










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista EIA
versão impressa ISSN 1794-1237
Rev.EIA.Esc.Ing.Antioq no.20 Envigado jul./dez. 2013
ASOCIACIÓN DE TRES SNPS CON ALGUNAS CARACTERÍSTICAS PRODUCTIVAS Y DE CALIDAD DE LA LECHE EN HATOS HOLSTEIN EN ANTIOQUIA, COLOMBIA
ASSOCIATION OF THREE SNPS WITH SOME PRODUCTION AND QUALITY CHARACTERISTICS OF HOLSTEIN CATTLE MILK IN ANTIOQUIA, COLOMBIA
ASOCIACAO DE TRÊS SNPs COM ALGUMAS CARATERÍSTICAS PRODUTIVAS E DE QUALIDADE DA LEITE EM MANADA HOLSTEIN EM ANTIOQUIA, COLÔMBIA
José Julián Echeverri Zuluaga*, Albeiro López Herrera**, Luis Ernesto López Rojas***
* Zootecnista, Ph. D. en Ciencias Agrarias. Profesor Asistente, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, sede de Medellín, Grupo Biodiversidad y Genética Molecular (BIOGEM). Medellín, Colombia.
Autor de correspondencia: (J.J. Echeverri-Zuluaga). Calle 59A no. 63 - 20, Medellín, Colombia. Teléfono: 440 90 00 ext. 904. Correo electrónico: jjecheve@unal.edu.co.
** Médico veterinario, PhD en Ciencias Básicas Biomédicas. Profesor Asociado, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, sede de Medellín, Grupo Biodiversidad y Genética Molecular (BIOGEM). Medellín, Colombia.
*** Biólogo, MSc en Biotecnología, MBA, candidato a PhD en Biotecnología, Universidad Nacional de Colombia, sede de Medellín. Jefe Programa de Biología, Universidad CES y Escuela de Ingeniería de Antioquia -EIA-, Grupo Biología CES-EIA. Medellín, Colombia.
Artículo recibido: 12-II-2013 / Aprobado: 04-XI-2013
Discusión abierta hasta diciembre de 2014
RESUMEN
Algunas características productivas y de la calidad de la leche están relacionadas con los polimorfismos de los genes kappa caseína (κ-CN), prolactina (PRL) y hormona de crecimiento bovina (bGH), estos han sido ampliamente estudiados y aplicados en programas de mejoramiento genético animal. De manera particular, la industria ganadera colombiana se ha cualificado de manera sustancial, gracias a la utilización de estas estrategias que permiten identificar las variantes alélicas relacionadas y la implementación de programas de selección asistida por marcadores moleculares. Con el fin de verificar la asociación de los SNP de estos genes con algunas características productivas y de la calidad de la leche en las ganaderías antioqueñas, se evaluaron 4.453 lactancias de 1.631 vacas pertenecientes a 69 hatos lecheros de diferentes municipios del departamento de Antioquia. En el análisis se incluyó la producción de leche ajustada (PA), el porcentaje de proteína (PRO) y grasa (GRA), el recuento de células somáticas (RSC) y el efecto de los genotipos de los marcadores antes mencionados. Los resultados corroboran la asociación de estos marcadores con las características evaluadas, y sugieren un efecto importante del genotipo de la κ-CN sobre el porcentaje de proteína (BB>AB>AA) (p<0,01), de bGH sobre el porcentaje de grasa (-/- > +/- > +/+) (p<0,01) y de PRL sobre la producción de leche (BB>AB>AA) (p=0,01).
PALABRAS CLAVE: bGH; grasa; κ-caseína; prolactina; proteína; RCS.
ABSTRACT
Some production and quality characteristics of milk are related to kappa casein (κ-CN), prolactin (PRL) and bovine growth hormone (bGH) gene polymorphisms, which have been widely studied and applied. In particular, the Colombian cattle industry has been substantially qualified, through the use of these strategies to identify the allelic variants related and implementation of breeding programs assisted by molecular markers. In order to verify the association of SNPs of these genes with some production traits and milk quality in herds of Antioquia, 4.453 lactations of 1.631 cows from 69 dairy herds in different municipalities of Antioquia was evaluated. The analysis included features such as milk production adjusted (PA), the protein (PRO) and fat (GRA) percentage, somatic cell score (SCS) and the effect of markers of the genotypes mentioned above. The results confirm the association of these markers with the evaluated characteristics, and suggest a significant effect of κ-CN genotype on protein content (BB>AB>AA) (p<0.0001), bGH on the percentage of fat (-/- > +/- > +/+) (p <0.0001) and PRL on milk production (BB>AB>AA) (p=0.0066).
KEY WORDS: bGH, fat, κ-casein, prolactin, protein, SCS.
SUMÁRIO
Algumas características produtivas e da qualidade da leita estão relacionado com os polimorfismos dos genes kappa caseína (κ-CN), prolactina (PRL) e hormônios de crescimento bovino (bGH), que foram estudados e aplicados em programas de melhoramento genético animal. De maneira particular, a indústria pecuária colombiana tornou-se qualificada de maneira substancial, graças à utilização destas estratégias que permitem identificar as variantes alélicas relacionadas e a implementação de programas de seleção assistidos por marcadores moleculares. Para verificar a associação dos SNP destes genes com algumas caraterísticas produtivas e da qualidade da leita nos lugares de criação de gado em Antioquia, avaliaram-se 4.453 lactâncias de 1.631 vacas pertencente a 69 manadas leiteiras de diferentes municípios do Departamento de Antioquia. Na análise, incluiu-se a produção de leite ajustado (PA), o percentagem de proteína (PRO) e gordura (GRA), o reconto de células somáticas (RSC) e o efeito dos genótipos dos marcadores mencionados. Os resultados corroboram a associação dos marcadores com as caraterísticas avaliadas, e sugerem um efeito importante do genótipo da κ-CN sobre o percentagem de proteína (BB>AB>AA) (p<0,01), de bGH sobre o percentagem de gordura (-/- > +/- > +/+) (p<0,01) e de PRL sobre a produção de leite (BB>AB>AA) (p=0,01).
PALAVRAS-CHAVE: bGH, gordura, κ-caseína, prolactina, proteína, RCS.
1. INTRODUCCIÓN
Desde la década del 90 se ha sugerido la importancia de estudiar y caracterizar los marcadores moleculares relacionados con la calidad de los productos lácteos. Lo anterior se fundamenta en la existencia de polimorfismos genéticos en los genes que codifican para las proteínas que participan en los procesos de producción y composición de la leche, como las de la familia de las caseínas, dado que la leche de los animales portadores del alelo B de la κ-CN, tiene un mayor rendimiento quesero, debido a una mayor cantidad y calidad de proteínas (Medrano, 1990).
De igual manera, se ha demostrado la importancia de algunas proteínas que participan en la regulación de la producción de lípidos y proteínas de la leche, como es el caso de la PRL (Le Provost, et al., 1994), o regulan su volumen, como la bGH (Hoj, et al., 1993), puesto que se ha documentado que ambas están involucradas en la activación de vías de señalización celular que inducen la expresión de κ-CN, β-lactoglobulina y de otras proteínas ácidas presentes en la leche (Hernández, 2001; Paul, et al., 2002). Hay evidencias que sugieren que los animales portadores del alelo B de la PRL producen un volumen mayor de leche, con un contenido de grasa superior (Alipanah, et al., 2007; Echeverri, et al., 2010b), y de manera similar, que el alelo (-) de bGH, está relacionado con mayor producción de leche y una concentración de proteínas superior (Echeverri, et al., 2010b), al parecer debido a una mayor cantidad circulante de la bGH (Loevendahl, 2004).
El hato especializado para leche en Colombia está compuesto en su mayor parte por individuos de la raza Holstein, debido a los altos volúmenes que produce. Sin embargo, en esta los alelos de los genes κ-CN (López y Vásquez, 2004; Trujillo, et al., 2000), PRL (Echeverri, et al., 2010a; Rincón, et al., 2012) y bGH (Echeverri, et al., 2010b) relacionados con una mayor cantidad de sólidos, los cuales son altamente valorados por la industria de productos lácteos, se encuentran en muy baja frecuencia. Esta circunstancia ha llamado la atención de los productores, quienes requieren desarrollar programas de mejoramiento y adoptar tecnologías que maximicen estas ventajas.
Si bien es necesario aumentar las frecuencias de los alelos relacionados con la calidad de la leche, es importante tener en cuenta algunos estudios que han evidenciado dificultades al tratar de cambiar estas proporciones, debido a que, al parecer, existen algunos efectos pleiotrópicos relacionados con la aparición de problemas de salud de la ubre, principalmente mastitis, al intentar aumentar la producción de leche y su contenido proteínico (Van Dorp, et al., 1999), logrando el efecto contrario, aumentando los costos y el riesgo de descarte de animales afectados (Rainard y Riollet, 2006).
Lo anterior propició la realización de estudios en las ganaderías nacionales, que permitieran tener un panorama de su estado y las posibilidades de intervención utilizando las herramientas de la biología molecular, aplicadas a la genética de mejoramiento animal. Teniendo en cuenta lo anterior, se determinó el efecto de los polimorfismos de los genes κ-CN, PRL y bGH sobre las características productivas y de calidad de la leche de bovino, en ganaderías del departamento de Antioquia.
2. METODOLOGÍA
Se evaluaron 4453 lactancias de 1631 vacas, pertenecientes a 69 hatos ubicados en 13 municipios del Valle de Aburrá (Bello, Envigado y Medellín), el Norte (Belmira, Don Matías, Entrerrios, Santa Rosa de Osos, San Pedro de los Milagros y Yarumal) y Oriente (El Retiro, La Ceja, La Unión y Rionegro) de Antioquia, incluidos en el programa de evaluación genética de toros lecheros en condiciones tropicales (Tabla 1). De cada lactancia se registró la identificación del animal, la del padre y la madre y su raza. Además se registró la producción de leche (litros por lactancia), número y duración de lactancia, número de servicios por concepción, el intervalo entre partos, porcentaje de proteína y grasa y el recuento de células somáticas en la leche. Finalmente, se genotipificaron los SNPs de los genes κ-CN (Medrano, et al., 1990), PRL (Brym, et al., 2004) y bGH (Dybus, 2002), mediante la evaluación del polimorfismo de los fragmentos de restricción (RFLP) de los productos amplificados utilizando la reacción en cadena de la polimerasa (PCR), PCR-RFLP.
Extracción de ADN: de cada individuo se colectaron 5 ml de sangre periférica en tubos al vacío con EDTA como anticoagulante y se almacenaron a 4°C. Para la extracción del ADN se utilizó el método de salting out descrito por Miller, et al. (1988) y los productos se diluyeron hasta alcanzar una concentración entre 30 ng/mL.
Amplificación por PCR: para cada marcador se realizó una reacción independiente, utilizando los cebadores sugeridos en la literatura para κ-CN (5`ATTTATGGCCATTCCACCAAAG3´, 5´ATTTCGCCTTCTCTGTAACAGA3´) (Medrano, et al., 1990), PRL (5`CGAGTCCTTATGAGCTTGATTCTT3´, 5´GCCTTCCAGAAGTCGTTTGTTTTC3´) (Brym, et al., 2004), y bGH (5`CCCACGGGCAAGAATGAGGC3´, 5´TGAGGAACTGCAGGGGCCCA3´) (Dybus, 2002). A la mezcla de reacción se adicionaron 0,4 mM de cada dNTP, 0.2 µM de cebador, 2,5 µl de tampón PCR 10X libre de magnesio, 50 mM MgCl2, 1U de Taq ADN polimerasa (Fermentas, California, U.S.A) y 150 ng de ADN genómico, diluidos en agua ultra pura en un volumen final de 25 µL, la reacción se realizó en un termociclador (Biometra, Göttingen Germany), así: cinco minutos a 94°C para la desnaturalización inicial del ADN, seguida de 10 ciclos desnaturalización a 94°C durante 1 minuto, 40 segundos en la temperatura de alineamiento, un minuto a 72°C para la extensión y tres minutos a 72°C para la extensión final.
Genotipificación mediante RFLP: la digestión de cada amplificado se realizó utilizando 0,5 µL de endonucleaza HinfI, RsaI, MspI 10 U/mL (Fermentas®) para los amplificados de κ-CN, PRL y bGH, respectivamente, 2 µL de solución tampón (R, B o tango) y 5 µL de amplificado, en un volumen final de 20 µL, incubando a 37°C durante 2 a 8 horas, de acuerdo a las instrucciones del fabricante. Los fragmentos digeridos fueron resueltos utilizando como patrón un marcador de peso molecular de 50pb (Fermentas®), en una electroforesis en gel de agarosa al 2 %, teñidos con 10 µg/ml bromuro de etídio, durante 3 horas a 100 voltios y luego visualizadas bajo luz ultravioleta. Para la identificación de los alelos de los genes (Tabla 1) se tuvo en cuenta el patrón de bandas esperado para cada genotipo para κ-CN (Medrano, et al., 1990), PRL (Brym, et al., 2004), y bGH (Dybus, 2002).
Estimación de las frecuencias alélicas y genotípicas: la frecuencia de cada alelo se estimó determinando la proporción de cada forma del gen entre el número de copias totales de la población en estudio, identificando los homocigotos y los heterocigotos. El cálculo se realizó cuantificando los homocigotos para cada alelo y adicionando la mitad de los heterocigotos. Además, se verificó el equilibrio de Hardy-Weinberg (H-W), comparando las frecuencias observadas y esperadas de los alelos de cada marcador mediante la prueba de Pearson y la prueba exacta de Fisher en SAS 9.1 (Tabla 2).
Asociación de los marcadores moleculares con las características productivas de la leche: se realizó un análisis de ligamiento para determinar el grado de asociación entre los marcadores (κ-CN, PRL y bGH) y las características productivas de la leche (porcentaje de grasa y proteína y recuento de células somáticas) mediante un modelo estadístico utilizando el procedimiento GLM de SAS 9.1, que incluye como efectos fijos a los marcadores y sus genotipos (niveles), el número de lactancia, la región, el hato, el año y el mes, y el efecto de algunas covariables como la duración de la lactancia y la producción ajustada de leche. Se determinó la significancia de cada efecto, donde la significancia del efecto del genotipo indica una asociación entre éste y la característica evaluada. Así:
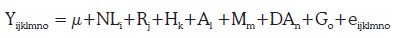
Donde, Yijklmno es la variable dependiente (Producción de leche por lactancia, Porcentaje de proteína de la leche, Porcentaje de grasa de la leche y puntaje de células somáticas); µ, la media general para todas las observaciones; NL, el efecto fijo del número de lactanciai; R efecto fijo de la regiónj; H, Efecto fijo del hatok; A, efecto fijo del añol; M, efecto fijo del mesm; DA, efecto de la covariable duración de la lactancia ajustadan; G, efecto fijo del genotipo para cada uno de los marcadores (bGH, PRL y κ-CN)o y e, el error aleatorio.
De igual manera se realizó un análisis de regresión entre cada una de las características dependientes y los genotipos, que fueron transformados como variables cuantitativas para estimar el efecto aditivo de cada uno de los alelos y determinar el efecto de la sustitución de un alelo por el recíproco de acuerdo con la característica dependiente, así:
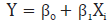
Donde, Y: fue la variable dependiente (producción de leche por lactancia, porcentaje de grasa, porcentaje de proteína o puntaje de células somáticas); β0, el intercepto; β1, el coeficiente de regresión estimado para el efecto del genotipo, y Xi, el genotipo del individuo (0, 1 ó 2). El coeficiente de regresión estimado es el efecto aditivo de cada uno de los alelos.
3. RESULTADOS
Frecuencias alélicas y genotípicas: en la Tabla 1 se muestran los lugares de muestreo y el número de individuos de cada genotipo de los genes evaluados, y en la Tabla 2 se muestran las frecuencias alélicas y genotípicas de cada uno de los marcadores, discriminadas por región y municipio. Se observa una baja frecuencia de los genotipos BB de κ-CN (0,061) y PRL (0,018) y -/- de bGH (0,014), en contraste con una alta frecuencia de los alelos A de la κ-CN (0,76) y la PRL (0,87) y + de la bGH (0,88); sin embargo la distribución de los alelos se encontró en equilibrio de H-W en la población general, así: κ-CN (χ2 p= 0,89), PRL (χ2 p=1) y bGH (χ2 p=0,96), y en las regiones Norte: κ-CN (χ2 p= 0,90), PRL (χ2 p= 0,99) y bGH (χ2 p=0,81); Oriente: κ-CN (χ2 p= 0,13), PRL (χ2 p= 0,24) y bGH (χ2 p= 1) y Valle de Aburrá: κ-CN (χ2 p= 0,30), PRL (χ2 p= 0,84) y bGH (χ2 p=0,85), es importante resaltar que no se encontró equilibrio de H-W en κ-CN (χ2 p=0,03) en el municipio de Belmira y la PRL (χ2 p= 0,01) en Envigado. Lo anterior sugiere que se conserva el carácter polimórfico de los loci evaluados, dado que en todos los casos el alelo más común se encontró en una frecuencia inferior a 0,95.
Análisis descriptivo: las lactancias evaluadas provienen de vacas genotipificadas para los genes κ-CN, PRL y bGH a las que se le registraron las características de producción de leche. Con el fin de balancear el modelo se agruparon los animales que tenían siete o más lactancias, de igual manera, se realizó un análisis para detectar datos extremos y, como consecuencia se descartaron los valores de producción de leche ajustada por lactancia superiores a 9.000 L e inferiores a 3.000 L, niveles de proteína inferiores a 2,5 % y superiores a 4% y niveles de grasa menores a 2 % y mayores a 4,5 %.
Se evidenció alta variabilidad en la producción ajustada de leche por lactancia, con una media de 5.595 ± 2.334 L, con CV de 41,71 %. El mayor volumen promedio se encontró en los municipios del Oriente Antioqueño (5.902,01±1.372,99 L), en segundo lugar en el Valle de Aburrá (5.472,83 ±1297,97 L), mientras que en las ganaderías del Norte se encontraron los volúmenes más bajos (5.448,64±1.400,64 L) (p<0,01) (Tabla 3).
La proteína presentó valores relativamente bajos, 3,05±0,28 %, CV de 7,45 %, se encontró diferencia estadística altamente significativa entre las regiones evaluadas (p<0,01), el Valle de Aburrá presentó el promedio más bajo (2,992±0,245 %), el Norte de Antioquia (3,016 ± 0,232 %), y el porcentaje más alto se encontró en las ganaderías lecheras del Oriente antioquieño (3,085 ±0,256 %) (Tabla 3).
El porcentaje promedio de grasa fue de 3,86±0,53%, CV 11,9, se encontró diferencia significativa entre las regiones evaluadas (p<0,05), el Valle de Aburrá (3,855±0,445 %), el Norte (3,811 ± 0,493 %), y el Oriente (3,913 ±0,548 %). Un comportamiento similar se observa en el RCS, cuyo valor promedio fue de 495±591, y su coeficiente de variación (CV) de 122,21 %; se encontró diferencia significativa entre las regiones evaluadas (p<0,05), en el Valle de Aburrá (336,97±304,23), el Norte de Antioquia (475,01±649), y el Oriente antioqueño (562,41 ±766,52) (Tabla 3).
Lo anterior podría estar relacionado con las bajas frecuencias alélicas y genotípicas de los marcadores asociados con esas características, se encontró una mayor proporción de individuos portadores de los alelos A de la κ-CN. Algo similar se encontró para el alelo A de la PRL, relacionado con una menor producción de proteína y grasa y el alelo (+) de la bHG variantes κ-CN, PRL y bGH (Tabla 2).
Análisis de ligamiento: al analizar los resultados del análisis del procedimiento GLM del SAS, se encuentra un efecto significativo de la región, el hato, el número de lactancia y duración de la lactancia sobre las variables producción ajustada de leche, el porcentaje de proteína y grasa y el recuento de células somáticas, (p<0,05), no obstante, no se observó efecto significativo sobre el RCS (p>0,05). Es importante resaltar que el genotipo para la κ-CN afectó el porcentaje de proteína en la leche (p<0,01), el genotipo para PRL la producción ajustada de leche (p<0,01), y el genotipo para bGH el porcentaje de grasa (p<0,01). El recuento de células somáticas no fue afectado por ninguno de los marcadores (p> 0,05) (Tabla 4).
Lo anterior se verificó realizando un análisis de comparación de medias (LSD) para comparar los diferentes niveles de los efectos significativos. En este caso, se encontró que la producción ajustada de leche fue mayor en los individuos con genotipos AA y AB de la κ-CN (p<0,05), mientras que el genotipo de la PRL fue BB>AB>AA (p<0,05) y la bGH no presentó ningún efecto. Para el caso del porcentaje de proteína se encontró un mayor valor promedio en los individuos BB>AB>AA del gen κ-CN, la media del porcentaje de grasa estuvo afectada por el genotipo de la bGH (-/- > +/- > +/+), mientras que en el RCS fue mayor en los individuos AA>AB>BB (p<0,05) (Tabla 5).
Los valores de r2 para el modelo de producción de leche (0,34), el porcentaje de proteína en la leche (0,28), el porcentaje de grasa en la leche (0,22) y el recuento de células somáticas (0,18), sugieren que estas variables son explicadas en un 34, 28, 22 y 18 % por sus respectivos modelos (Tabla 4). Además, que la variación restante se debe atribuir a otros efectos que no se incluyeron en esta investigación.
El coeficiente de regresión β para el porcentaje de grasa en la leche sugiere una disminución de 0,07 % a medida que aumenta el número de alelos (-) del gen bGH. En el caso de los genes κ-CN y PRL, el efecto del genotipo no explica de forma significativa (P>0,05) la variación en el porcentaje de grasa (Tabla 6). Por su parte, el porcentaje de proteína en la leche aumentó 0,04 % por cada alelo B del gen de la κ-CN, mientras que los efectos de los genotipos de PRL y bGH no tuvieron significancia estadística (P<0,05).
La producción de leche ajustada no mostró ninguna correlación con los alelos de los genes κ-CN, PRL y bGH, los cuales presentaron valores de p>0,05, no obstante se encontró una tendencia con el gen κ-CN, que disminuye la producción de leche en 139 L a medida que aumenta el número de alelos B. Para el caso del puntaje de células somáticas en la leche, no se encontró correlación con los genotipos de los genes κ-CN, PRL y bGH, con valores p>0,05.
4. CONCLUSIONES Y DISCUSIÓN
Los volúmenes de leche encontrados en las diferentes regiones de Antioquia son similares a los encontrados en trabajos previos (Echeverri, et al., 2011; Rodríguez, et al., 2013; Echeverry et al., 2010a; Echeverry, et al., 2010b) pero inferiores a la producción reportada por diversos investigadores en otros lugares del mundo: 6928 L (Dematawewa y Berger, 1998), 8258 L (Van Tassell, et al.,1999), 8687 L (Washburn, et al., 2001), 8662 L (West y Hill, 1990), 9076 L (Welper y Freeman, 1991), 9760 L (Rodríguez, et al.,1996), y 9912 L (Bitman, et al.,1996). Un comportamiento similar presentó el porcentaje de proteína y de grasa, coherente con los hallazgos reportados para algunas ganaderías del departamento de Antioquia (Ceballos, et al., 2012; Echeverry, et al., 2010a; Echeverry, et al., 2010b).
Con respecto a las frecuencias alélicas y genotípicas halladas se puede indicar que los reportes encontrados en Colombia para los alelos de bGH y PRL, en trabajos realizados por Echeverry, et al. 2010b, son coherentes con los hallados en esta investigación. Algunas frecuencias para κ-CN reportadas en vacas Holstein en Nariño por Solarte, et al. (2009), también fueron similares a las halladas en las poblaciones evaluadas en este trabajo.
En las poblaciones de ganado Holstein evaluadas, se percibe una baja frecuencia de los alelos favorables relacionados con las características de productividad de la leche, lo anterior confirma lo sugerido por la literatura (López y Vásquez, 2004). Esta situación impacta los programas de mejora genética, debido a que reduce la posibilidad de que estos se incrementen en la población. Solo los programas de selección asistida por marcadores moleculares objeto de esta investigación, permitirán que a futuro las frecuencias alélicas y genotípicas se desbalanceen a favor de los genotipos favorables, logrando mejoras en términos fenotípicos. Se ha encontrado que el porcentaje de grasa está relacionado con el genotipo de la bGH (p<0,0001), la producción de leche con el genotipo de la PRL (p=0,066) y el porcentaje de proteína con el genotipo de la κ-CN (p<0,0001), y que la variación en el recuento de células somáticas no es afectado por el genotipo de estos genes.
Lo anterior contrasta con algunos reportes que sugieren que el alelo B de la PRL regula tanto la producción de leche, como su contenido de lípidos y proteínas (Alipanah, et al., 2007; Echeverri, et al., 2010b; Le Provost, et al., 1994), de igual manera se sugiere que el alelo (-) de la bGH está relacionado con una concentración mayor de bGH circulante (Loevendahl, 2004), la cual afecta el volumen de leche (Hoj, et al., 1993; Echeverri, et al., 2010b) y contenido de proteínas debido a su influencia en la regulación de la expresión de κ-CN y la β-lactoglobulina (Hernández, 2001; Paul, et al., 2002).
Por tanto, sugiere un efecto pleiotrópico de los diferentes genes que podría dificultar los procesos de selección asistida por marcadores, toda vez que al aumentar las proporciones de un alelo particular para favorecer una característica, se estaría impactando de manera negativa en otras características relevantes en la eficiencia de la ganadería. De manera puntual, los alelos B de la κ-CN que aumentan el porcentaje de proteína (p<0, 01) muestran una tendencia a disminuir el volumen de leche (p=0,0561). Resultados similares fueron reportados para el gen de la κ-CN, en una investigación llevada a cabo en el departamento de Nariño por Zambrano, et al. (2010), quienes determinaron que el rendimiento quesero y la formación de la cuajada son superiores en vacas con genotipo BB.
Es claro que la producción de leche, su calidad sanitaria y nutritiva va en aumento, pero aún no se alcanzan los volúmenes reportados en las ganaderías europeas, los porcentajes de grasa y proteínas son muy modestos, mientras que el RCS aún es muy alto. Lo anterior trae como consecuencia bajos rendimientos de industrialización para la producción de quesos y otros derivados lácteos, y dificultades de comercialización.
Finalmente, aunque en los últimos años se han logrado grandes adelantos en la ganadería de leche en Colombia, es necesario realizar estudios minuciosos, que permitan optimizar los procesos productivos, obteniendo productos de calidad adecuada, sin detrimento de las poblaciones ganaderas.
REFERENCIAS
Alipnanah, M., Kalashnikova, L. and Rodionov, G. (2007). Asociation of Prolactin Gene Variants with Milk Production Tratits in Russian Red Pied catlle. Iranian Journal of Biotechnology 5(3) July, pp. 158-161. [ Links ]
Brym, P. (2004). Identification of Polymorphism within Bovine PRL, PRLR and STAT5A genes. Ph.D. Thesis. University of Warmia and Mazury. [ Links ]
Dematawewa, C. and Berger, P. (1998). Genetic and Phenotypic Parameters for 305-Day Yield, Fertility, and Survival in Holsteins. Journal of Dairy Science, 81(10) October, pp. 2700-2709. [ Links ]
Dybus, A. (2002). Associations of growth (GH) and Prolactin (PRL) Genes Polymorphisms with Milk Production Traits in Polish Black and White cattle. Animal Science Papers and Reports. 20(4), pp. 203-212. [ Links ]
Echeverri-Zuluaga, J.J., Vásquez-Araque, N.A. y Gallo-García, Y.M. (2010a). Efecto de la transición adenina/guanina del gen de la prolactina bovina sobre características de importancia en producción lechera. Revista Lasallista de Investigación, 7(2), pp. 16-23. [ Links ]
Echeverri-Zuluaga, J.J., Vásquez-Araque, N.A. y Gallo-García, Y.M. (2010b). Polimorfismo del gen de la Somatotropina Bovina y su asociación con características de importancia en la producción lechera. Revista Lasallista de Investigación, 7(1), pp. 58-65. [ Links ]
Echeverri, J., Salazar, V. y Parra, J. (2011). Análisis comparativo de los grupos genéticos Holstein, Jersey y algunos de sus cruces en un hato lechero del Norte de Antioquia en Colombia. Zootecnia Tropical. 29(1), pp. 49-59. [ Links ]
Hernández, D. (2001). Utilización de información molecular en programas de mejora genética. Revista Frisona Española. 125 septiembre-octubre, pp. 12-19. [ Links ]
Hoj, S., Fredholm, M., Karsen, N. y Nielsen, V. (1993). Growth Hormone Gene Polymorphism Associated with Selection for Milk Fat Production in Lines of cattle. Animal Genetics, 24, pp. 91-96. [ Links ]
Le Provost, E., Leroux, C., Martin, P., Gafe, P. and Dijane, J. (1994). Prolactin Gene Expression in Ovine and Caprine Mammary Gland. Neuroendocrinology, 60(3), pp. 305-313. [ Links ]
López, E., Vásquez-Araque y N.A. (2004). Determinación del sexo y genotipificación del gen de la  -caseína en embriones bovinos. Revista Colombiana Ciencias Pecuarias, 17(3) julio-septiembre, pp. 231-240. [ Links ]
-caseína en embriones bovinos. Revista Colombiana Ciencias Pecuarias, 17(3) julio-septiembre, pp. 231-240. [ Links ]
Loevendahl, P. (2004). Polymorphism of the Somatotropic Axis Genes in cattle - Physiology and Productivity. Animal Science Papers and Reports, 22(1), pp. 101-108. [ Links ]
Medrano, J.F. and Aguilar-Córdova, E. (1990). Genotyping of Bovine Kappa-Casein Loci Following DNA Sequence Amplification. Biotechnology, 8, pp. 144-146. [ Links ]
Miller, SA., Dykes, DD. and Polesky, HF. (1988). A Simple Salting out Procedure for Extracting ADN from Human Nucleated Cells. Nucleic Acids Research; 16(3), pp. 1215. [ Links ]
Paul, K., Bachelot, A., Kedzia, C., Hennighausen, L., Ormanndy, C., Kopchick, J. and Binart, N. (2002). The Role of Prolactin and Growth Hormone in Mammary Gland Development. Molecular and Cellular Endocrinology, 197, pp. 127-131. [ Links ]
Rainard, P. and Riollet, C. (2006). Innate Immunity of the Bovine Mammary Gland - Review Article. Veterinary Research, 37(3) May-June, pp. 369-400. [ Links ]
Rincón, JC., López, A. y Echeverri, J. (2012). Variabilidad genética del loci Rsal de la prolactina bovina en ganado Holstein de Antioquia, Revista Colombiana de Ciencias Pecuarias, 25(2), pp. 191-201. [ Links ]
Rodríguez, LA., Stallings, CC., Herbein, JH. and Mcgilliard, ML. (1997). Effect of Degradability of Dietary Protein and Fat on Ruminal, Blood, and Milk Components of Jersey and Holstein Cows. Journal Dairy Science, 80(2) February, pp. 353-363. [ Links ]
Rodríguez-Neira, J.D., Correa-Londoño, G.A. y Echeverri- Zuluaga, J.J. (2013). Prediction Models for Total Milk Yield and Fat Percentage Using Partial Samples. Revista Facultad Nacional Agronomía Medellín, 66(1), pp. 6909-6917. [ Links ]
Solarte-Portilla, CE., Rosero, CY., Eraso, JM., Zambrano, GL., Cárdenas, H. y Burgos, WO. (2009). Frecuencias alélicas del gen Kappa caseína en la raza Holstein del trópico alto de Nariño - Colombia. Livestock Research for Rural Development 21(1) en: http://www.lrrd.org/lrrd21/1/sola21003.htm. [ Links ]
Trujillo-Bravo, E.R., Camargo-Guerrero, M. y Noriega, D. (2000). Genotipificación de kappa-caseína bovina y evaluación de las frecuencias fenotípicas y alélicas de sus polimorfismos en cuatro razas. Actualidades Biológicas, 22, pp. 145-152. [ Links ]
Van Dorp, R.T. Martin, S.W., Shoukri, M.M., Noordhuizen, J.P. and Dekkers, JC. (1999). Cananadian. An Epidemiologic Study of Disease in 32 Registered Holstein Dairy Herds in British Columbia. Journal Veterinary Research, 63, pp. 185-192. [ Links ]
Washburn, S.P., Silvia, W.J., Brown, C.H., Mcdaniell, B.T. and McAllister, AJ. (2001). Trends in Reproductive Performance in Southeastern Holstein and Jersey DHI Herds. Journal Dairy Science, 85(1) January, pp. 244-251. [ Links ]
Welper, R.D. and Freeman, A.E. (1991). Genetic Parameters for Yield Traits of Holsteins, Including Lactose and Somatic Cell Score. Journal Dairy Science, 75(5) May, pp. 1342-1348. [ Links ]
West, J.W. and Hill, G.M. (1990). Effect of a Protected Fat Product on Productivity of Lactating Holstein and Jersey Cows. Journal Dairy Science, 73, pp. 3200-3207. [ Links ]
Zambrano-Burbano, G.L., Eraso-Cabrera, J.M., Solarte-Portilla, C.E. and Rosero-Galindo, C.Y. (2010). Kappa casein Genotypes and Curd Yield in Holstein Cows. Revista Colombiana de Ciencias Pecuarias, 23(4), pp. 422-428. [ Links ]
