007; 46(3):215-21. [ Links ]
2. Akbari O, Stock P, De Kruyff R, Umetsu D. Role of regulatory T cells in allergy and asthma. Curr Opin Immunol. 2003;15(6):627-33. [ Links ]
3. Siachoque H, Rondón F, Restrepo JF, Pérez R, Iglesias A. Defectos en la síntesis de cadena ζ en el linfocito T, generando señales intracelulares equivocadas, responsables de lupus eritematoso sistémico. Rev Colomb Reumatol. 2010; 17(4):219-30. [ Links ]
4. Tsokos CG, Liossis SN. Immune cell signaling defects in lupus: activation, anergy and death. Immunol. Today. 1999; 20:119-24. [ Links ]
5. Jensen JP, Hou D, Ramsburg M, Taylor A, Dean M, Weissman AM. Organization of the human T cell receptor zeta/eta gene and its genetic linkage to the Fc gamma RII-Fc gamma RIII gene cluster. J Immunol. 1992; 148(8):2563-71. [ Links ]
6. Takeuchi T, Tsuzaka K, Pang M, Amano K, Koide J, Abe T. TCR zeta chain lacking exon 7 in two patients with systemic lupus erythematosus. Int Immunol.1998; 10(7):911-21. [ Links ]
7. Nambiar MP, Enyedy EJ, Fisher CU, Krishnan S, Warke VG, Guilliland WR, et al. Abnormal expression of various molecular forms and distribution of T cell receptor zeta chain in patients with systemic lupus erythematosus. Arthritis Rheum. 2002;46(1):163-74. [ Links ]
8. Vassilopoulos D, Kovacs B, Tsokos GC. TCR/CD3complex-mediated signal transduction pathway in T cells and T cell lines from patients with systemic lupus erythematosus. J Immunol.1995;155(4):2269-81. [ Links ]
9. Krishnan S, Nambiar MP, Warke VG, Fisher CU, Mitchell J, Delaney N, et al. Alterations in lipid raft composition and dynamics contribute to abnormal T cell responses in systemic lupus erythematosus. J Immunol. 2004;172(12):7821-31. [ Links ]
10. Spies CM, Strehl C, Van der Goes MC, Bijlsma J, Buttgereit F. Glucocorticoids. Best Practice & Research Clinical Rheumatology. 2011;25:891-900. [ Links ]
11. Hoffman GS. Immunosuppressive therapy for autoimmune diseases.Ann Allergy.1993;70(4):263-74. [ Links ]
12. Coutinho AE, Chapman KE. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol Cell Endocrinol. 2011;335(1):2-13. [ Links ]
13. Clark AR. Anti-inflammatory functions of glucocorticoid-induced genes. Mol Cell Endocrinol. 2007;275(1-2):79-97. [ Links ]
14. Alessandri AL, Sousa LP, Lucas CD, Rossi AG, Pinho V. Teixeira M. Resolution of inflammation: mechanisms and opportunity for drug development. Pharmacol Ther. 2013;139(2):189-212. [ Links ]
15. Nambiar MP, Enyedy EJ, Fisher CU, Warke VG, Juang Y, Tsokos GC. Dexamethasone modulates TCR zeta chain expression and antigen receptor-mediated early signaling events in human T lymphocytes. Cell Immunol. 2001;208(1):62-71. [ Links ]
16. Liberman AC, Druker J, Perone MJ, Arzt E. Glucocorticoids in the regulation of transcription factors that control cytokine synthesis. Cytokine Growth Factor Rev 2007;18(1-2):45-56. [ Links ]
17. Flammer JR, Rogatsky I. Minireview: Glucocorticoids in autoimmunity: unexpected targets and mechanisms. Mol Endocrinol. 2011;25(7):1075-86. [ Links ]
18. Fahey AJ, Robins RA, Kindle KB, Heery DM, Constantinescu CS. Effects of glucocorticoids on STAT4 activation in human T cells are stimulus-dependent. J Leukoc Biol. 2006;80(1):133-44. [ Links ]
19. Luther C, Adamopoulou E, Stoeckle C, Brucklacher-Waldert V, Rosenkranz D, Stoltze L, et al. Prednisolone treatment induces tolerogenic dendritic cells and a regulatory milieu in myasthenia gravis patients. J Immunol. 2009;183(2):841-8. [ Links ]
20. Zhou L, Ivanov II, Spolski R, Min R, Shenderov K, Egawa T, et al. IL-6 programs Th-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat Immunol. 2007;8(9):967-74. [ Links ]
21. Flammer JR, Dobrovolna J, Kennedy MA, Chinenov Y, Glass CK, Ivashkiv LB, Rogatsky I. Type I interferon signaling pathway is a target for glucocorticoid inhibition. Mol Cell Biol. 2010;30(19):4564-74. [ Links ]
22. Velten FW, Duperrier K, Bohlender J, Metharom P, Goerdt S. A gene signature of inhibitory MHC receptors identifies a BDCA3(+) subset of IL-10-induced dendritic cells with reduced allostimulatory capacity in vitro. Eur J Immunol. 2004;34(10):280011. [ Links ]
23. Buttgereit F, Straub RH, Wehling M, Burmester GR. Glucocorticoids in the treatment of rheumatic diseases: an update on the mechanisms of action. Arthritis Rheum. 2004;50(11):3408-17. [ Links ]
24. Buttgereit F, Wehling M, Burmester GR. A new hypothesis of modular glucocorticoid actions: steroid treatment of rheumatic diseases revisited. Arthritis Rheum. 1998;41:761-7. [ Links ]
25. Croxtall JD, Choudhury Q, Flower RJ. Glucocorticoids act within minutes to inhibit recruitment of signalling factors to activated EGF receptors through a receptor-dependent, transcription-independent mechanism. Br J Pharmacol. 2000;130(2):289-98. [ Links ]
26. Rhen T, Cidlowski JA. Anti-inflammatory action of glucocorticoids-new mechanisms for old drugs. N Engl J Med. 2005;353(16):1711-23. [ Links ]
27. Stahn C, Buttgereit F. Genomic and nongenomic effects of glucocorticoids. Nat Clin Pract Rheumatol. 2008;4(10):525-33. [ Links ]
28. Hoffman H, Rosengren S, Boyle DL, Cho JY, Nayar J, et al. Prevention of cold-associated acute inflammation in familial cold autoinflammatory syndrome by interleukin-1 receptor antagonist. Lancet. 2004;364:1779-1785. [ Links ]
29. Buttgereit F, Burmester GR, Straub RH, Seibel MJ, Zhou H. Exogenous and endogenous glucocorticoids in rheumatic diseases. Arthritis Rheum. 2011;63(1):1-9. [ Links ]
30. Sakai A, Han J, Cato AC, Akira S, Li JD. Glucocorticoids synergize with IL-1beta to induce TLR2 expression via MAP kinase phosphatase 1- dependent dual Inhibition of MAPK JNK and p38 in epithelial cells. BMC Mol Biol. 2004;5:2. [ Links ]
31. Beaulieu E, Morand EF. Role of GILZ in immune regulation, glucocorticoid actions and rheumatoid arthritis. Nat Rev Rheumatol. 2011;7(6):340-8. [ Links ]
32. Nambiar MP, Fisher CU,Warke VG, Krishnan S, Mitchell JP, Delaney N, et al. Reconstitution of deficient T cell receptor chain restores T cell signaling and augments T cell receptor/CD3-induced interleukin-2 production in patients with systemic lupus erythematosus. Arthritis Rheum. 2003;48(7):1948-55. [ Links ]
33. Tenbrock K, Juang T, Kyttaris VC, Tsokos GC. Altered signal transduction in SLE T cells. Rheumatology. 2007;46(10):1525-30. [ Links ]
34. Franchimont D, Galon J, Gadina M, Visconti R, Zhou Y, Aringer M, et al. Inhibition of Th1 immune response by glucocorticoids: dexamethasone selectively inhibits IL-12-induced Stat-4 phosphorylation in T lymphocytes. J Immunol. 2000;164:1768-74. [ Links ]
35. Li R, Yang W, Zhang J, Hirankarn N, Pan HF, Mok CC, et.al. Association of CD247 with systemic lupus erythematosus in Asian populations. Lupus. 2012;21(1):75-83. [ Links ]
36. Yoshimoto K, Setoyama Y, Tsuzaka K, Abe T, Takeuchi T. Reduced Expression of TCR zeta is involved in the abnormal production of cytokines by peripheral T cells of patients with systemic lupus erythematosus. J Biomed-Biotechnol. 2010;10:1-6. [ Links ]
37. Irving BA, Chan AC, Weiss A. Functional characterization of a signal transducing motif present in the T cell antigen receptor z chain. J Exp Med. 1993;177(4):1093-103. [ Links ]
38. Zenner G, Vorherr T, Mustelin T, Burn P. Differential and multiple binding of signal transducing molecules to the ITAMs of the TCR zeta chain. J. Cell Biochem. 1996;63(1):94-103. [ Links ]
39. Wange RL, Samelson LE. Complex complexes: signalingat the TCR. Immunity. 1996;5(3):197-205. [ Links ]
40. Kersh EN, Shaw AS, Allen PM. Fidelity of T cell activation through multistep T cell receptor zeta phosphorylation. Science. 1988;281(5376):572-5. [ Links ]
41. Kersh EN, Kersh GJ, Allen PM. Partially phosphorylated T cell receptor zeta molecules can inhibit T cell activation. J Exp Med. 1999;190(11):1627-36. [ Links ]
42. Migliorati G, Bartoli A, Nocentini G, Ronchetti R, Moraca R, Riccardi C. Effect of dexamethasone on T-cell receptor/CD3 expression. Mol Cell Biochem.1997;167(1-2):135-44. [ Links ]
43. Boumpas DT, Anastassiou ED, Older SA, Tsokos GC, Nelson DL, Balow JE. Dexamethasone inhibits human interleukin 2 but not interleukin 2 receptor gene expression invitro at the level of nuclear transcription. J. Clin. Invest. 1991;87(5):1739-47. [ Links ]
44. Barnes PJ, Adckock I. Anti-inflammatory actions of steroids: molecular mechanisms. Trends Pharmacol Sci. 1993;14(12):436-41. [ Links ]
45. Giunchi L, Nocentini G, Ronchetti S, Bartoli A, Riccardi C, Migliorati G. TCR kappa, a new splicing of the murine TCR zeta gene locus, is modulated by glucocorticoid treatment. Mol Cell Biochem. 1999;195(1-2):47-53. [ Links ]
46. Boldizsar F, Szabo M, Kvell K, Czompoly T, Talaber G, Bjorkan J, et al. ZAP-70 tyrosines 315 and 492 transmit non-genomic glucocorticoid (GC) effects in T cells. Mol Immunol. 2013;53(1-2):111-7. [ Links ]
47. Ronchetti S, Nocentini G, Giunchi L, Bartoli A, Moraca R, Riccardi C, Migliorati G. Short term dexamethasone treatment modulates the expression of the murine TCR zeta gene locus. Cell Immunol. 1997;178(2):124-31. [ Links ]
