










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Lasallista de Investigación
Print version ISSN 1794-4449
Rev. Lasallista Investig. vol.7 no.2 Caldas July/Dec. 2010
Artículo original / Original article / Artigo original
Efecto de la transición adenina/guanina del gen de la prolactina bovina sobre características de importancia en producción lechera
Effect of adenina/guanina transition in prolactine gen on important milk traits
Efeito de transição de adenina/guanina do gene da prolactina bovinos em características de importância na produção leiteira
* Profesor auxiliar, Departamento de Producción Animal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, sede Medellín. jjecheve@unal.edu.co. Grupo BIOGEM
** Profesor asistente, Facultad de Ciencias, Universidad Nacional de Colombia, sede Medellín
*** Ingeniera biológica, Universidad Nacional de Colombia, sede Medellín
Correspondencia: José Julián Echeverri Zuluaga. e-mail: jjecheve@unal.edu.co.
Artículo recibido: 19/12/2009; artículo aprobado: 3/12/2010
Resumen
Entre las características de mayor importancia en ganado lechero, están la producción de leche y la calidad de la misma. Entre las hormonas de importancia en estos procesos, están la hormona del crecimiento y la prolactina. La prolactina cumple diversas funciones; por esta razón se han buscado secuencias polimórficas en el gen que codifica dicha proteína con el uso de técnicas moleculares que permiten detectar polimorfismos para regiones específicas del DNA; esto con el fin de utilizarlos para construir mapas genéticos y evaluar su efecto sobre la expresión de características. El objetivo principal fue determinar las frecuencias alélicas y genotípicas del gen de la prolactina y la asociación entre los polimorfismos de este y algunas características productivas. La determinación de los genotipos se realizó mediante PCR-RFLP, con DNA extraído de sangre periférica por salting out. Se analizaron 470 lactancias correspondientes a 165 vacas Holstein, a las cuales se les hizo seguimiento para producción y composición de la leche durante, mínimo, una lactancia completa. El alelo más frecuentes para el gen PRL fue PRLA (0.91); el genotipo de mayor frecuencia fue AA (0.83) mientras el de menos frecuente fue PRLBB (0.02); producción de leche, porcentaje de proteína y cantidad de proteína tuvieron diferencia estadísticamente significativa; las demás asociaciones no tuvieron significancia. Estos resultados coinciden con los de otros investigadores; por esto se debe buscar a través de la selección los individuos con genotipos favorables y utilizarlos en programas de mejoramiento genético.
Palabras clave: marcadores moleculares, PCR, proteína.
Abstract
Introduction. Among the most important characteristics of Dairy cattle, milk production and quality are remarked. Among the important hormones for these processes, growth hormone and prolactin are highlighted. Prolactin has many functions; therefore polymorphic sequences have been searched in the gen that codifies that protein, by the use of molecular techniques to detect polymorphisms in specific DNA regions. This is useful to make genetic maps and evaluate the effect on the characteristic's expression. Objective. To determine allelic and phenotypic frequencies of the prolactin gen and the association between its polymorphisms and some production characteristics. Materials and methods. The determination of genotypes was made by means of PCRRFLP, with DNA extracted from peripheral blood by salting out. 470 lactations from 165 Holstein cows were analyzed, and a follow–up to their milk production and composition was made during a minimum a complete lactation. Results. The most frequent allele for the PRL gen was PRLA (0.91); the most frequent genotype was AA (0.83) while the less frequent was PRLBB (0.02); Milk production, protein percentage and protein quantity had a significant statistic difference, and the others had no significance. Conclusion. These results match those achieved by other researchers. Therefore, individuals with favorable phenotypes must be found through selection and they must be used in genetic increase programs.
Key words: molecular markers, PCR, protein.
Resumo
Introdução. Entre as características de maior importância em gado leiteiro, estão a produção de leite e a qualidade da mesma. Entre os hormônios de importância nestes processos, estão o hormônio do crescimento e a prolactina. A prolactina cumpre diversas funções; por esta razão se procuraram seqüências polimórficas no gene que codifica dita proteína com o uso de técnicas moleculares que permitem detectar polimorfismos para regiões específicas do DNA; isto com o fim de utilizá-los para construir mapas genéticos e avaliar seu efeito sobre a expressão de características. Objetivo. Determinar as freqüências alélicas e genotípicas do gene da prolactina e a associação entre os polimorfismos deste e algumas características produtivas. Materiais e métodos. A determinação dos genótipos se realizou mediante PCR-RFLP, com DNA extraído de sangue periférico por salting out. Analisaram-se 470 latências correspondentes a 165 vacas Holstein, às quais se lhes fez seguimento para produção e composição do leite durante, mínimo, uma latência completa. Resultados. O alelo mais frequentes para o gene PRL foi PRLA (0.91); o genótipo de maior freqüência foi AA (0.83) enquanto o de menos frequente foi PRLBB (0.02); produção de leite, percentagem de proteína e quantidade de proteína tiveram diferença estatisticamente significativa; as demais associações não tiveram significância. Conclusão. Estes resultados coincidem com os de outros pesquisadores; por isto se deve procurar através da seleção os indivíduos com genótipos favoráveis e utilizá-los em programas de melhoramento genético.
Palavras importantes: marcadores moleculares, PCR, proteína.
Introducción
Desde la aparición de la ganadería como actividad económica, el ganado lechero se ha visto sometido a selección fenotípica, tendente a mantener los caracteres de mayor producción. Las principales limitantes de la selección fenotípica son: 1. El tiempo que se requiere para determinar las características de la progenie y el impacto de las habilidades predichas de transmisión (por sus siglas en inglés PTA) de los toros, y 2. Los caracteres seleccionados pueden ser en ocasiones ambiguos, debido al impacto de las influencias ambientales y fisiológicas que los afectan1. El gen de la prolactina bovina está localizado en el cromosoma 23; está conformado por 5 exones y 4 intrones y codifica para una proteína de 229 aminoácidos con un peso molecular aproximado de 23 KDa2. Esta hormona es considerada como la más versátil en términos de sus funciones biológicas e inmunológicas3,4,5.
El desarrollo y la diferenciación de la glándula mamaria ocurren principalmente durante la preñez. Hembras homocigóticas (-/-) para una mutación del gen del receptor para PRL son estériles debido a un fallo completo en la implantación del blastocisto. En ratones tratados con progesterona, la preñez se logra recuperar pero se observa un notable subdesarrollo de la glándula mamaria6. En bovinos, la prolactina forma parte del complejo de hormonas que intervienen en el desarrollo mamario (mamogénesis), y es considerada como el factor primario requerido para el crecimiento y la diferenciación celular de este tejido7,8.
Actualmente se han descrito polimorfismos genéticos en algunos genes, relacionados con la producción y calidad composicional de la leche como Kappa-caseina, Beta-lactoglobulina, hormona de crecimiento y prolactina.9 Para la prolactina se ha reportado un polimorfismo bialélico en la posición +7544, en donde el alelo A presenta una guanina (G) y el alelo B presenta una adenina (A); esta transición genera una mutación silenciosa que codifica para el aminoácido valina en la posición 1032.
El objetivo principal de este trabajo fue determinar las frecuencias alélicas y genotípicas de los genes de la prolactina y hormona del crecimiento bovinas, así como explorar la asociación entre los polimorfismos de estos genes y algunas características productivas como producción y composición de la leche en 8 fincas ubicadas en diferentes municipios del departamento de Antioquia.
Materiales y métodos
Se seleccionaron 165 ejemplares de una población de ganado productor de leche en 8 hatos ubicados en varios municipios del departamento de Antioquia. De cada uno de los individuos se colectaron 5 ml de sangre periférica. Para la obtención del DNA de sangre periférica se utilizó el método modificado descrito por Miller y colaboradores en 198810. Se realizó una amplificación por PCR individual para cada región específica en un volumen final de 50 µL que contiene buffer PCR (1.0 -1.5 mM de MgCl2, 50 mM de KCl, 10 mM de Tris-HCl, pH de 8.3), 100-300 nM de cebadores; 2.0 mM de cada dNTPs, 0.5 unidades de taq polimerasa y DNA de sangre de bovino.
Los cebadores utilizados para amplificar las regiones específicas de los genes de la PRL (+7474 a +7627 Acceso AF426315)19 que contenían los polimorfismos fueron descritos por Dybus en 20022 y son los siguientes:
PRL:
F. 5'-CGAGTCCTTATGAGCTTGATTCTT-3'
R 5' GCCTTCCAGAAGTCGTTTGTT TTC-3'
La PCR se realizó en un termociclador (Biometra®). Las condiciones de la PCR para la amplificación de la región específica del gen de PRL fueron descritas por Dybus en 20022. Luego, los fragmentos amplificados por PCR de los genes de bGH (329 pb) y PRL (156 pb) fueron sometidos a digestión con endonucleasas de restricción para determinar los alelos. Para la determinación de los alelos de los genes PRL, se tomaron 13µl del producto de la PCR, se sometieron a digestión con 5 unidades de la enzima de restricción RsaI, y se incubaron a 37°C durante 12 horas. Para el gen de PRL el producto de PCR tiene un tamaño de 156 pb; al someterlo a digestión con la enzima RsaI, el alelo A no es cortado, mientras que el alelo B es digerido y genera los fragmentos de 82 y 74 pb2. Los productos de la amplificación y digestión se resolvieron por electroforesis en geles de agarosa al 4% (Amresco®).
Análisis estadístico
La frecuencia de los diferentes alelos se realizó con el método descrito por (Hartl, 2000)11. La frecuencia total (p) de los alelos 1 en la población es:
p= Fa/a +1/2 Fa/b, Frecuencia total (q) de los alelos 2 en la población es:
q= Fb/b + 1/2Fa/b, donde; F a/a = Homocigótico y F a/b = Heterocigótico.
Determinación de la asociación genotípica con características productivas
Se realizó un análisis de varianza basado en las fuentes de variación conocidas para cada una de las variables dependientes (producción de leche, porcentaje de grasa y porcentaje de proteína).
El modelo de efectos fijos llevado a cabo fue el siguiente:
Yijklmnop = µ + Gi + Gpm + Hj + NPl + G*Gp +Eijklmnop
Donde Yijklmnop = Característica analizada (producción láctea, porcentaje de proteína, porcentaje de grasa), µ = Media para la característica, Hj = Efecto del hato (j = 1....8), NPl Efecto del número de parto (l = 1....8), Gi = Efecto fijo del genotipo para GH(i= 1...3), Gpm = Efecto fijo del genotipo para PRLGH(i= 1...3), Gi* Gpm= Efecto de la interacción entre genotipos, Eijklmp = Error experimental
Los factores de corrección para los efectos medioambientales se estimaron mediante la utilización de el método de mínimos cuadrados, descrito por Harvey, y los análisis estadísticos restantes, con procedimientos del paquete estadístico Stat Graphics plus (versión 4.0).
Resultados
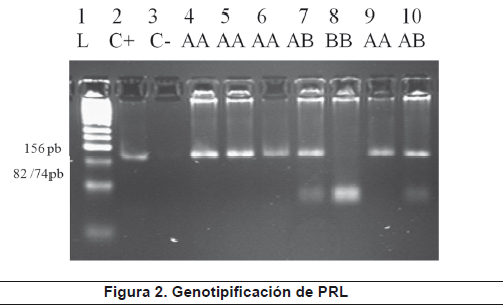
El fragmento amplificado de 156 pb (figura 1) que corresponde a la región del gen PRL que presenta el polimorfismo fue sometido a digestión con la enzima RsaI para la determinación de los genotipos. Los fragmentos de restricción de DNA específicos para cada genotipo fueron: 82 y 74 pb para el genotipo BB; 156, 82 y 74 para el genotipo AB y 156 pb (no digerido) para el genotipo AA (figura 2).
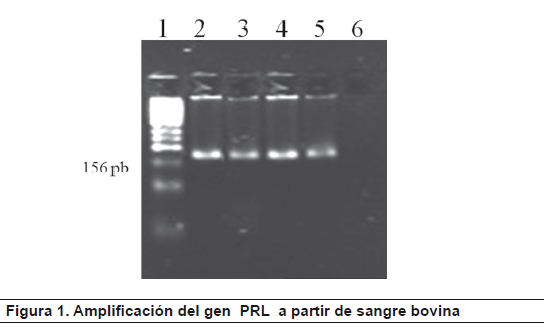
Electroforesis en gel de agarosa al 4.0%. La figura presenta el fragmento amplificado de 156pb correspondiente al gen de PRL, en algunos de los individuos analizados (carriles 2, 3 4 y 5), y el carril 6 control negativo (sin DNA). El carril 1 fue un marcador de peso molecular de 50pb.
Electroforesis en gel de agarosa al 4.0%. El carril numero 1 corresponde a marcador de peso molecular de 50 pb, carril 2 corresponde al control positivo de amplificación ( C+), carril 3 control negativo de amplificación, los carriles 4, 5 y 6 corresponde a amplicones de tres individuos analizados antes de digestión con RsaI, el carril 7 presenta 2 bandas de 156pb, 82/74 pb correspondientes a los fragmentos de restricción generados por digestión con RsaI para el genotipo PRL AB, El carril 8 exhibe una banda de 82/74 pb que corresponde al genotipo PRL BB, el carril 9 muestra el fragmento de 156 pb correspondiente al genotipo PRL AA, carril 10 control positivo de digestión.
Frecuencias alélicas y genotípicas
La frecuencia alélica fue de 0.91 para el alelo PRL A y de 0.09 para el alelo PRL B (tabla 1). El genotipo PRL AA fue el más frecuente en todos los hatos estudiados (0.73-0.95), seguido por el PRL AB (0.0-0.26); el de menor frecuencia fue PRL BB (0.0-0.02) (tabla 2). Este último genotipo fue hallado sólo en 2 de los hatos estudiados.
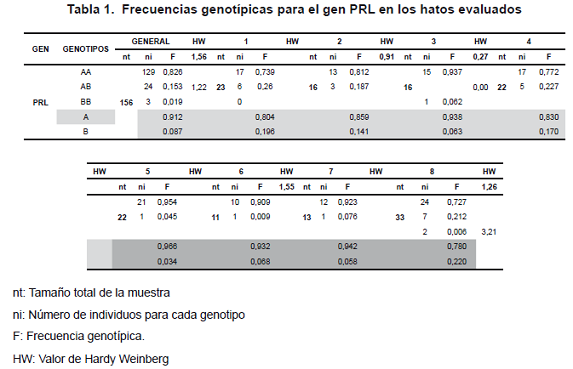
En la tabla se observa la distribución de los alelos y genotipos en cada uno de los hatos; ninguno de los hatos mostró equilibrio de hardy weinberg; igual sucedió con la población total. La fórmula de equilibrio estuvo en algunos casos por debajo y en otros por encima de 2, lo que indica que estas poblaciones han estado sometidas a selección y que ha habido una pérdida de la heterocigocidad.
En la mayoría de los hatos del país se ha efectuado una selección de individuos con base en su desempeño fenotípico y, además, se ha importado semen de toros seleccionados para características como producción y composición de la leche; por esta razón las frecuencias genotípicas han ido aumentando hacia la homocigocidad, disminuyendo la variabilidad genética y genotípica.
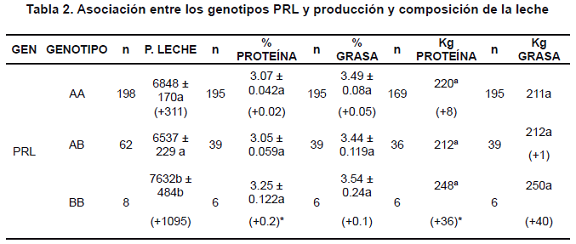
Efecto del genotipo de PRL sobre características productivas de ganado lechero Producción de leche. De acuerdo con el análisis realizado, las vacas con genotipo PRL BB produjeron 1095 L de leche más por lactancia que las vacas PRL AB; de la misma forma, los individuos PRL AA fueron superiores en 311 L a los PRL AB que presentaron el menor rendimiento (tabla 3). Estas diferencias fueron estadísticamente significativa (p<0.05), y sus medias fueron comparadas usando el test de Duncan, con un 95% de confianza.
Porcentaje de proteína de la leche. Los individuos con genotipo PRL BB produjeron leche con 0.2% y 0.18% más de proteína cuando se compararon con los PRL AB y PRL AA, respectivamente. Esta diferencia fue estadísticamente significativa al realizar la comparación de medias por el test de Duncan con un 95% de confianza (tabla 3) e implica un ingreso adicional para el productor de $150.000 por lactancia.
Producción de proteína láctea. Los individuos con genotipo PRL BB produjeron 36 y 28 Kg de proteína más por lactancia con respecto a los PRL AB y PRL AA, respectivamente. Esta diferencia fue estadísticamente significativa al realizar la comparación de medias por el test de Duncan con un 95% de confianza (Tabla 2).
Porcentaje de grasa de la leche. No se encontró diferencia significativa entre los diferentes genotipos (tabla 2).
Producción de grasa de la leche. Las vacas con genotipo PRL BB produjeron 38 y 39 Kg de grasa más por lactancia con respecto a los individuos con genotipo PRL AB y PRL AA, respectivamente. Esta diferencia no fue estadísticamente significativa (tabla 2).
La tabla presenta el número de lactancias evaluadas para cada característica, dentro de cada recuadro aparece la media de mínimos cuadrados ± Error estándar. El valor entre paréntesis representa la superioridad de cada genotipo con respecto al de peor desempeño. Los kg de proteína y grasa se calcularon con base en las lactancias que tenían información para las características completas. Letras diferentes en la misma columna y dentro de las filas correspondientes al mismo genotipo indican diferencia significativa p<0.05. *Diferencia significativa entre las medias del mismo grupo por el test de Duncan, 95% de confianza.
Discusión
Frecuencias alélicas y genotípicas
Las frecuencias alélicas y genotipicas son similares a las reportadas por Mitra y colaboradores 199520, Chung y colaboradores 199612, Udina y colaboradores 200113 (0.91), y Brym y colaboradores14 2005 (0.9), mayor a la reportada por Dybus 20022 (0.86), Golijow y colaboradores 199915 (0.87), y menor a la reportada por Chrenek y colaboradores 199816 (0.95), Ripoli y colaboradores en 199917 (0.98).
En la mayoría de los hatos del país se ha efectuado una selección de individuos con base en su desempeño fenotípico y, además, se ha importado semen de toros seleccionados para características como producción y composición de la leche; por esta razón las frecuencias genotípicas han ido aumentando hacia la homocigocidad, disminuyendo la variabilidad genética y genotípica.
Producción de leche. Los resultados difieren de los reportados por Dybus 20022, Chung y colaboradores 199612, quienes hallaron mejor desempeño en producción de leche para los individuos PRL AA. Este resultado, en particular, muestra superioridad del genotipo menos frecuente (PRL BB); el genotipo PRL AA es el segundo en desempeño; es importante tener en cuenta esto en el momento de definir la orientación genética de los hatos hacia uno de los genotipos, ya que el mejor genotipo para esta característica tuvo muy baja representatividad en el tamaño total de la muestra.
Porcentaje de proteína de la leche. Los resultados hallados difieren con los reportados por Dybus 20022 y de Pawel y colaboradores 200518, quienes encontraron el genotipo PRL AA más frecuente en vacas productoras de leche con mayor contenido de proteína.
Producción de proteína láctea. Los resultados encontrados difieren con los reportados por Dybus 20022 y de Pawel y colaboradores 200518, quienes encontraron el genotipo PRL AA más frecuente en vacas que produjeron mayor cantidad de proteína por lactancia. En las condiciones del mercado nacional es más importante el porcentaje de proteína de la leche que los Kg totales de esta, por tanto, es este el parámetro más importante para la selección de individuos.
Porcentaje de grasa de la leche. Para este parámetro los resultados coinciden con los descritos por Dybus 20022 y Pawel 200518, pero difieren de lo reportado por Chung y colaboradores 199612, quienes encontraron que los individuos con genotipos PRL AA producían leche con mayor contenido de grasa que los individuos PRL BB.
Producción de grasa de la leche. Estos resultados difieren con los reportados por Dybus 20022 y de Pawel y colaboradores 200518, quienes encontraron el genotipo PRL AA más frecuente en vacas que produjeron mayor cantidad de grasa por lactancia.
El ganadero debe orientar el mejoramiento de su hato hacia la producción de leche con altos contenidos de sólidos. Para cumplir con este objetivo el ganadero tiene diferentes herramientas. Esta investigación se presenta como uno de los primeros pasos en el interés de utilizar los marcadores genéticos como herramientas de selección en hatos lecheros.
La baja frecuencia de algunos de los genotipos encontrados en esta investigación genera una gran dificultad para determinar el grado de asociación que tiene cada uno de estos con características como producción de leche, grasa y proteína. Se requiere redoblar esfuerzos por parte de los investigadores para aumentar el tamaño muestral y conseguir resultados que generen mayor confiabilidad en cuanto a los marcadores genéticos utilizados en los programas de mejoramiento genético.
Referencias
1. GARY, W. R. Use of genetic marker for improving dairy cattle. Pennsylvania: Universidad Park, 1993. [ Links ]
2. DYBUS, A. Associations of growth (GH) and Prolactin (PRL) genes polymorphisms with milk production traits in Polish Black and White cattle. En: Animal Science Papers and Reports. 2002. Vol. 20, no. 4, p. 203-212. [ Links ]
3. BOLE-FEYSOT, Christine; et al. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. En: Endocrine Reviews. 1998. Vol. 19, no. 3, p. 225-268. [ Links ]
4. WICKS, Joan R. and BROOKS, Charles L. Biological activity of phosphorylated and dephosphorylated bovine prolactin. En: Molecular and Cellular Endocrinology. 1995. Vol. 112, no. 2, p. 223-229. [ Links ]
5. FREEMAN, Marc; et al. Prolactin: Structure, Function, and Regulation of secretion. En: Physiological Review. 2000. Vol. 80, no. 4, p. 1523-1631. [ Links ]
6. KELLY, Paul A.; et al. The role of prolactin and growth hormone in mammary gland development. En: Molecular and Cellular Endocrinology. 2002. Vol. 197, no. 1-2, p. 127-131. [ Links ]
7. KNIGHT, C. H. Overeview of prolactin'srole in farm animal lactation. En: Livestock Production Science. 2001. Vol. 70, no. 1-2, p. 87-93. [ Links ]
8. CHUGHTAI, Naila; et al. Prolactin Induces SHP-2 Association with Stat5, Nuclear Traslocation, and Binding to the B-Caseina Gene Promoter in Mammary Cells. En: The Journals of Biological Chemistry. 2002. Vol. 227, no. 34, p. 31107-31114. [ Links ]
9. JACOB, C.; et al. Nuevas estrategias en mejora genética. En: Frisona española. 1999. No. 112. [ Links ]
10. MILLER, S. A.; DYKES, D.D. and POLESKY, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. En: Nucleic Acids Research. 1988. Vol. 16, no. 3, p. 1215. [ Links ]
11. HARTL, Daniel L. A primer of populations Genetics. 3th ed. Sunderland, Mássachussets: Sinauer Associates Inc. Publishiers, 2000. p. 26-31. [ Links ]
12. CHUNG, E. R.; RHIM, T. J. and HAN S. K. Associations between PCR-RFLP marker of growth hormone and prolactin genes and production traits in dairy cattle. En: Korean Journal of Animal Science. 1996. Vol. 38, p. 321-336. [ Links ]
13. UDINA, I.G.; et al. Polymorphism of bivine prolactin gene: Microsatellites, PCR-RFLP. En: Russian Journal of Genetics. 2001. Vol. 37, no. 4, p. 407-411. [ Links ]
14. BRYM, Pawel, KAMINSKI, Stanislaw and WOJCIK, Elzbieta. Nucleotide sequence polymorphism within exon 4 of the bovine prolactin gene and its associations with performance traits. En: Journal Applied Genetics. 2005. Vol. 45, no. 2, p. 179-185. [ Links ]
15. GOLIJOW, C.D.; et al. Genetic variability and population structure in loci related to milk production traits in native argentine Creole and commercial argentine Holstein cattle. En: Genetics and Molecular Biology. 1999. Vol. 22, no. 3, p. 395-398. [ Links ]
16. CHRENEK, P. Simultaneus analysis of bovine growth hormone and prolactin alleles by multiplex PCR and RFLP. En: Czech Journal of Animal Science. 1998. Vol. 43, no. 2, p. 53-55. [ Links ]
17. RIPOLI, María.; et al. Formación de un plantel base de ganado bovino criollo argentino para producción lechera. Efecto sobre las frecuencias génicas de los loci de k-caseína, S1-caseína y prolactina. En: Archivos de Zootecnia. 1999. Vol. 48, p. 101-106. [ Links ]
18. BRYM, Pawel, KAMINSKI, Stanislaw and WOJCIK, Elzbieta. Nucleotide sequence polymorphism within exon 4 of the bovine prolactin gene and its associations with performance traits. En: Journal Applied Genetics. 2005. Vol. 45, no. 2, p. 179-185. [ Links ]
19. Acceso GenBank: No. AF426315, M57764. [ Links ]
20. MITRA, A.; et al. Polymorphism at growth hormone and prolactin loci in Indian cattle and buffalo. En: Journal of Animal Breeding and Genetics. 1995. Vol. 112, no. 1, p. 71-74. [ Links ]

