










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Lasallista de Investigación
Print version ISSN 1794-4449
Rev. Lasallista Investig. vol.6 no.1 Caldas Jan./June 2009
Artículo original
Metodologías utilizadas para la medición de las pérdidas intestinales de nitrógeno endógeno en cerdos
Methodologies used for the measurement of endogenous intestinal nitrogen losses in growing pigs
Metodologias utilizadas para a medição das perdas intestinais de nitrogênio endógeno em porcos.
José Julián Echeverri Zuluaga*, Andrés Goméz Zapata**, Jaime Eduardo Parra Suescún***
* Zootecnista, MsC Biotecnología Animal, Profesor Auxiliar, Departamento de Producción Animal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín.
** Zootecnista, Aspirante a Maestría en Biotecnología Animal, Universidad Nacional de Colombia, Sede Medellín, Facultad de Ciencias.
*** Zootecnista, MsC Nutrición Animal, Profesor Auxiliar, Departamento de Producción Animal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín. jeparrasu@unalmed.edu.co
Resumen
La digestibilidad ileal de los aminoácidos es uno de los factores más importantes para determinar el valor nutricional de un alimento. Sin embargo, los valores obtenidos en las pruebas de digestibilidad ileal pueden variar de acuerdo con las pérdidas de nitrógeno endógeno del animal y por esto, pueden ser expresados como aparentes o verdaderos. En la determinación del valor aparente no se tienen en cuenta las pérdidas de nitrógeno endógeno, por lo que se subestima la cantidad de aminoácidos absorbidos durante la digestión. Al tomar en cuenta el componente endógeno, la digestibilidad ileal se transforma en verdadera, siendo ésta el mejor parámetro para representar los aminoácidos absorbidos en el tracto digestivo. Las pérdidas de nitrógeno endógeno han sido clásicamente definidas como el nitrógeno encontrado en la digesta o en las heces de animales alimentados con dietas libres de nitrógeno. Tradicionalmente, las pérdidas de nitrógeno endógeno en cerdos han sido determinadas mediante los métodos de regresión lineal o por el de ofrecer dietas libres de nitrógeno. Actualmente se han desarrollado varias técnicas alternativas para estimar las pérdidas de nitrógeno endógeno en cerdos que consumen dietas que contienen niveles variables de proteína. Cada una de estas técnicas posee limitaciones y todas requieren diferentes suposiciones. Basados en un creciente número de resultados obtenidos con diferentes técnicas, las pérdidas de nitrógeno endógeno parecen estar altamente afectadas por el método utilizado para su determinación y por factores como son peso corporal, consumo de materia seca, y constituyentes de la dieta (factores antinutricionales, fibra y proteína).
Palabras clave: Flujo de nitrógeno endógeno, cerdos en crecimiento, técnicas de cuantificación.
Abstract
Amino acid ileum digestibility is one of the most important factors to determine the nutritional value of foods. However, the values obtained in ileum digestibility tests can vary according to the endogenous nitrogen losses in the animal. Thus, ileum digestibility values can be classified as apparent or true. When determining the apparent value, endogenous nitrogen losses are not taken into account and the amounts of absorbed amino acid during digestion are then underestimated. When the endogenous component is taken into account, digestibility data become true, being the best parameter to represent absorbed amino acid in the digestive tract. Endogenous nitrogen losses are classically defined as the nitrogen found in the digesta or in the feces of animals fed with nitrogen-free diets. Traditionally, endogenous nitrogen losses in pigs have been determined by feeding nitrogen-free diets or by the lineal regression method. More recently, several alternative techniques have been developed to evaluate endogenous nitrogen losses in pigs that are fed with diets that contain variable protein levels. Each of these techniques has some limitations and all require different assumptions. Based on the increasing number of results recently obtained with different techniques, endogenous nitrogen losses seem to be highly affected by the used method and by factors as weight, dry matter consumption, and diet components (anti nutritional factors, fiber and protein).
Keywords: Endogenous nitrogen flow, growing pigs, quantification techniques.
Resumo
A digestibilidad ileal dos aminoácidos é um dos fatores mais importantes para determinar o valor nutricional de um alimento. No entanto, os valores obtidos nas provas de digestibilidad ileal podem variar de acordo às perdas de nitrog&ecird;nio endógeno do animal. Devido ao anterior, os valores de digestibilidad ileal podem ser expressados como aparentes ou verdadeiros. Na determinação do valor aparente não se têm em conta as perdas de nitrogênio endógeno, pelo que se subestima a quantidade de aminoácidos absorvidos durante a digestão. Ao tomar em conta o componente endógeno, a digestibilidad ileal se transforma em verdadeira, sendo esta o melhor parâmetro para representar os aminoácidos absorvidos no tracto digestivo. As perdas de nitrogênio endógeno foram classicamente definidas como o nitrogênio encontrado na digesta ou nas fezes de animais alimentados com dietas livres de nitrogênio. Tradicionalmente, as perdas de nitrogênio endógeno em porcos foram determinadas mediante os métodos de regressão linear ou pelo de oferecer dietas livres de nitrogênio. Atualmente se desenvolveram várias técnicas alternativas para estimar as perdas de nitrogênio endógeno em porcos que consomem dietas que contêm níveis variáveis de proteína. Cada uma destas técnicas possui limitações e todas requerem diferentes suposições. Baseados num crescente número de resultados obtidos com diferentes técnicas, as perdas de nitrogênio endógeno parecem estar altamente afetadas pelo método utilizado para sua determinação e por fatores como são peso corporal, consumo de matéria seca, e constituintes da dieta (fatores antinutricionales, fibra e proteína).
Palavras chaves: Fluxo de nitrogênio endógeno, porcos em crescimento, técnicas de quantificação.
Introducción
El valor nutricional de un ingrediente para animales puede ser determinado por su contenido de nutrientes disponibles, en especial por su aporte de aminoácidos (AA) y energía1. La digestibilidad ileal de los AA es uno de los factores más importantes para calificar la calidad de la dieta y la respuesta productiva de los animales; la cual, desde hace años, se ha determinado en animales monogástricos a través del muestreo del contenido ileal2. Sin embargo, los valores obtenidos en las pruebas de digestibilidad ileal pueden variar de acuerdo con las secreciones intestinales de nitrógeno o pérdidas de nitrógeno endógeno (PNE) del animal y por ello los valores pueden ser expresados como aparentes o verdaderos. En la determinación del valor aparente no se tienen en cuenta las PNE, por lo que se subestima la cantidad de AA absorbidos durante la digestión3. Al tomar en cuenta el componente endógeno, la digestibilidad ileal se transforma en verdadera, siendo ésta el mejor parámetro para representar los AA absorbidos en el tracto digestivo4. Por lo anterior, las PNE en cerdos en crecimiento deben ser determinadas y cuantificadas, ya que éstas pueden afectar la disponibilidad de nutrientes y la respuesta productiva en cerdos en crecimiento.
1. Pérdidas de nitrógeno endógeno
Las pérdidas de nitrógeno endógeno (PNE), son clásicamente definidas como el nitrógeno encontrado en la digesta o en las heces de animales alimentados con dietas libres de nitrógeno. Estos compuestos provienen principalmente de mucoproteínas5, enzimas pancreáticas e intestinales, saliva, secreciones biliares y gástricas, células descamadas de la mucosa intestinal, albúmina sérica, péptidos, aminoácidos (AA), amidas, aminas, glucosaminas6-8 y la proteína de origen bacteriano9, aunque esta última, estrictamente hablando, no es nitrógeno endógeno. Sin embargo, las PNE representan sólo una fracción del total de nitrógeno secretado dentro del intestino y su incremento está asociado principalmente a una mayor tasa de síntesis de proteína intestinal, y a una ineficiente tasa de síntesis proteica corporal. Aunado a lo anterior, estas pérdidas, aumentan el requerimiento de energía y aminoácidos en el animal.
2. Métodos para estimar las pérdidas de nitrógeno endógeno en cerdos
Muchos métodos han sido desarrollados para cuantificar el flujo de nitrógeno endógeno en cerdos en crecimiento. Estos métodos implican la utilización de dietas libres de nitrógeno, que contienen fuentes de proteínas que se asumen como 100% digestibles, y técnicas de regresión matemática. En los últimos tiempos, se han desarrollado técnicas que incluyen el uso de marcadores y ultrafiltrado de péptidos en el laboratorio. Entre los métodos más utilizados en experimentación se encuentran:
Dietas libres de nitrógeno: Esta técnica consiste en alimentar animales con una dieta libre de nitrógeno, midiendo el flujo de nitrógeno y AA en la digesta colectada en el íleon distal. Se asume que todo el nitrógeno (N) y los AA que aparecen en la digesta son de origen endógeno. La principal crítica a este método, es su naturaleza antifisiológica, porque disminuye el estímulo de la actividad normal del intestino, afecta el metabolismo de la proteína dietaria, reduce la secreción de compuestos nitrogenados en el lumen intestinal y subestima la concentración de PNE en íleon4,10. El resultado obtenido es un balance de proteína negativo11 en los animales evaluados, con un efecto poco representativo sobre las PNE en el íleon distal. Además, la ausencia de proteína en el alimento disminuye las secreciones endógenas y no hay factores que estimulen su producción o impidan su reabsorción por la pared intestinal12, por cuanto afecta la capacidad de absorción de los AA en el intestino delgado debido al contenido de proteína en la dieta13. El intestino delgado responde a una ingesta reducida de proteína con el incremento de la absorción intestinal de AA14. Adicionalmente, los constituyentes de la dieta, como la fibra y factores antinutricionales, los cuales están asociados a la proteína de la dieta, podrían aumentar las PNE15-18.
Regresión lineal: El método de regresión lineal (RL), consiste en alimentar al animal, incrementando las cantidades de N en la dieta (a un consumo constante de materia seca) y estimar las PNE por extrapolación del flujo ileal de AA a cero N consumido. Esto permite que las PNE sean determinadas bajo condiciones normales de alimentación proteica. La principal suposición con este método, es que el incremento en el contenido de N de la digesta ileal se debe a un incremento proporcional en la cantidad de N dietético indigestible. Se supone además, que no hay cambios en la calidad de N endógeno excretado19. La técnica de regresión es, potencialmente, el mejor estimador de las PNE, comparado con las dietas libres de nitrógeno20.
Técnica de Dilusión Isotópica: El método de Dilusión Isotópica (DI), implica la marcación del nitrógeno de la dieta con el isótopo pesado 15N13,21. Ha sido utilizado para marcar cualquier tipo de pool de N en el animal22 o en el N dietético23 y permite una diferenciación entre el nitrógeno endógeno y el N dietético no digerido. La cantidad de PNE puede ser subestimada por este método, ya que no tiene en cuenta las células de la mucosa secretadas por vía endógena, las cuales son sintetizadas a partir de AA dietéticos marcados en el lumen24. El N suministrado por los AA dietéticos puede ser absorbido e incorporado rápidamente en las proteínas corporales, lo cual complica la diferenciación del N no digerible y del NE25.
Técnica de la Homoarginina: Es una técnica que involucra la transformación de la lisina dietética a Homoarginina (HA), a través de un proceso de guanidación (una reacción con OMetilisourea)26,27. Esta técnica permite la diferenciación entre la lisina dietética (convertida en HA) y la lisina de la proteína del intestino y debe cumplir varios requisitos: a) la guanidación de la lisina en la proteína a ensayar debe ser uniforme o al menos completa; b) la guanidación no debe afectar la digestibilidad de la proteína; c) la HA no puede ser hidrolizada a urea o lisina en el intestino delgado; d) la HA debe ser absorbida y metabolizada como cualquier AA; e) la HA no puede reingresar al tracto digestivo; f) la presencia de HA en el plasma o intestino, no debe afectar las PNE28. Con este método, la lisina no es utilizada para la síntesis de proteína, por lo que debe ser usado poco tiempo, para evitar posibles deficiencias de lisina29. Además, con esta técnica la digestibilidad verdadera de los demás AA no es determinada directamente30.
Proteínas hidrolizadas enzimáticamente: Esta técnica permite el flujo de NE y de AA en el intestino delgado, para ser determinados en animales alimentados con dietas que contienen proteínas y AA como única fuente de N. Los animales son alimentados con una dieta en la que la única fuente de N es caseína hidrolizada enzimáticamente, la cual contiene aproximadamente 56% de AA libres y 41% de di-tripéptidos con pesos moleculares menores de 5000 Da31. Se asume entonces, que todo el NE está en fracciones con pesos moleculares mayores a 10000 Da. De esta manera, al separar pesos moleculares bajos (<5000 Da) y altos (>10000 Da) de compuestos nitrogenados en la digesta ileal, se puede hacer una identificación entre cualquier N exógeno no absorbido y cualquier N de origen endógeno32. La mayor restricción de esta técnica, es que no puede ser usada para estimar la secreción de NE donde haya una asociación directa entre la proteína y factores dietéticos, los cuales son conocidos por influenciar las secreciones endógenas y las PNE10. Este método puede ser aplicado en alimentos que contengan fuentes de proteína animal33.
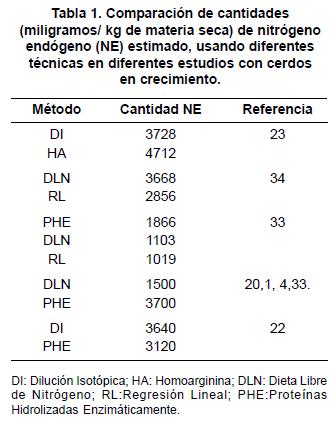
Comparación entre técnicas para determinar pérdidas de nitrógeno endógeno: En la literatura no se han reportado estudios en los que se comparen y evalúen todas las técnicas para determinar las PNE al mismo tiempo. Sin embargo, muchos estudios han comparado algunas de estas técnicas en cerdos (tabla 1), donde se ha observado que las PNE difieren entre métodos. El trabajo realizado por Roos et al23, indica que cuando se utiliza la técnica de HA se encuentran valores de PNE altos, comparados con los obtenidos con la técnica de DI. Esto puede deberse a que con la técnica de DI el N marcado puede ser absorbido e incorporado dentro de los diferentes ciclos de proteína de reciclaje intestinal, y por último, puede volver a ser secretado hacia el intestino delgado. Con la técnica de proteínas hidrolizadas enzimáticamente se obtienen mayores valores de PNE, al ser comparada con las técnicas de DLN y RL. Al parecer, la técnica de PHE induce mayores PNE, confrontadas al ofrecer dietas que se asumen como 100% digestibles. Lo anterior evidencia que los péptidos suministrados, ya sea como tales o derivados de la digestión natural de las proteínas dietarias, tienen influencia en las PNE del tracto digestivo1, 6. En una comparación directa, los resultados obtenidos con la técnica de PHE fueron similares a los obtenidos con la técnica de DI. Los resultados de estos estudios (tabla 1) sugieren que los métodos convencionales (DLN, RL y PHE) resultan en una subestimación de PNE en cerdos alimentados con dietas que contienen diferentes fuentes y niveles de proteínas. Además, estos resultados indican que los valores obtenidos de PNE son altamente variables34.
3. Factores que afectan las pérdidas de nitrógeno endógeno en cerdos.
Además de las técnicas anteriormente mencionadas, existen varios factores que pueden afectar los resultados y mediciones de las PNE en cerdos en crecimiento. Así mismo, la secreción y reabsorción de NE en el tracto gastrointestinal pueden estar influenciadas por muchos factores, entre los que se pueden encontrar:
Consumo de materia seca (MS) y peso corporal: Aparentemente, el efecto de la materia seca MS, consumida sobre las PNE está relacionado con el peso corporal del animal34, 36. Posiblemente, esto se deba a que los animales aumentan su consumo y comienzan a ser más pesados; se podría esperar que a mayor consumo de alimento, mayor es la secreción de las enzimas digestivas, las cuales contribuyen a aumentar el flujo de AA endógenos37. Liddle et al.38 han demostrado el efecto positivo que tiene la concentración de sustrato en la dieta (proteína, carbohidratos, grasas) sobre la secreción y actividad de algunas enzimas pancreáticas.
Procesamiento del alimento: Los alimentos molidos disminuyen el tamaño de la partícula (aumentando la superficie de contacto), la cual es expuesta a enzimas digestivas, mejorando la digestibilidad ileal de N y la utilización del alimento39. Los tratamientos térmicos son procesos que tienen efectos importantes en la digestibilidad ileal de N, ya que pueden destruir algunos factores antinutricionales contenidos en los alimentos, como las lectinas, factores inhibidores de proteasas, taninos, etc. Los tratamientos térmicos más comúnmente utilizados son: La extrusión, cocimiento y torrefacción40. El descascarillado, es otro tratamiento mecánico de gran importancia, debido a que disminuye el contenido de fibra y de taninos de algunas materias primas, localizadas principalmente, en la cáscara de leguminosas.
Factores antinutricionales: Estos han sido reportados para aumentar el flujo de NE en el íleon de cerdos. Los Inhibidores de Tripsina (IT)41, Lectinas42 y Taninos43 elevan la cantidad de N y NE que pasan al íleon terminal. Los taninos disminuyen la digestibilidad de N debido a que tienen la capacidad de unirse a las proteínas de la dieta, mediante la formación de puentes de hidrógeno e interacciones hidrofóbicas de sus grupos hidróxilos con los grupos carbonilo de las proteínas dietéticas, disminuyendo la actividad de la pepsina gástrica, la tripsina, la quimotripsina pancreática y las peptidasas intestinales del borde en cepillo. Por otro lado, los taninos actúan negativamente al cambiar la morfología de la mucosa gastrointestinal, disminuyendo la absorción transmembranal de nutrientes y al incrementar las secreciones endógenas de proteína44. Los IT interfieren directamente con la digestión del N de la dieta por unirse en forma irreversible a la tripsina y quimotripsina. Además, los IT estimulan la secreción compensatoria de estas enzimas pancreáticas y como resultado, se da un incremento en las PNE45. Los IT disminuyen la digestibilidad aparente de la cisteína, debido a que cerca del 50% de la cisteína en las leguminosas se localiza en los inhibidores de tripsina. Además, como las enzimas pancreáticas son ricas en este aminoácido, la formación de complejos irreversibles entre los IT con tripsina y quimotripsina, incrementan las pérdidas endógenas de cisteina45. Las lectinas son proteínas que tienen la capacidad de unirse a azúcares específicos, como la matriz de carbohidratos de la pared intestinal y del borde en cepillo, aumentando las pérdidas de NE. Las lectinas estimulan la proliferación de bacterias en el lumen, debido a una gran disponibilidad de nutrientes para las bacterias y una alta tasa de recambio celular, al tiempo que crece el número de sitios potenciales de unión para las bacterias en las células epiteliales42.
Contenido y calidad de la fibra dietaria: Las secreciones de jugos pancreáticos, bilis y moco, se incrementan cuando aumenta el consumo de pectinas y celulosa46. Las pectinas poseen propiedades de viscosidad y gelificación que disminuyen la digestión y absorción de nutrientes al reducir la mezcla de los contenidos intestinales e interferir con la unión de enzima-sustrato y formar una capa de agua estática que crea una barrera física para la absorción de nutrientes10. Estos componentes pueden interactuar con los AA liberados durante la hidrólisis de las proteínas y permitir que algunos escapen a la absorción en el intestino delgado (llegando al íleon distal), para luego pasar al intestino grueso47. Este componente también aumenta la secreción y pérdidas de mucinas e induce la proliferación de las células epiteliales y mucosas.
Fuentes y solubilidad de la proteína: Las fuentes de proteína animal son más digestibles que las fuentes de proteína vegetal, por cuanto los cambios en la composición de las proteínas de las plantas pueden causar cambios en la digestibilidad ileal de AA en cerdos48. Aunado a esto, la presencia de factores antinutricionales en alimentos vegetales puede disminuir la digestibilidad de los AA. La solubilidad de una proteína en un medio acuoso, como es el tracto gastrointestinal, es un factor determinante en la digestibilidad ileal aparente de AA, ya que propicia un medio único para la fácil unión entre enzima-sustrato49.
Cantidad y fuente de grasa dietética: La cantidad de grasa en la dieta afecta la digestión de proteínas en cerdos. En estudios con cerdos en crecimiento, se presentó un efecto lineal significativo a medida que aumentó la ingesta de grasa en la dieta. La grasa dietaria disminuye el tiempo de tránsito en el intestino delgado, lo cual permite un mayor tiempo de digestión y absorción de los AA antes de llegar al íleon terminal40. Un incremento en la grasa de la dieta retrasa el vaciado gástrico, lo que podría resultar en unabaja tasa de pasaje de la dieta en el intestino delgado, a su vez, en una alta tasa de digestibilidad de los AA. Esto es particularmente cierto, cuando el tiempo requerido para la digestión de las proteínas y la absorción de los AA es un factor limitante50.
Conclusiones
Esta revisión ha sido desarrollada con el fin de demostrar la importancia de las PNE en el íleon distal, y su efecto sobre la nutrición de cerdos en crecimiento. Muchos estudios citados en esta revisión se centran en las metodologías, para estimar las PNE, y en los efectos de los componentes dietarios sobre las PNE. El método utilizado para determinar las PNE tiene gran influencia sobre la cuantificación del flujo de nitrógeno en ingredientes utilizados en nutrición animal. Los métodos convencionales (DLN, RL y PHE) subestiman las PNE en cerdos alimentados con dietas que contienen diferentes fuentes y niveles de proteína. Estos estudios sugieren que la técnica de la Homoarginina (HA) podría ser una buena opción para la determinación de las PNE en una gran cantidad de alimentos. Basados en los resultados obtenidos con estas técnicas, las PNE parecen estar altamente afectadas por factores como el peso corporal, consumo de materia seca, fibra, contenido de factores antinutricionales y proteína (digestible) en la dieta. Una reducción en las PNE puede mejorar la eficiencia de la utilización de los aminoácidos y energía de la dieta en la producción de cerdos. Por tanto, los componentes específicos que causan estas pérdidas en dietas e ingredientes alimenticios para cerdos deben ser identificados.
Referencias
1. GUTIÉRREZ C., Lourdes, et al. Comparación de métodos para determinar el flujo ileal de aminoácidos endógenos y la digestibilidad verdadera de aminoácidos de harina de pescado suministrada a ratas. En: Revista Científica. Septiembre-octubre, 2005, vol.15, no.5, p.451-457. [ Links ]
2. JORGENSEN, H.; SAUER, W.C. and THACKER, P.A. Amino acid availabilities in soybean meal, sunflower meal, fish meal and meat and bone meal fed to growing pigs. En: Journal of Animal Science. March, 1984. vol. 58, no 4, p.596-604. [ Links ]
3. ARENTSON, R.A. and ZIMMERMAN, D.R. True digestibilities of amino acids and protein in pigs using C13 as a label to determine metabolic amino acid excretion. En: Journal of Animal Science. 1992. vol. 70, Suppl. 1, p.68. [ Links ]
4. DARRAGH, A.J.; MOUGHAN, P.J. and SMITH, W.C. The effect of amino acid and peptide alimentation on the determination of endogenous amino acid flow at the terminal ileum of the rat. En: Journal of the science of food and agriculture. 1990. vol. 51, no. 1, p.47-56. [ Links ]
5. CORRING, T. and JUNG, J. The amino acid composition of pig pancreatic juice. En: Nutrition Reports International. 1972. vol. 6, p.187-198. [ Links ]
6. SOUFFRANT, W.B. Endogenous nitrogen losses during digestion in pigs. En: International Symposium on Digestive Physiology in Pigs (5: 24-26, april: Pudoc, Wageningen, The Netherlands). Memorys. PUDOC, Wageningen Doorwerth, The Netherlands: Verstegen. M.W.A; Huisman, J.and den Hartog, L.A, 1991. p. 147-166. [ Links ]
7. MOUGHAN, Paul J., et al. Evaluation of the isotope dilution technique for determining ileal endogenous nitrogen excretion in the rat. En: Journal of the Science of Food and Agriculture. 1992. vol. 58, no. 2, p. 165-172. [ Links ]
8. JANSMAN, A. J., et al. Evaluation through literature data of the amount and amino acid composition of basal endogenous crude protein at the terminal ileum of pigs. En: Animal Feed Science and Technology. 2002. vol. 98, no. 1-2, p.49-60. [ Links ]
9. CAINE, W.R., et al. Apparent ileal digestibilities of amino acids in newly weaned pigs fed diets with protease-treated soybean meal. En: Journal of Animal Science. 1997. vol. 75, no. 11p.2962-2969. [ Links ]
10. NYACHOTI, C.M., et al. Significance of endogenous gut nitrogen losses in the nutrition of growing pigs: A review. En: Canadian Journal of Animal Science. March, 1997. vol. 77, no. 1, p.149-163. [ Links ]
11. OTTO, E.R., et al. Nitrogen balance and ileal amino acid digestibility in growing pigs fed diets reduced in protein concentration. En: Journal of animal Science. July, 2003. vol. 81, no. 7, p.1743-1753. [ Links ]
12. LETERME, Pascal, et al. Effect of oral and parenteral N nutrition vs N-free nutrition on the endogenous amino acid flow at the ileum of the pig. En: Journal of the Science of Food and Agriculture. 1996. vol. 71, no. 2, p. 265-272. [ Links ]
13. De LANGE, C.F.M. et al. The effect of protein status of the pig on the recovery and amino acid composition of endogenous protein in digesta collected from the distal ileum. En: Journal of Animal Science. 1989. vol. 67, p.755-762. [ Links ]
14. CHRISTENSEN, H.N. The regulation of amino acid and sugar absorption by diet. En: Nutrition Reviews. July. 1984, vol. 42, no. 7, p.237-242. [ Links ]
15. SKILTON, Grant A.; MOUGHAN, Paul J. and SMITH, William C. Determination of endogenous amino acids flow at terminal ileum of the rat. En: Journal of the Science of Food and Agriculture. 1988. vol. 44, no. 3, p. 227-235. [ Links ]
16. MARISCAL, G.; REIS, T.C. e PARRA, J.E. Estimação dos coeficientes de digestibilidade ileal aparente das proteínas e aminoácidos da torta de canola em leitões recém-desmamados. En: Revista Colombiana de Ciencias Pecuarias. 2008. vol. 21, no. 2, p.201-209. [ Links ]
17. NYACHOTI, C.M.; ATKINSON, J.L. and Leeson S. Sorghum tannins: a review. En: World's Poultry Science Journal. 1997. vol. 53, p.5-21. [ Links ]
18. REIS, T.C.; MARISCAL, G. y AGUILERA, A. Efecto de diferentes cereales en dietas de iniciación para lechones sobre la digestibilidad sobre la digestibilidad de los nutrimentos y la referencia alimentaria: En: Veterinaria. México. Enero-marzo, 2005. vol. 36, no. 1, p.11-24. [ Links ]
19. HOOGKINSON, S.M., et al. Effect peptide concentration on endogenous ileal amino acids loss in the growing pig. En: British Journal of Nutrition. 2000. vol. 83, no. 4, p.421-430. [ Links ]
20. FAN, M.Z.; SAUER, W.C. and MCBURNEY, M.I. Estimation by regression analysis of endogenous amino acids levels in digesta collected from the distal ileum of pigs. En: Journal of Animal Science. 1995. vol. 73, no. 8, p.2319-2328. [ Links ]
21. KRAWIELITZKI, K., et al. Estimation of N absorption, secretion, and reabsorption in different intestinal sections of growing pigs using the 15N isotope dilution method. En: International Symposium on Digestive Physiology in Pigs (6: 4-6 october: Bad Doberan, Danmar). Memorys. Bad Doberan, Danmar: W,B. Souffrant and H. Hagemeister, Proc, 1994. p.79-82. [ Links ]
22. SCHULZE, H., et al. Effect of level of dietary neutral detergent fiber on ileal apparent digestibility and ileal nitrogen losses in pigs. En: Journal of Animal Science. 1994. vol. 72, no. 9, p.2362-2368. [ Links ]
23. ROOS, Nils; PFEUFFER, María and HAGEMEISTER, Hans. Labelling with 15N as compared with homoarginine suggest a lower prececal digestibility of casein in pigs. En: Journal of Nutrition. December, 1994. vol. 124, no. 12, p.2404-2409. [ Links ]
24. DE LANGE, C.M.M., et al. 15N-Leucine 15NIsoleucine isotope dilution techniques versus the 15N-Isotope dilution technique for determining the recovery of endogenous protein and amino acids in digesta collected from the distal ileum in pigs. En: Journal of Animal Science. 1992. vol. 70, no. 6, p.1848-1858. [ Links ]
25. TAMMINGA, S., et al. The nutritional significance of endogenous N-losses along the gastrointestinal tract of farms animals. En: Archives of Animal Nutrition. 1992. vol. 48, no, 1-2, p.9-22. [ Links ]
26. HAGEMEISTER, H. and ERBERSDOBLER, H. Chemical labelling of dietary protein by transformation of lysine to homoarginine: a new technique to follow intestinal digestion and absorption. En: The Proceedings of the Nutrition Society. 1985. vol. 44 p.133. [ Links ]
27. HODGKINSON, S.M.; SOUFFRANT, W.B. and MOUGHAN, P.J. Comparison of the enzymehydrolyzed casein, guanidination, and isotope dilution methods for determining ileal endogenous protein flow in the growing rat and pig. En: Journal of Animal Science. 2003. vol. 81, no. 10, p.2525-2534. [ Links ]
28. DRESCHER, H., et al. Guanidination (homoarginine- labelling) of proteins does affect prececal 15N recovery in pigs. En: International Symposium on Digestive Physiology in Pigs (6: 4-6 october: Bad Doberan, Danmar). Memorys. Bad Doberan, Danmar: W,B. Souffrant and H. Hagemeister, Proc, 1994. p.64-66. [ Links ]
29. TEWS, Jean K. and HARPER, Alfred E. Induction in rats of lysine imbalance by dietary homoarginine. En: Journal of Nutrition. 1986, vol. 116, no. 10, p.1910-1921. [ Links ]
30. MARTY, B.J.; CHAVEZ, E.R. and DE LANGE C.F.M. Recovery of amino acids at the distal ileum for determining apparent and true amino acid digestibilities in growing pigs fed various heatprocessed full-fat soybean bean products. En: Journal of Animal Science. 1994. vol. 72, p.2029-2037. [ Links ]
31. LETERME, P., et al. Effect of oral and parenteral N nutrition vs N-free nutrition on the endogenous amino acid flows at ileum of the pig. En: International Symposium on Digestive Physiology in Pigs (6: 4-6 october: Bad Doberan, Danmar). Memorys. Bad Doberan, Danmar: W,B. Souffrant and H. Hagemeister, Proc, 1994. p.60-63. [ Links ]
32. MOUGHAN, Paul J. and RUTHERFURD, Shane M. Endogenous flow of total lysine and other amino acids at the distal ileum of the protein- or peptide-fed rat: The chemical labeling of gelatin protein by transformation of lysine to homoarginine. En: Journal of the Science of Food and Agriculture. 1990. vol. 52, p.179-192. [ Links ]
33. DONKOH, A.; MOUGHAN, P.J. and MORE, P.C.H. Comparison of methods to determine the endogenous amino acids flow at terminal ileum of growing rats. En: Journal of the Science of Food and Agriculture. 1995. vol. 67, no. 3, p.359-366. [ Links ]
34. MARISCAL-LANDIN, Gerardo, et al. Endogenous amino nitrogen collected from pigs with end-toend ileorectal anastomosis is affected by the method of estimation and altered by dietary fiber. En: Journal of Nutrition. 1995. vol. 125, no. 1, p.136-146. [ Links ]
35. BUTTS, Christine A.; MOUGHAN, P.J. and SMITH, William C. Endogenous amino acid flow at the terminal ileum of the rat determined under conditions of peptide alimentation. En: Journal of the Science of Food and Agriculture. 1991. vol. 55, no, 2, p.175-187. [ Links ]
36. HESS, V. and SÉVE, B. Effects of body weight and feed intake level on basal ileal endogenous losses in growing pigs. En: Journal of Animal Science. 1999. vol. 77, no. 12, p.3281-3288. [ Links ]
37. CERVANTES RAMIREZ, Miguel, et al. Pérdida de aminoácidos endógenos en cerdos con niveles variables de consumo de alimento. En: Agrociencia. Noviembre-diciembre, 2000. vol. 34, no. 6, p. 707-715. [ Links ]
38. LIDLLE, A.R. Proteins but not amino acids, carbohydrates, or fats stimulate cholecystokinin secretion in the rat. En: American Journal of Physiology. August, 1986. vol. 251, no. 2, parte 1, p.G243-G248. [ Links ]
39. OWSLEY, Walter F.; KNABE D.D. and TANKSLEY, T.D. Effect of sorghum particle size on digestibility of nutrients at terminal ileum and over the total digestive tract of growing-finishing pigs. En Journal of Animal Science. March, 1981. vol. 52, no. 3, p.557-566. [ Links ]
40. LI, S. and SAUER, William C. The effect of dietary fat content on amino acid digestibility in young pigs. En: Journal of Animal Science. 1994. vol. 72, no. 7, p.1737-1743. [ Links ]
41. BARTH, Christian A., et al. Soybean trypsin inhibitor(s) reduce absorption of exogenous and increase loss of endogenous protein in miniature pigs. En: Journal of Nutrition. December, 1993. vol. 123, no. 12, p.2195-2200. [ Links ]
42. GRANT, George and DRIESSCHE, Edilbert. Legume lectins: physiochemical and nutritional properties. En: Recent advances of research in antinutritional factors in legume seeds. 2 ed. Wageningen, The Netherlands, 1993. p.219-234. [ Links ]
43. JANSMAN, A.J.M., et al. Effects of hulls of faba beans (Vicia faba) with low or high content of condensed tannins on the apparent ileal and fecal digestibility of nutrients and excretion of endogenous protein in ileal digesta and feces of pigs of pigs. En: Journal of Animal Science. 1975. vol. 73, no. 1, p.118-127. [ Links ]
44. JANSMAN, A.J.M. Tannins in feedstuffs for single- stomached animals. En: Nutrition Research Reviews. 1993. vol. 6, p.209-236. [ Links ]
45. HUISMAN, J. and JANSMAN, A.J.M. Dietary effects and some analytical aspects of nutritional factors in peas (Pisum sativum), beans (Phaseolus vulgaris) and soybean (Glycine max L.) in monogastric farm animals. A literature review. En: Nutrition Abastract Review. 1991. vol. 61, p.901-921. [ Links ]
46. MOSENTHIN, Rainer; SAUER, William C. and AHRENS, Frank. Dietary pectin's effect on ileal and fecal amino acids digestibility and exocrine pancreatic secretions in growing pigs. En: Journal of Nutrition. August, 1994. vol. 124, no. 8, p.1222-1229. [ Links ]
47. LOW, A.G. Nutrient absorption in pigs. En: Journal of the Science of Food and Agricultura. November, 1980. vol. 31, no. 11, p.1087-1130. [ Links ]
48. SOHN, K.S., et al. Improved soybean proteins sources for early-weaned pigs: II. Effects on ileal amino acids digestibility. En: Journal of Animal Science. 1994. vol. 72, no. 3, p.631-637. [ Links ]
49. JUST, A.; JORGENSEN, H. and FERNANDEZ, J.A. The digestive capacity of the caecum-colon and the value of the nitrogen absorbed from the hindgut for proteins syntesis in pigs. En: British Journal of Nutrition. July, 1981. vol. 46, no. 1, p.209-219. [ Links ]
50. INBEAH, M., and SAUER, W.C. The effect de dietary level of fat on amino acid digestibilities in soybean meal and canola meal and rate of passage in growing pigs. En: Livestock Production Science. 1991. vol. 29, p.227-239. [ Links ]

