










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista EIA
Print version ISSN 1794-1237
Rev.EIA.Esc.Ing.Antioq no.21 Envigado Jan./June 2014
EVALUACIÓN DE REQUERIMIENTOS NUTRICIONALES EN VIVERO DE ESPECIES TROPICALES EMPLEADAS EN SILVICULTURA URBANA
EVALUATION OF NUTRITIONAL REQUIREMENTS IN NURSERIES FOR CERTAIN SPECIES USED IN URBAN FORESTRY
AVALIAÇÃO DE REQUERIMENTOS NUTRICIONAIS EM VIVEIROS DE ESPÉCIES TROPICAL EMPREGADAS EM SILVICULTURA URBANA
David Andrés Herrera Ramírez*, Juan Diego León Peláez**, Mónica Ruiz Rendón***, Nelson Walter Osorio Vega****, Guillermo Correa Londoño*****, Ricardo Esteban Ricardo******, Ángela Uribe Bravo*******
*Ingeniero Forestal, Universidad Nacional de Colombia, sede Medellín. MSc.(c) Bosques y Conservación Ambiental, Universidad Nacional de Colombia, sede Medellín, (Colombia).
Autor de correspondencia Herrera-Ramírez, D.A. (David Andrés): Universidad Nacional de Colombia sede Medellín, Facultad de Ciencias Agropecuarias, Departamento de Ciencias Forestales, Medellín, Colombia. Tel: (574) 482 66 40. Correo electrónico: daherrerr@unal.edu.co.
**Ingeniero Forestal, Universidad Nacional de Colombia, sede Medellín. Especialista en Gestión Ambiental, Universidad de Antioquia (Colombia). MSc. en Evaluación del Impacto Ambiental, Instituto de Investigaciones Ecológicas, Málaga (España). PhD. en Manejo y Conservación de Recursos Naturales: Aplicación a la Gestión Ambiental, Universidad de Salamanca, Salamanca (España). Profesor Asociado, Universidad Nacional de Colombia, sede Medellin, Facultad de Ciencias Agrarias.
***Ingeniera Forestal, Universidad Nacional de Colombia, sede Medellín (Colombia).
****Ingeniero Agronómo, Universidad Nacional de Colombia, sede Medellín. MSc. y PhD. Agronomy and Soil Science, University of Hawaii. Profesor Asociado, Facultad de Ciencias Universidad Nacional de Colombia, Sede Medellín.
*****Ingeniero Forestal, Universidad Nacional de Colombia, sede Medellín. MSc. en Ciencias especialidad en Estadística, Colegio de Postgraduados, Texcoco (México). PhD. Estadística Multivariante Aplicada, Universidad de Salamanca, Salamanca (España). Profesor Asociado, Universidad Nacional de Colombia, sede Medellín, Facultad de Ciencias Agrarias.
******Ingeniero Forestal, Universidad Nacional de Colombia, sede Medellín. M.Sc. Medio Ambiente y Desarrollo, Universidad Nacional de Colombia, Sede Medellín (Colombia).
*******Ingeniera Forestal, Universidad Nacional de Colombia, sede Medellín. Especialista en Finanzas y Formulación de Proyectos, Universidad de Antioquia (Colombia).
Artículo recibido: 26-XI-2013 / Aprobado: 14-I-2014
Disponible online: 12 de mayo 2014
Discusión abierta hasta mayo de 2015
RESUMEN
La fertilización es determinante del crecimiento y desarrollo de árboles urbanos. Por ello, estudiamos, mediante la técnica del elemento faltante, los requerimientos nutricionales de cinco especies tropicales usadas en silvicultura urbana (Tabebuia chrysantha, Margaritaria nobilis, Hamelia patens, Apeiba aspera, Cupania americana). Se encontró algunas especies sensibles a la deficiencia de nitrógeno y otras a la deficiencia de azufre. El grupo de T. chrysanta, M. nobilis y H. patens mostró alta sensibilidad a la deficiencia de nitrógeno, mientras que C. americana y A. aspera presentaron sensibilidad a la deficiencia de azufre, representada en menor crecimiento promedio para todas las variables evaluadas: altura, diámetro en el cuello de la raíz, materia seca radical y materia seca aérea. Las representaciones HJ-Biplot, utilizadas para caracterizar la respuesta por especie a cada tratamiento en todas las unidades experimentales, evidenciaron los bajos rendimientos en crecimiento para las deficiencias en nitrógeno y azufre. El desbalance de bases calcio/magnesio y magnesio/potasio afectó el crecimiento en altura de todas las especies, evidenciado en los altos crecimientos obtenidos para los tratamientos deficientes en calcio y magnesio. Debido a esto, las fórmulas nutricionales utilizadas en vivero deben reajustarse a las exigencias individuales de las especies, para producir mejores árboles para la ciudad.
PALABRAS CLAVE: silvicultura urbana; especies tropicales; fertilización; nutrición forestal.
ABSTRACT
One of the determining factors in urban tree development and growth is fertilization. We therefore studied the nutritional requirements of five tree species used in urban forestry (Tabebuia chrysantha, Margaritaria nobilis, Hamelia patens, Apeiba aspera, Cupania americana) using the missing element technique. Differential responses by groups were found for nitrogen and sulfur deficiencies. The T. chrysanta, M. nobilis, and H. patens groups showed high sensitivity to nitrogen deficiencies. This was evident in a lower mean growth for all variables assessed: height, root neck diameter, aerial dry matter, and radical dry matter. The C. americana and A. aspera group showed the lowest mean growth for sulfur deficiencies. The HJ-Biplot representations, used to characterize the species' response to each treatment, showed low growth yields for nitrogen and sulfur deficiencies in every individual tree species assessed. The imbalance of calcium/magnesium bases and magnesium/potassium bases affected height growth in all species. This was evident in the high growth obtained for the calcium- and magnesium-deficient treatments. The treatments that added calcium and/or magnesium accentuated the imbalance and gave low growth in all variables.
KEY WORDS: Urban Forestry; Tropical Species; Fertilization; Forest Nutrition.
RESUMO
A fertilização é determinante do crescimento e desenvolvimento de árvore urbanos. Por isso estudamos através da técnica do elemento faltante, os requerimentos nutricionais de cinco espécies tropicais usadas em silvicultura urbana (Tabebuia chrysantha, Margaritaria nobilis, Hamelia patens, Apeiba aspera, Cupania americana). Encontramos algumas espécies sensíveis à deficiência de nitrogênio e outras à deficiência de enxofre. O grupo de T. chrysanta, M. nobilis y H. patens mostraram alta sensibilidade à deficiência de nitrogênio, enquanto C. americana y A. aspera apresentaram sensibilidade à deficiência de enxofre, simbolizavam um menor crescimento promédio para todas às variáveis avaliadas: altura, diâmetro da raiz, matéria seca radical e matéria seca aéreo. As representações HJ-Biplot, utilizadas para caracterizar a resposta por espécie a cada tratamento, em todas as unidades experimentais, evidenciaram baixos rendimentos em crescimento para às deficiências em nitrogênio e enxofre. O desordem de bases cálcio/magnésio e magnésio/potássio afetou o crescimento em altura de todas as espécies, evidenciado nos altos crescimentos obtidos para os tratamentos deficientes em cálcio e magnésio. Devido a isso, as formulas nutricionais utilizadas em viveiros devem-se ajustar para produzir melhores árvores para a cidade.
PALAVRAS-CHAVE: Silvicultura urbana; Espécies tropicais; Fertilização; Nutrição florestal.
1. INTRODUCCIÓN
La producción y fertilización de árboles urbanos ha sido estudiada en zonas templadas y subtropicales alrededor del mundo (Watkins, 1998, Harris, et al., 2008, Sæbø y Ferrini, 2006, Ferrini y Baietto, 2006). Cada especie vegetal toma los nutrientes del suelo conforme a sus necesidades, por tanto el nivel de fertilidad edáfico es uno de los mayores limitantes para los árboles urbanos (Stanley y Montagnini, 1999, Schimann et al., 2008, Ferrini y Alesio, 2010). Los análisis de fertilidad de suelo y foliar se han empleado para estudiar los nutrientes que usan las plantas en su crecimiento (Salas y Ramírez, 2001). Además de estos, la técnica del elemento faltante, descrita por Jenny, et al. (1950), se utiliza para develar las deficiencias nutricionales en los suelos en que crecen las plantas (Salas y Ramírez, 2001). Sin embargo, también es posible aplicar esta técnica para conocer las necesidades nutricionales de árboles urbanos en sus dos etapas: producción en vivero y establecimiento en bosques urbanos. Así, es posible identificar los nutrientes que juegan un papel fundamental en el crecimiento de las especies.
Los elementos nutricionales claves que limitan el crecimiento de especies tropicales usadas en silvicultura urbana son poco conocidos. Conforme aumenta la población mundial los bosques urbanos cobran vital importancia, debido a los múltiples servicios que prestan (Madureira et al., 2011, Soares et al., 2011, Ferrini y Alesio, 2010, Molina y Vargas, 2007). Allí, los árboles son más susceptibles a sufrir ataques por plagas, daños mecánicos y deficiencias nutricionales, que ocasionan altas tasas de mortalidad (Ferrini y Alesio, 2010). Un aspecto clave de la silvicultura urbana es garantizar la supervivencia de los árboles que se produzcan y planten, la cual depende, entre otros factores, del nivel original de fertilidad del suelo y de la fertilización suministrada, bien sea en la etapa de producción (vivero) o en la etapa de mantenimiento en el sitio de plantación. El conocimiento de los requerimientos nutricionales particulares de las especies nativas, utilizadas en programas de silvicultura urbana, puede contribuir a incrementar las tasas de crecimiento radical, en altura y biomasa de las plántulas en vivero, así como a reducir las tasas de mortalidad, una vez plantadas en el entorno urbano (Hoyos, et al., 2007, Herrera, 2009). Por ello, es esencial desarrollar herramientas que permitan identificar los elementos nutricionales que limitan el crecimiento de las especies tropicales que se producen en vivero para conformar los bosques urbanos.
Este trabajo tuvo como objetivo identificar los elementos que juegan un rol fundamental en la nutrición en vivero de cinco especies tropicales empleadas en bosques urbanos en Medellín, Colombia. Estas especies son Tabebuia chrysantha (Jacq.) Nichol., Cupania americana L., Hamelia patens Jacq., Margaritaria nobilis L.f. y Apeiba aspera Aubl. Con esto se pretende aportar herramientas para la formulación de programas de fertilización de estas especies tanto en vivero como en bosques urbanos, que contribuyan a la optimización de su crecimiento y desarrollo iniciales, y su establecimiento y supervivencia en campo.
2. METODOLOGÍA
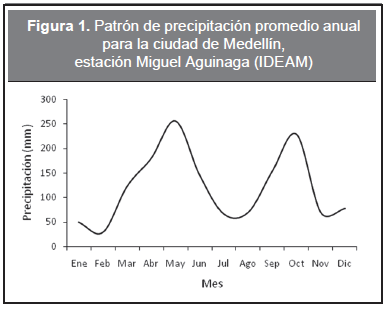
El experimento consistió en emplear la técnica del elemento faltante, descrita por Jenny, et al. (1950), en cinco especies arbóreas usadas en la silvicultura urbana de la ciudad de Medellín, Colombia; estas especies son: Tabebuia chrysantha (Jacq.) Nichol., Cupania americana L., Hamelia patens Jacq., Margaritaria nobilis L.f. y Apeiba aspera Aubl. Se estableció un experimento independiente para cada especie, con diseño al azar, con 10 tratamientos y 10 repeticiones por tratamiento. La unidad experimental consistió en una planta sembrada en bolsa plástica de 20 cm de diámetro por 30 cm de alto. Las plantas fueron germinadas en cuarzo y se trasplantaron a la bolsa cuando tenían 10 cm de alto. Los tratamientos se formularon basándose en los análisis de fertilidad de suelo, realizados a cada uno de los suelos empleados en los experimentos (Tabla 1). Se emplearon dos suelos muy similares. Los suelos derivados de ceniza volcánica provenientes de material de corte, horizonte A, de la afueras de la ciudad de Medellín. Se empleó el Suelo 1 para T. chrysantha, C. americana y H. patens; y el Suelo 2 para M. nobilis y A. aspera. A cada planta se le aplicó, luego del trasplante, 200 cm3 de la solución nutritiva formulada correspondiente a cada tratamiento específico y no se volvió a fertilizar. Los tratamientos fueron: un tratamiento testigo (T0), el cual consistió únicamente en suelo sin fertilización; un tratamiento completo (TC), el cual consistió en fertilizar el suelo con todos los nutrientes necesarios para el crecimiento óptimo de las plantas; siete tratamientos para inducir deficiencias (-N, -P, -K, -Ca, -Mg, -S, -B), cada uno consistente en la misma fórmula del tratamiento completo menos el nutriente deficitario que se deseó evaluar. Y por último, un tratamiento comparativo (ST), que consistió en sembrar las plántulas en el sustrato utilizado en el vivero municipal de la ciudad de Medellín en sus actividades de producción. Este sustrato consiste en una mezcla 4:3:3 de tierra, cascarilla de arroz y compost, y triple 18 (N:P:K), respectivamente, sin agregarse ningún otro fertilizante. Los experimentos se realizaron en las instalaciones del vivero municipal de la ciudad de Medellín; el régimen de precipitaciones es bimodal (Figura 1) y la temperatura varía entre 19 y 24 °C. Se midió mensualmente durante 4 meses la altura (A) y el diámetro en el cuello de la raíz (D) en todas las plántulas utilizando un pie de rey digital sobre la marca realizada en la primera medición. Finalizados los 4 meses, se cosecharon las plantas y se obtuvo la masa seca aérea (Ma) y la masa seca radical (Mr) de cada planta mediante un proceso de secado en horno a 65 °C hasta alcanzar peso constante. Luego se calculó el índice de esbeltez propuesto por Villar (2003), como la relación entre la materia seca aérea (Ma) y la materia seca radical (Mr).
La altura y el diámetro en el cuello de la raíz se analizaron mediante un estudio de varianza (ANDEVA) con medidas repetidas en el tiempo. Esto es, un examen de varianza de dos vías (tratamientos y tiempo), en el que se modela la estructura de covarianzas mediante el uso de modelos mixtos. En el ANDEVA por tratamientos, de estas dos variables, se usaron los rendimientos absolutos al momento final del experimento. Se ajustaron las covarianzas mediante modelos mixtos y se estimaron los errores estándar más adecuados para las diferentes comparaciones, acorde con las características de cada conjunto de datos (Littell, et al., 1998), usando PROC MIXED de SAS, versión 9.0. Para las variables masa seca aérea y radical se empleó un ANDEVA simple. Se obtuvieron representaciones HJ-Biplot para cada especie (Galindo, 1986). Esta técnica multivariada permite resumir en un plano la información de las variables y de las unidades experimentales de manera simultánea (Correa, et al., 2007), así como observar la relación entre las variables según el ángulo entre los planos factoriales; para el efecto se utilizó una rutina programada en Matlab®, versión 7.0.
3. RESULTADOS
3.1 T. chrysantha
El nitrógeno fue el elemento con mayor incidencia sobre el crecimiento y desarrollo de las plantas; el tratamiento deficiente en nitrógeno dio lugar a los menores rendimientos a nivel de la muestra (desempeño con respecto al tratamiento completo), en todas las variables; para masa seca radical no se observaron diferencias significativas entre los tratamientos -N, -S y ST (Tabla 3). No se observaron diferencias significativas en el índice de esbeltez para ningún tratamiento, sin embargo los mayores valores se observaron bajo el tratamiento -N (4,8) y los menores valores bajo el tratamiento -P (2,8) (Tabla 3). El ANDEVA con medidas repetidas en el tiempo para altura y diámetro del cuello de la raíz mostró diferencias significativas entre los tiempos de medición 1 y 4 en todos los tratamientos como lo muestra la Figura 3. El primer plano factorial de la representación HJ-Biplot (Figura 2A), recogió el 90,2 % de la variabilidad total. Con este método se logra una disminución en el número de tratamientos, pues se eliminaron aquellos que tenían comportamientos muy alrededor del centro e introducían ruido. Todas las unidades experimentales sometidas al tratamiento -N obtuvieron bajos rendimientos en las cuatro variables; mientras que el tratamiento -Ca benefició la mayoría de las unidades experimentales en masa seca radical y masa seca aérea, sin diferenciarse estadísticamente del tratamiento deficiente en Mg y P. Los tratamientos -P y -S generaron los mejores resultados para altura, mientras que el -Mg determinó los mayores valores para diámetro en el cuello de la raíz, diferenciándose únicamente de -N (Figura 2A, Tabla 3). Además, se encontró relación positiva entre las variables de biomasa y las variables biométricas (Figura 2A).
3.2 C. americana:
Varios tratamientos mostraron bajos rendimientos sin diferencias significativas entre ellos. El tratamiento -S fue el que reportó los rendimientos más bajos pero nunca difirió de TC en ninguna variable (Tabla 3). Las plantas bajo el tratamiento ST presentaron alto rendimiento en todas las variables, al igual que bajo -N para diámetro del cuello de la raíz y -Ca para masa seca radical. C. americana fue la única especie que respondió bien al tratamiento comparativo ST, diferenciándose estadísticamente del resto de los tratamientos. El índice de esbeltez no se diferenció estadísticamente en ningún tratamiento, pero el mayor valor se alcanzó para el tratamiento -K y el menor valor se alcanzó para el tratamiento T0. El tiempo influyó en altura y diámetro del cuello de la raíz entre los tiempos de medición 1 y 4 del experimento (Figura 3). El primer plano factorial de la representación HJ-Biplot, recogió el 89,37 % de la variabilidad total (Figura 2B). El tratamiento ST afectó la mayoría de la unidades experimentales mejorando su rendimiento, en contraste con los tratamientos -S y TC que mostraron bajos resultados al igual que la mayoría de tratamientos como se puede observar en la Figura 2B y la Tabla 3.
3.3 H. patens:
Las plantas bajo los tratamientos ST y T0 mostraron la mayor limitación para el crecimiento y desarrollo, en todas las variables; -N solo afecto la masa seca aérea disminuyendo su rendimiento como se observa en la Tabla 3 y la Figura 2E. Los mejores rendimientos se obtuvieron para el tratamiento completo (TC), en cuanto a diámetro del cuello de la raíz, masa seca aérea y masa seca radical, sin diferir significativamente de otros tratamientos (Tabla 3). En altura los mejores rendimientos se obtuvieron para varios tratamientos (-Mg, -Ca, -S, -B, y -K), siendo ST, T0, -N, TC y -P los tratamientos más limitantes en el desarrollo de H. patens. El tratamiento en el que más alto índice de esbeltez se obtuvo fue -S sin diferir estadísticamente de los tratamientos -B, -Mg, -Ca y -K; los tratamientos para los que se obtuvieron menores valores del índice de esbeltez fueron ST y T0, sin diferir de -N, -P y -TC (Tabla 3). Existieron diferencias significativas para el diámetro del cuello de la raíz y la altura entre los tiempos 1 y 4 (Figura 3). El primer plano factorial de la representación HJ-Biplot (Figura 2E) recogió el 92,47 % de la variabilidad total. Se observó una estrecha relación positiva entre materia seca aérea y diámetro en el cuello de la raíz (Figura 2E). El tratamiento -S tuvo una influencia positiva en altura, en todas las unidades experimentales (Figura 2E).
3.4 M. nobilis:
Las plantas bajo los tratamientos ST, -N y T0 mostraron los menores rendimientos, en todas las variables (Tabla 3); para masa seca radical no presentaron diferencias significativas con -S, -Mg, -B, -K y -Ca (Tabla 3). Por tanto si se quiere promover el crecimiento en masa seca de raíces se debería optar por el TC disminuyendo un poco el fósforo. Los mejores rendimientos ocurrieron en los tratamientos completo y deficiente en fósforo para todas las variables, con excepción de la altura, para la que los tratamientos con mejor rendimiento fueron -Mg y -Ca, similares además al tratamiento completo (Tabla 3). El índice de esbeltez varió considerablemente entre tratamientos, siendo -S y -Mg los que mejores resultados arrojaron y -P y TC los más limitantes, sin diferir de T0, -Ca y -K. Solo se observaron diferencias significativas para altura y diámetro en el cuello de la raíz entre los tiempos de medición 1 y 4, las otras dos variables no se diferenciaron en el tiempo (Figura 3). El primer plano factorial de la representación HJ-Biplot (Figura 2C) recogió el 89,38 % de la variabilidad total. Se observó una asociación positiva entre las variables de biomasa y diámetro en el cuello de la raíz (Figura 2C). Los tratamientos -P, TC y -Ca incrementaron el crecimiento de la mayoría de las unidades experimentales en diámetro, masa seca radical y masa seca aérea (Figura 2C).
3.5 A. aspera:
Las plantas bajo los tratamientos ST y T0 presentaron los menores rendimientos en masa seca aérea y altura, y bajo ST y -S en diámetro en el cuello de la raíz y masa seca radical; sin embargo, no se diferenciaron de varios tratamientos en todas las variables (Tabla 3). El tratamiento TC mostró mejores rendimientos, pero no hubo diferencias significativas con otros como -K, -P para masa seca radical, -Ca para diámetro en el cuello de la raíz y -Ca, -Mg, -K y -B para altura (Tabla 3). Los tratamientos -S, -N, -Mg y -Ca reportaron los mejores rendimientos para el índice de esbeltez, mientras que los tratamientos ST, -B, TC, -P, T0 y -K mostraron los rendimientos más bajos, sin diferenciarse de -N, -Mg y -Ca. Solo se encontraron diferencias significativas entre los tiempos de medición 1 y 4 para la variable altura en los tratamientos deficientes en nitrógeno, potasio, calcio, azufre y el tratamiento testigo, el resto no presentaron diferencias significativas en el tiempo (Figura 3). El primer plano factorial de la representación HJ-Biplot (Figura 2D) recogió el 92,88 % de la variabilidad total. Se encontró asociación positiva entre el grupo de variables biométricas y entre el de variables de biomasa (Figura 2D). El tratamiento completo aporta un claro beneficio en el crecimiento en todas las variables para casi todas las unidades experimentales (Figura 2D).
4. DISCUSIÓN
Los nutrientes claves en el crecimiento de las cinco especies estudiadas en este ensayo fueron el nitrógeno y el azufre, sin embargo deficiencias de otros elementos también afectaron el crecimiento en las cuatro variables evaluadas en las cinco especies (Tabla 3). Esto confirma la hipótesis de que las especies tropicales tienen complejas, diferentes y contrastantes necesidades nutricionales. Se podría, de forma muy general, clasificar en dos grupos las especies aquí estudiadas: T. chrysantha, H. patens y M. nobilis sensibles a la deficiencia de nitrógeno y A. aspera y C. americana sensibles a la deficiencia de azufre. Algunos estudios han encontrado que deficiencias de nitrógeno, fósforo y potasio presentan el mayor carácter limitante para el crecimiento de numerosas especies de árboles tropicales, dado que estos nutrientes tienen problemas de disponibilidad en el suelo y son requeridos por las plantas en grandes cantidades (Davidson, et al., 1998, Salas y Ramírez, 2001, Pedrol, et al., 2010). Sin embargo en este estudio se reporta la cantidad asimilable de azufre como elemento clave en la nutrición de A. aspera y C. americana, mientras que en T. chrysantha, H. patens y M. nobilis la presencia de azufre pareciera limitar el crecimiento en altura. Los resultados de este ensayo sugieren que los suelos utilizados fueron deficientes principalmente en nitrógeno y azufre, dado que el crecimiento de las especies en ausencia de tales elementos fue similar al del tratamiento testigo y difirió del tratamiento completo y del tratamiento que mostró los mejores resultados (Figura 1, Tabla 3). Estas respuestas contrastantes manifestadas como deficiencias nutricionales, es probable que resulten de las condiciones fisiológicas y demandas particulares de cada especie (Nussbaum et al., 1995, Stanley y Montagnini, 1999).
La mayoría de las especies tuvieron menor crecimiento bajo el tratamiento comparativo (ST) respecto a los otros tratamientos, excepto C. americana. Este es un sustrato genérico que se usa en los viveros y se emplea para plantar los árboles en la ciudad. En contraste, el tratamiento completo (TC) y algunos otros tratamientos mostraron resultados positivos en el crecimiento de las plántulas de T. chrysantha, H. patens, M. nobilis y A. aspera (Figura 2). Este resultado pone de presente la necesidad de mejorar las fórmulas de fertilización utilizadas en los viveros y en la siembra de árboles urbanos, para ajustarse a los requerimientos básicos de cada especie. Todo esto con el objetivo de disminuir el tiempo de permanencia en vivero y aumentar la sobrevivencia y longevidad de árboles urbanos. Para el efecto, se sugiere emplear fórmulas de fertilización alternativas que permitan obtener mayor crecimiento en vivero para las especies aquí estudiadas: se recomienda para el grupo T. chrysantha, H. patens y M. nobilis emplear una fórmula similar a la utilizada en el tratamiento completo (TC, Tabla 2), pero enriquecida con nitrógeno y con un equilibrio entre bases que promueva el crecimiento en biomasa; para A. aspera usar una fórmula similar a la usada en el tratamiento completo, pero enriquecida en azufre y un equilibrio entre bases Mg/K; y para C. americana usar una fórmula similar a la del tratamiento comparativo (ST). Es indispensable avanzar en futuras investigaciones para definir las cantidades óptimas de estos elementos, y comprender mejor la nutrición de árboles tropicales. En algunos estudios se ha encontrado que sustratos similares al utilizado en el tratamiento ST corrigen las deficiencias de fósforo y el desbalance de bases (Macci, et al., 2012, Pedrol, et al., 2010). Esto podría explicar los mayores rendimientos obtenidos en el tratamiento comparativo (ST) para todas las variables en C. americana; pero las especies restantes presentaron bajos rendimientos en este tratamiento, a pesar de los altos contenidos de nutrientes del tratamiento comparativo ST. En algunos sustratos orgánicos, como el empleado en el tratamiento comparativo, puede ser difícil la mineralización del nitrógeno contenido en la materia orgánica, tardando en quedar disponible para las plantas (Salifu y Timmer, 2001). Se cree que otro factor sumado a la afectación en el crecimiento de las demás especies, sea el alto contenido de potasio en el tratamiento comparativo (ST) (Tabla 1), lo cual supone un desbalance de bases Mg/K muy severo (0,52) y muy inferior al reportado como óptimo (Mg/K = 3,3; (Stover y Simmonds, 1987). Para A. aspera, la dependencia de nitrógeno no fue acentuada y mostró la misma dependencia al azufre que C. americana. Sin embargo, C. americana mostró afectación severa cuando se le restringió el suministro de magnesio, deficiencia que debió subsanarse en el tratamiento comparativo (ST). Por el contrario, A. aspera presentó los mejores rendimientos para la deficiencia de potasio en las variables de biomasa, debido, posiblemente, a su alta sensibilidad al desbalance de bases magnesio-potasio (Mg/K). Los autores de este estudio piensan que el alto contenido de potasio presente en el tratamiento comparativo afectó el crecimiento de A. aspera, como se ha encontrado en otros estudios (dos Reis et al., 2012, Navarro, 2003). Asimismo, el alto desempeño de esta especie bajo el tratamiento -Ca evidencia que el excesivo contenido de esta base en el tratamiento comparativo causa un desbalance de bases Ca/Mg que disminuyó el crecimiento de esta especie. Es posible que A. aspera no presentara crecimiento significativo en altura entre los primeros meses del ensayo debido a la magnitud de la limitación que el desbalance de bases implica para la especie. Esto pone de presente que la nutrición de especies de árboles tropicales requiere de particular atención no solo con respecto a las cantidades de nutrientes disponibles en el suelo, sino a las proporciones entre ellos.
Desequilibrios en las bases intercambiables del suelo afectan el crecimiento y desarrollo vegetal (Wilmot, et al., 1996). Grandes cantidades de bases intercambiables en el suelo pueden causar un desbalance nutricional para las plantas y reversar su crecimiento y desarrollo (Pedrol, et al., 2010). Al parecer esta situación ocurrió en la producción de masa seca aérea y radical de las especies T. chrysantha y C. americana, para las que sin suministro de calcio (-Ca) se registraron los mejores rendimientos, incluso por encima del tratamiento completo (Tabla 3). Hay que notar que, los contenidos de calcio y magnesio en los suelos utilizados para el ensayo se encontraban en niveles muy bajos (Tabla 1); por tanto no se espera esta situación. En ocasiones no son las cantidades de nutrientes las que limitan el desarrollo vegetal, sino la proporción en que estos se encuentran en el suelo. Así, para las bases intercambiables, una inadecuada relación calcio-magnesio (Ca/Mg) es causante de los principales desbalances que actúan como limitantes del crecimiento (Pedrol, et al., 2010). Se ha reportado que las enmiendas a base de fertilizantes químicos pueden crear desbalances Ca/Mg por la excesiva oferta de alguno de estos dos cationes (Wilmot, et al., 1996), lo cual posiblemente ha ocurrido con los ensayos donde se adicionó calcio o magnesio. Los valores de la relación Ca/Mg para los suelos empleados (5,1 y 3,1 para los suelos 1 y 2, respectivamente) fueron muy superiores a los indicados como apropiados (Ca/Mg = 2). Es factible pensar que la adición de calcio acentuó el desequilibrio de bases en el suelo, lo cual fue más notorio en el suelo 1 que en el suelo 2, en el cual el valor Ca/Mg = 3,1 pudiera ser aceptable. Esta podría ser una de las causas que determinaron los mayores rendimientos obtenidos para las tres especies cultivadas en el suelo 1 bajo el tratamiento deficiente en calcio. Las especies A. aspera y M. nobilis, cultivadas en el suelo 2, no mostraron gran afectación por los cambios en los contenidos de estas bases intercambiables, posiblemente debido a que la relación de estas bases en el suelo no presentaba valores muy por encima de los óptimos. En algunos suelos el desequilibrio magnesio-potasio (Mg/K) genera problemas en la capacidad de intercambio catiónico (CIC) y en la absorción de nutrientes (Pedrol, et al., 2010). En los suelos aquí utilizados el contenido de potasio fue de medio a alto y su relación con el magnesio estuvo relativamente equilibrada. Sin embargo, A. aspera aumentó su producción de masa seca radical (Mr) en el tratamiento deficiente en potasio, lo cual puede indicar su mayor sensibilidad al balance Mg/K, y aun más en la producción de raíces.
El índice de esbeltez (Ma/Mr) mostró coherencias con los resultados ya encontrados para la mayoría de las especies en algunos de los parámetros biométricos, así las deficiencias que más limitaron el crecimiento mostraron a su vez las relaciones Ma/Mr más desproporcionadas, exceptuando M. nobilis que mostró las mayores desproporciones para los tratamientos menos limitantes. Este es el primer trabajo que compara índices de esbeltez para cada una de estas especies en su producción y fertilización en vivero. Valores bajos obtenidos de la proporción entre la masa seca aérea y la masa seca radical suelen indicar, a nivel interespecífico, una mayor eficiencia en el uso del agua (Stewart y Bernier, 1995, Villar, 2003), ya que las plantas pueden mantener un mejor estado hídrico con un consumo más moderado de agua en situaciones de deficiencia hídrica. Esto mejora la supervivencia de las plantas levantadas en vivero y luego sembradas en ambientes hostiles como la mayoría de los espacios urbanos (Lloret et al., 1999). En este trabajo se observó que cada especie tiene un rango de valores promedio asociados para Ma/Mr y, diferente sensibilidad a las deficiencias nutricionales. Por ejemplo, T. chrysantha y C. americana no fueron sensibles a las deficiencias nutricionales pues ningún tratamiento mostró diferencias para esta relación; A. aspera fue medianamente sensible pues solo un tratamiento (-N) tuvo efecto positivos en la relación Ma/Mr y; H. patens y M. nobilis fueron sensibles a las deficiencias nutricionales, las cuales afectaron de forma significativa su relación Ma/Mr. M. nobilis fue la única especie que presentó la mayor desproporción Ma/Mr en los tratamientos que optimizaron su crecimiento (TC y -P). Sin embargo, los valores óptimos de Ma/Mr para estas especies deberán evaluarse mediante ensayos de sobrevivencia y crecimiento, de esta manera podría determinarse con mayor exactitud el tipo de planta óptima para uso urbano y su tratamiento de fertilización asociado, pues los alcances de esta investigación no permiten llegar a esta conclusión.
Finalmente, el crecimiento de un árbol urbano supone un costo nutricional para el sitio, un costo de oportunidad y un costo logístico, entre otros costos, con lo cual, si no se asegura la supervivencia del árbol se incurriría en pérdidas económicas considerables. Por ello es necesario incrementar la investigación en múltiples aspectos de la silvicultura urbana que aporten al conocimiento que se requiere para construir bosques urbanos sostenibles. Un aspecto importante en la supervivencia de los árboles urbanos tropicales es la nutrición. Aplicar en cantidades óptimas los elementos nutricionales limitantes para cada especie aumenta su probabilidad de sobrevivencia y contribuye a la sostenibilidad de los bosques urbanos. Hay que ahondar más en investigaciones de este tipo para desarrollar enmiendas nutricionales para cada especie utilizada en silvicultura urbana y producida en los viveros con este objetivo.
5. CONCLUSIONES
Las especies de árboles tropicales presentan comportamientos nutricionales complejos, que no solo se limitan a las cantidades de elementos nutricionales disponibles en el suelo, sino que también dependen de la relación que estos elementos guardan entre ellos. Es posible identificar grupos de especies que responden a patrones nutricionales comunes como se encontró en este estudio para las deficiencias de nitrógeno y azufre. Algunas especies como A. aspera, T. chrysantha y C. americana son altamente sensibles al desequilibrio de bases, afectando la producción de masa seca radical y aérea. Esto supone que, para especies tan sensibles a la desigualdad de bases, es fundamental realizar su precisa y adecuada formulación nutricional.
La técnica del elemento faltante es útil para comprender la forma como las plantas usan los diferentes nutrientes disponibles en el suelo. Con base en estos resultados, se recomendaría usar fórmulas de fertilización, específicas para cada especie, que se asemejen a las fórmulas utilizadas en el tratamiento completo (TC) ajustadas a las deficiencias y desequilibrios de bases que afectan el crecimiento y desarrollo de las especies de forma negativa. Además, estas fórmulas deben estar basadas en un análisis previo de fertilidad de suelo o del sustrato donde se vaya a sembrar las plantas. Estas fórmulas podrían funcionar muy bien en la producción de plantas en vivero, y dar un referente nutricional para la siembra de árboles urbanos.
AGRADECIMIENTOS
Se agradece a la Alcaldía de Medellín, en especial a la Unidad de Paisajismo, Zonas Verdes y Arborización de la Secretaría de Obras Públicas, por su apoyo e interés en la ejecución de este estudio. Se hace reconocimiento a B. Calle por su participación en algunas actividades de vivero y laboratorio.
REFERENCIAS
Correa, L.; Lavalett, O.; Galindo, V. (2007). Uso de métodos multivariantes para la agrupación de aislamientos de Colletotrichum spp. con base en características morfológicas y culturales. Revista Facultad Nacional de Agronomía 60, pp. 3671-3690. [ Links ]
Davidson, R.; Gagnon, D.; Mauffette, Y. and H., H. (1998). Early Survival, Growth and Foliar Nutrients in Native Ecuadorian Trees Planted on Degraded Volcanic Soil. Forest Ecology and Management 105, 1-19. [ Links ]
Dos Reis, B.E.; De Paiva, H. N.; Barros, T. C.; Ferreira, A. L. and Cardoso, W. D. (2012). Growht and Seedling Quality of Jacaranda-da-Bahia in Response to Potassium and Sulfur Fertilization. Ciencia Florestal, 22, 389-U1505. [ Links ]
Ferrini, F. and Alesio, A. (2010). Sustainable Management Techniques for Trees in the Urban Areas. Journal of Biodiversity and Ecological Sciences 1, pp. 1-20. [ Links ]
Ferrini, F.; and Baietto, M. (2006). Response to Fertilization of Different Tree Species in the Urban Environment. Arboriculture and Urban Forestry 32(3), May, pp. 93-99. [ Links ]
Galindo, M. P. (1986). Una alternativa de representación simultánea: HJ-biplot. Questió, 10, pp. 13-23. [ Links ]
Harris, J. R.; Day, S. D. and Kane, B. (2008). Nitrogen Fertilization During Planting and Establishment of the Urban Forest: A Collection of Five Studies. Urban Forestry and Urban Greening, 7, pp. 195-206. [ Links ]
Herrera, S. (2009). Árboles de la Universidad del Valle. Santiago de Cali, Colombia, Cali, Colombia, Programa Editorial Universidad del Valle. [ Links ]
Hoyos, M.; Cogollo, A. and Villa, D. (2007). Silvicultura Urbana para Medellín, Medellín, Fondo editorial Jardín Botánico de Medellín. [ Links ]
Jenny, H.; Vlamis, J. and Martin, W. (1950). Greenhouse assay of fertility of California soils. Hilgardia, 20, pp. 1-18. [ Links ]
Littell, R.; Henry, P. and Ammerman, C. (1998). Statistical Analysis of Repeated Measures Using SAS Procedures. Journal of Animal Science 76, pp. 1216-1231. [ Links ]
Lloret, F.; Casanovas, C. and Peñuelas, J. (1999). Seedling Survival of Mediterranean Shrubland Species in Relation to Root: Shoot Ratio, Seed Size and Water and Nitrogen Use. Functional Ecology, 13, pp. 210-216. [ Links ]
Macci, C.; Doni, S.; Peruzzi, E.; Masciandaro, G.; Mennone, C. and Ceccanti, B. (2012). Almond Tree and Organic Fertilization for Soil Quality Improvement in Southern Italy. Journal of environmental management, 95 Suppl, pp. S215-22. [ Links ]
Madureira, H.; Andresen, T. and Monteiro, A. (2011). Green Structure and Planning Evolution in Porto. Urban Forestry and Urban Greening, 10, pp. 141-149. [ Links ]
Molina, L. and Vargas, E. (2007). Árboles para Villavicencio: Especies que fortalecen la estructura ecológica principal. Revista Nodo 2, pp. 85-98. [ Links ]
Navarro, G. (2003). Química Agrícola, Madrid, España, Mundi-Prensa. [ Links ]
Nussbaum, R.; Anderson, J. and Spencer, T. (1995). Factors Limiting the Growth of Indigenous Tree Seedlings Planted on Degraded Rainforest Soils in Sabah, Malaysia. Forest Ecology and Management 74, pp. 449-459. [ Links ]
Pedrol, N.; Puig, C. G.; Souza, P.; Forján, R.; Vega, F. A.; Asensio, V.; González, L.; Cerqueira, B.; Covelo, E. F. and Andrade, L. (2010). Soil Fertility and Spontaneous Revegetation in Lignite Spoil Banks under Different Amendments. Soil and Tillage Research, 110, 134-142. [ Links ]
Saebo, A. and Ferrini, F. (2006). The Use of Compost in Urban Green Areas - A Review for Practical Application. Urban Forestry and Urban Greening, 4, pp. 159-169. [ Links ]
Salas, E. and Ramírez, C. (2001). Determinación del N y P en abonos orgánicos mediante la técnica del elemento faltante y un bioensayo microbiano. Agronomía Costarricense 25, 25-34. [ Links ]
Salifu, K. F. and Timmer, V. R. (2001). Nutrient Retranslocation Response of Picea mariana Seedlings to Nitrogen Supply. Soil Science Society of America Journal, 65, pp. 905-913. [ Links ]
Schimann, H.; Ponton, S.; Hättenschwiler, S.; Ferry, B.; Lensi, R.; Domenach, A.-M. and Roggy, J.-C. (2008). Differing Nitrogen Use Strategies of Two Tropical Rainforest Late Successional Tree Species in French Guiana: Evidence from 15N Natural Abundance and Microbial Activities. Soil Biology and Biochemistry, 40, pp. 487-494. [ Links ]
Soares, A. L.; Rego, F. C.; Mcpherson, E. G.; Simpson, J. R.; Peper, P. J. and Xiao, Q. (2011). Benefits and Costs of Street Trees in Lisbon, Portugal. Urban Forestry and Urban Greening, 10, pp. 69-78. [ Links ]
Stanley, W. G. and Montagnini, F. (1999). Biomass and Nutrient Accumulation in Pure and Mixed Plantations of Indigenous Tree Species Grown on Poor Soils in the Humid Tropics of Costa Rica. Forest Ecology and Management, 113, pp. 91-103. [ Links ]
Stewart, J. and Bernier, P. (1995). Gas Exchange and Water Relations of 3 Sizes of Containerized Picea Mariana Seedlings Subjected to Atmospheric and Edaphic Water Stress Under Controlled Conditions. Annals of Forestry Science, 52, pp. 1-9. [ Links ]
Stover, R. and Simmonds, N. (1987). Bananas, London, UK, Longman Scientific and Tecnichal. [ Links ]
Villar, P. (2003). Importancia de la calidad de planta en los proyectos de revegetación En: Rey-Benayas, J. M.; Espigares, T. and Nicolau-Ibarra, J. M. (eds.) Restauración de Ecosistemas Mediterráneos. Alcalá de Henares: Universidad de Alcalá / Asociación Española de Ecología, Terrestre. [ Links ]
Watkins, J. R. 1998. Fertilization and Woody Plant Nutrition in the Context of the Urban Forest. Master of Forestry Ms.c., Virginia Polytechnic Institute and State University. [ Links ]
Wilmot, T. R.; Ellsworth, D. S. and Tyree, M. T. 1996. Base Cation Fertilization and Liming Effects on Nutrition and Growth of Vermont Sugar Maple Stands. Forest Ecology and Management, 84(1), pp. 123-134. [ Links ]
