











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The fossil record of pliosaurid plesiosaurs ranges from the Lower Jurassic to the Upper Cretaceous. However, that of the Lower Cretaceous is scarce. The main taxonomically identified material from Lower Cretaceous beds includes several specimens found in layers of the Hauterivian of western Russia (Fischer et al., 2015; Fischer et al., 2017), of the Barremian and Aptian of Colombia (Hampe, 1992; 2005; Páramo-Fonseca et al., 2016; Gómez-Pérez and Noé, 2017); and of the Aptian and Albian of Australia (Longman, 1924; Kear, 2003). The Russian specimens comprise a fragmentary skeleton of the holotype of Makhaira rossica Fischer et al., 2015, and a nearly complete, three-dimensionally preserved skeleton, the holotype of Luskhan itilensis Fischer et al., 2017 (Fischer et al., 2015; Fischer et al., 2017). The Colombian material includes a skull and cervical vertebrae recently identified as Acostasaurus pavachoquensis Gómez-Pérez and Noé, 2017 (Gómez-Pérez, 2001; Gómez-Pérez and Noé, 2017) and two nearly complete three-dimensionally preserved skeletons that represent the holotypes of Stenorhynchosaurus munozi Páramo-Fonseca et al., 2016 (Hampe, 2005; Páramo-Fonseca et al., 2016) and '"Kronosaurus" boyacensis Hampe, 1992 (Hampe, 1992) (as will be discussed later, the inclusion of "K." boyacensis in the genus Kronosaurus is considered doubtful, and for this reason it is written in quotes). Finally, the Australian material comprises fragmentary remains assigned to Kronosaurus queenslandicus Longman, 1924 (Longman, 1924; 1930; 1935; White, 1935; Romer and Lewis, 1959; Kear, 2003; McHenry, 2009).
The pliosaurids of the Lower Cretaceous of Colombia are also represented by abundant material from Villa de Leiva not yet identified (Páramo-Fonseca, 2015). Other Cretaceous pliosaurid records includes teeth of PolyptychodonOwen, 1841 from middle to upper Albian beds of England (Madzia, 2016) and from upper Albian to middle Cenomanian beds of Poland (Bardet et al., 2016; Madzia and Machalski, 2017) (Polyptychodon interruptus Owen, 1841, the type species of the genus, is considered nomem dubium (Madzia, 2016)).
The Upper Cretaceous pliosaurid records include some cranial and postcranial remains collected from the Turonian of North America (Williston, 1903; 1907; Schumacher et al., 2013) and Morocco (Angst and Bardet, 2016). This material supports two taxa: Brachauchenius lucasi Williston 1903 and Megacephalosaurus eulerti Schumacher, Carpenter and Everhart, 2013.
Recently, a new specimen (MP111209-1) was found in the Barremian beds of Sáchica, a town near Villa de Leiva. It is the most complete skeleton found in the region. It was excavated in 2013 in the framework of a paleontological exploration carried out by the Universidad Nacional de Colombia and ECOPETROL S.A, the Colombian petroleum company.
The new specimen was found southeast of the town of Sáchica (Figure 1). It was excavated from argillaceous gypsiferous shales in an abandoned quarry that was exploited more than 30 years ago for the mining of gypsum. These rocks are identified as beds of the middle part of the Arcillolitas Abigarradas Member of the Paja Formation (sensu Etayo-Serna, 1968; 1979) (Figure 1) that are equivalent to the upper part of segment C and lower part of segment D of Etayo-Serna (1968; 1979), and the upper part of segment II and segment III of Forero and Sarmiento (1985). An ammonite, Heinzia sp. (Etayo-Serna, personal communication) was collected from beds directly overlying the pliosaur bearing beds. In Colombia this genus indicates the lower part of the upper Barremian (Patarroyo, 2004). A detailed stratigraphic and biostratigraphic study of the rocks in which the pliosaurid remains were found is the subject of an upcoming publication (Ramos and Páramo-Fonseca, in prep.).
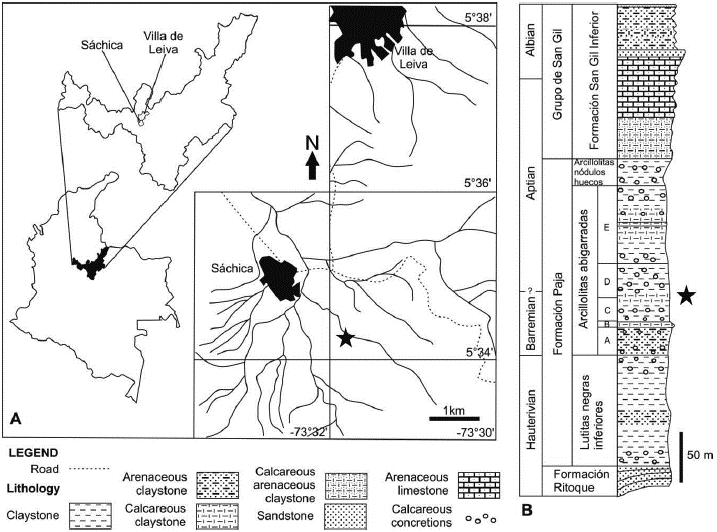
Figure 1 Geographic and stratigraphic origin of Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. A, map of Villa de Leiva - Sáchica region with geographical location of the finding site (marked by a star). B, generalized stratigraphic column of the Paja Formation cropping out around Villa de Leiva and Sáchica, showing the specimen occurrence (marked by a star) (modified from Etayo-Serna, 1979).
The aim of this study is to describe the morphological features of the new specimen (MP111209-1). It is a large pliosaurid, of almost 10 m in length with anatomical features resembling that of "K." boyacensis. However, the presence of distinctive features that differentiate it from Kronosaurus and all other pliosaurids suggest that the new specimen belongs to a new genus and species that we name Sachicasaurus vitae gen. et sp. nov.
Material and Methods
The new specimen (MP111209-1) is an almost complete skeleton that only lacks the distal part of the two right fins. It is three-dimensionally preserved and almost entirely articulated. Only a few skeletal remains are minimally displaced from their anatomical position. A fraction of the right side of the skeleton was removed during gypsum mining but several fragments were recovered on the surface around the excavation site and could be fitted into the skeletal structure (Figure 2). The elements of the left side of the appendicular skeleton were found in anatomical position. However, these appendicular elements, including girdles and fins, have been diagenetically distorted, substantially modifying the anatomical morphology of the bones. This condition results in a morphological difference between the appendicular skeleton of the two sides of the specimen that highlights the problems when describing specimens found in evaporite deposits, and the importance of having complete specimens for a correct determination of post-cranial features. Details of the preservation state will be given in the anatomical description section.
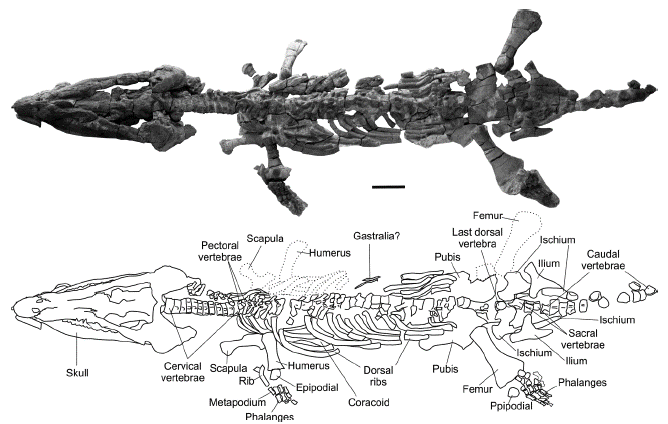
Figure 2 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. Photograph and schematic drawing of the skelet al remains. Doted lines: elements found separated from the skeleton. Scale bar=500 mm.
The fossil was taken out from the excavation site in 14 blocks and each block was prepared mechanically using pneumatic hammers. The skull and the cervical vertebrae were dorsally and ventrally cleaned while the postcranial skeleton was only cleaned dorsally to keep the bone elements in place. Once the blocks were prepared, the specimen was assembled in the same position that it was found.
Because MP111209-1 does not fit completely in the definition of "K." boyacensis, further detailed comparison with K. queenslandicus was done. The published works describing the fragmentary remains of K. queenslandicus (Longman, 1924; 1930; 1935; White, 1935; Romer and Lewis, 1959; Molnar, 1982; 1991; Kear, 2003) were carefully revised to identify the traits used to define the genus Kronosaurus Longman, 1924 (Longman, 1924; Kear, 2003). The unpublished PhD thesis of McHenry (2009) provided important descriptive and photographic information about the material assigned to Kronosaurus.
In order to infer the phylogenetic position of the new specimen, a phylogenetic analysis was performed using the data matrix of Fischer et al. (2017), which complements that of Benson and Druckenmiller (2014) for Plesiosauria. Two new OTUs were incorporated to the matrix: "K." boyacensis and A. pavachoquensis, from the Paja Formation of Colombia, making a total of 99 OTUs in the data set. Most of the character states of "K." boyacensis were scored according to Hampe (1992) and the following states were adjusted based on author's observations (M.E.P.F and C.D.B.C.): the number of premaxillary teeth [131] was scored as [?] given that the the premaxilla-maxilla contact cannot be established in the holotype. The number of dorsal vertebrae [179] was scored as [1] because the holotype of "K." boyacensis has two pectoral and 20 dorsal vertebrae instead of the three pectoral and 19 dorsal reported by Hampe (1992). The scoring of A. pavachoquensis was based on the descriptive works of Gómez-Pérez (2001) and Gómez-Pérez and Noé (2017). Given that there are no posterior small alveoli in premaxilla (Gómez-Pérez and Noé, 2017: Text-fig 3B), the regularity of the posterior premaxillary dentition [132] was scored as homodont [0], and not heterodont as mentioned by Gómez-Pérez and Noé (2017). A list with all character states of the new OTUs is shown in appendix A.
The cladistic analysis was run on TNT (v1.5) (Goloboff and Catalano, 2016), a free program available with the sponsorship of the Willi Hennig Society. An initial exploration for the shortest-length tree islands was performed using the new technology search options (Ratchet + Drift + Tree fusing) with 200 ratchet iterations (1 random seed) and 20.000 trees limit. The resulting trees were then used as the starting point for a Tree bisection reconnection (TBR) branch swapping algorithm. A strict consensus was applied and the iterPCR algorithm was run in order to determine the unstable OTUs in the consensus (Pol and Escapa, 2009). A reduced strict
Anatomical Abbreviations
a, angular; acs, acetabular surface; aiv, anterior interpterygoid vacuity; ar, articular; at, atlas; ax, axis; bo, basioccipital; bs, basisphenoid; c, coronoid; c#, cervical centra number; cav, caudal vertebrae; d, dentary; d#, dorsal vertebrae number; D#, dentary tooth number; db, dorsal blade; dr, dorsal rib; dr#, dorsal rib number; dv, dorsal vertebrae; ec, ectopterygoid; en, external nares; epd, epipodials; ex-op, exoccipital-opisthotic; f, frontal; fe, femur; glr, glenoid ramus; hu, humerus; h, hyoid; in, internal nares; j, jugal; lil, left ilium; lis, left ischium; lj, left jugal; lmx, left maxilla; lp, left pubis; lptz, left postzygapophysis; lsq, left squamosal; mx, maxilla; Mx#, maxillary tooth number; mpd, mesopodials; na, neural arch; nc, neural canal; ne, neural spine; p, parietal; p#, pectoral vertebrae number; pdf, pedicles facets; pdp, posterodorsal process; pf, prefrontal; phl, phalanges; pif, pineal foramen; piv, posterior interpterygoid vacuity; pl, palatine; pmx, premaxilla; Pmx#, Premaxillary tooth number; po, postorbital; pof, postfrontal; popr, paraoccipital process; pr#, pectoral rib number; pra, prearticular; pro, prootic; ps, parasphenoid; pt, pterygoid; q, quadrate; r, rib; rar, retroarticular process; rf, rib facet; ril, right ilium; ris, right ischium; rj, right jugal; rmx, right maxilla; rp, right pubis; rptz, right postzygapophysis; rsq, right squamosal; rtp, rostrum perforations; sa, surangular; sf, subcentral foramina; so, supraoccipital; sp, splenial; sq, squamosal; sv, sacral vertebrae; t, tooth; tap, transverse process; v, vomer; vf, vascular foramina; vp, ventral plate; vr, vertebral ramus.
Nomenclatural Acts
This publication, and the nomenclatural act contained herein, has been registered in ZooBank, the ICZN online registration system for new taxa. The ZooBank Life Science Identifier (LSID) for this publication is urn:lsid:zoobank.org:pub:8EE2C3B0-FBFB-4EA4-9FDA-673AC2AF1CF4. The LSIDs for the new taxon are given under the Systematic paleontology section (below).
SYSTEMATIC PALEONTOLOGY
PLESIOSAURIA de Blainville, 1835
PLIOSAUROIDEA Welles, 1943
PLIOSAURIDAE Seeley, 1874
BRACHAUCHENINAE Benson and Druckenmiller, 2014
Sachicasaurus gen. nov.
urn:lsid:zoobank.org:act:B82F603B-79B0-4616-8040-56DCB649D0EF
Type and only species: Sachicasaurus vitae, sp. nov.
Type Locality: Vereda Arrayan, Sáchica, Boyacá, Colombia; coordinates: 5° 34' 17.33" N, 73° 31' 51.29" W (Figure 1).
Stratigraphic horizon: Arcillolitas Abigarradas Member, Paja Formation. Barremian to Aptian, Lower Cretaceous (Figure 1).
Derivation of name: From Sáchica, locality where the fossil was found and greek saurus, lizard.
Diagnosis: As for type and only known species. Sachicasaurus vitae sp. nov. urn:lsid:zoobank.org:act:B62884A0-ADE1-4B13-A37D-D0B1C32C90E1
Holotype: MP111209-1, a nearly complete skeleton, lacking the distal part of the right fins (Figure 2). At the present time the specimen is part of the material collected for a research project of the Universidad Nacional de Colombia, but it is to be administrated by the Colombian geological survey (SGC: Servicio Geológico Colombiano).
Derivation of name: From the Latin vitae, life. It refers to the vitality that this fossil has given to the Sáchica village since its discovery.
Diagnosis: Sachicasaurus vitae (MP111209-1) is a brachauchenine pliosaurid characterized by two autapomorphies: a very short mandibular symphysis ending at the mid length of the fourth mandibular alveoli and highly reduced number of mandibular teeth (17-18). It is distinguished from all other pliosaurid, except L. itilensis in having an ilium with a long posterior projection. It differs from all other brachauhenines in having a pineal foramen in a very anterior position (similar to L. itilensis) and cervical centra without dorsal but with ventral foramina. Sachicasaurus vitae is also distinguished from other bracauchenines by the following unique combination of characters: skull exceeding two meters in length (as in K. queenslandicus and "K." boyacensis), lateral expansion of the maxilla posterior to the premaxilla-maxilla suture (unlike B. lucasi), premaxilla with four teeth (unlike in L. itilensis), caniniform teeth present (as K. queenslandicus and "K." boyacensis), dental sequence showing only two non-contiguous largest teeth (D4, Mx1) (probably also in "K." boyacensis, C.D.B.C. pers. obs.), posteriorly bifurcated vomer (asM. eulerti), palatines separated in the midline by the pterygoids (unlike B. lucasi and probably K. queenslandicus), slender hyoid bones (unlike K. queenslandicus), 12 cervical vertebrae (as K. queenslandicus and "K." boyacensis), cervical centra showing ventral foramina (also in L. itilensis), single rib facet in all cervical centra (unlike M. eulerti), rib facets of posterior cervical non divided horizontally (unlike "K." boyacensis), at least 37 pre-sacral vertebrae (unlike "K." boyacensis), and zygapophyses present in posterior dorsal vertebrae (unlike K. queenslandicus).
Ontogenetic Stage
Based on the ontogenetical characteristics proposed by Brown (1981) for plesiosaurs, we interpret MP111209-1 as a sub-adult individual. As an adult, the specimen presents a well-ossified sutured skull, fully formed teeth exhibiting noticeable ridges on their enameled crowns, and capitulum and tuberosity or trochanter differentiated on propodials. However, it preserves the cervical vertebrae without fused ribs, almost flat articular facets of the vertebral centra, and the atlas and axis fused but with indication of the suture between them, all of which suggest traces of a juvenile stage.
DESCRIPTION
The specimen MP111209-1 is an almost complete articulated skeleton that lost the distal part of the two right fins and a right portion of the rib cage (Figure 2). It is possible that some posterior caudal vertebrae are missing because there is no evident reduction in size in the preserved ones. The total length of the specimen, measured from the anterior tip of the snout to the posterior end of the last preserved caudal vertebra, is 990 cm.
The skull shows dorsoventral compression. Nevertheless, the snout including the mandibular symphysis is preserved in a tridimensional state. There are two irregular openings located near the midline in the anterior dorsal surface of the snout that seems to correspond to taphonomic distortion.
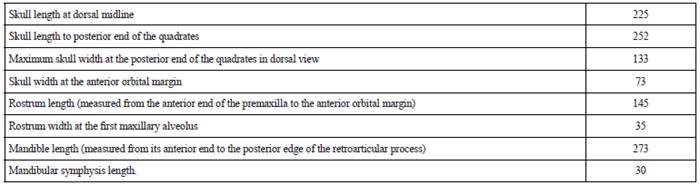
Cranium (Figures 3 and 4; Table 1)
The skull-roof region is collapsed, and the rostrum is preserved between the mandibular rami, which are slightly tilted dorsomedially. The external nares are almost closed due to the collapsing of the dorsal bones; the left one is more visible than the right. The circumorbital region of the skull is crushed so its bones cannot be identified with certainty. In dorsal view the cranium is triangular in outline, with a long and narrow snout and a wide and robust posterior region (for measures see table 1). The preorbital skull length is greater than half of the total skull length, as in brachauchenine pliosaurids (Williston, 1907; McHenry, 2009; Schumacher et al., 2013; Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016; Fischer et al., 2017), PeloneustesphilarchusSeeley, 1869 (Andrews, 1913; Ketchum and Benson, 2011a) and HauffiosaurusO'Keefe, 2001 (O'Keefe, 2001; Benson et al., 2011). The alveolar margin of the upper jaw in lateral view is not regular, as in some non-brachauchenine thalassophoneans (Andrews, 1913; Sassoon et. al., 2012; Benson et al., 2013).
Table 1 Measurements (in cm) of the skull and mandible of Sachicasaurus vitae gen. et sp. nov, holotype MP111209-1.
The premaxilla is a long bone with a wide anterior region and a narrow posterodorsal process. Each premaxilla bears four alveoli, as in Brachauchenius (Albright et al., 2007), M. eulerti (Schumacher et al., 2013) and A. pavachoquensis (Gómez-Pérez and Noé, 2017). Posterior to the four premaxillary teeth there is a diastema as long as the mesiodistal length of an alveolus where the fourth large dentary tooth is accommodated. The premaxilla-maxilla suture ascends dorsally interdigitated through the diastema, and then it abruptly turns posterodorsally (Figures 4A and B). The posterodorsal process contacts posteriorly the parietal and frontal in an interdigitating suture (Figure 3A). The anterior surface of the premaxilla has numerous dispersed vascular foramina.
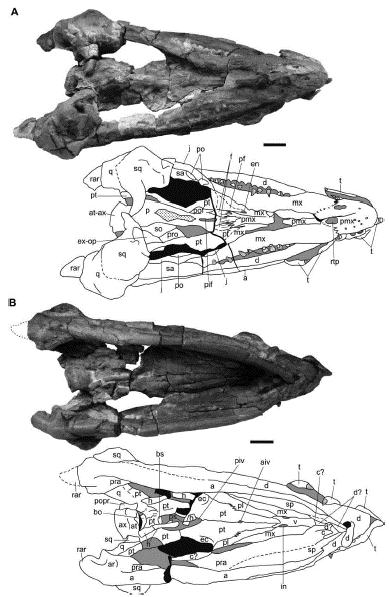
Figure 3 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. Photographs and interpretive schemes of the skull in A, dorsal and B, ventral views. Dotted lines: interpretive limits; dotted surfaces: broken bones; in gray: sediment; in black: empty spaces. Scale bars=200 mm
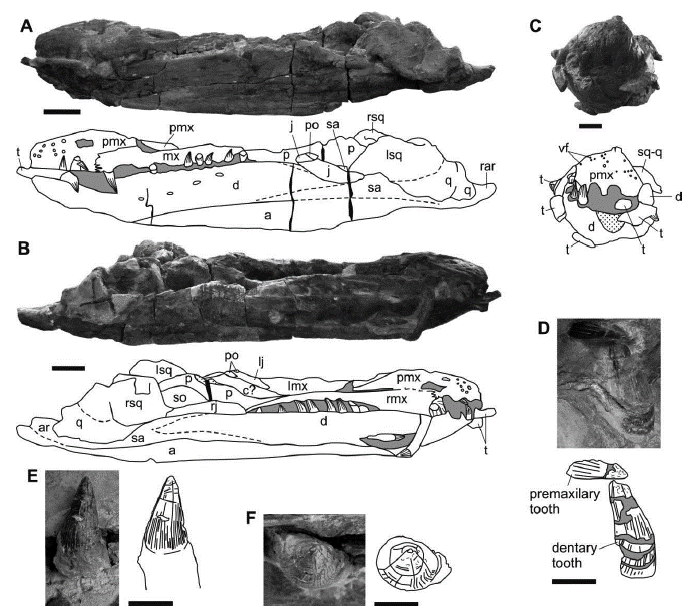
Figure 4 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. Photographs and interpretive schemes of: A-C, skull in A, right lateral, B, left lateral and C, anterior views. D, third premaxillary (replacement tooth) and second dentary teeth in labial view; E-F, ninth dentary tooth in E, labial and F, apical views. Dotted lines: interpretive limits; dotted surfaces: broken bones; in gray: sediment; in black: empty spaces. Scale bars: A-B=200mm in; C=100mm; D-F=40mm.
The maxillae are dorsally crushed. Posterior to the diastema, each maxilla expands laterally and ventrally to accommodate a large caniniform tooth (M1) (Figure 3A), differing from most of the basal pliosaurids and from S. munozi and B. lucasi, in which no expansion is present (Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016). The first maxillary teeth are the largest and occlude externally to the dentaries. Posteriorly, the maxillary teeth are reduced in size and occlude internally to the mandibles. Different than in K. queenslandicus (McHenry, 2009), there are no two large adjacent teeth in the maxilla. In lateral view, the maxillae extend backwards to the level of the middle of the orbit. Medial to the external nares the maxillae extend behind the posterior margin of the narial openings, forming the anterior and dorsal margins of the nares (Figure 3). This configuration is also found in other brachauchenine pliosaurids (Williston, 1903; 1907; Schumacher et al., 2013; Fischer et al., 2017). In palatal view, the maxilla is widely exposed (Figure 3B). It forms the lateral border of the internal nares and contacts the vomer anteriorly and the palatine posteriorly.
The morphology of the frontals, prefrontals, and postfrontals cannot be determined precisely due to fracturing and loss of bones in this region. However, it is possible to note that the posterior and ventral margins of the nares are limited by prefrontals (Figure 3A).
The parietals are fused. The pineal foramen is elongated and forms a long and narrow concavity located in the anterior region of the parietal at the level of the anterior portion of the orbits (Figure 3A). This character is only shared with L. itilensis among the brachauchenines (Williston, 1907; Schumacher et al., 2013; Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016; Fischer et al., 2017). In front of the pineal foramen the parietal is ornamented with numerous ridges oriented anteroposteriorly, which are located just posterior to the premaxilla-parietal suture and obscure the contact with the frontals (Figure 3A). Behind the pineal foramen there is a broken sagittal crest. The crest fragment is high and narrow and is preserved near its anatomical position.
The jugals are preserved on the external surface of the mandibular rami. Each jugal is a long and slender bone that contact the squamosals posteriorly (Figures 3A, 4A and B). The jugal seems to have formed at least a small portion of the temporal bar, as in other brachauchenines, except L. itilensis (Fischer et al., 2017).
On each side of the skull, medial to the jugal, there is a fragment of the postorbital. Only some fragments of the ventral region of the postorbitals are recognizable. The squamosal and quadrate are massivelly-built bones (Figures 3, 4A and B). These bones are taphonomically moved and crushed down against the mandible. The sutures between them are no identifiable. The dorsomedial ramus of the squamosal is anteroposteriorly compressed. It extends posterolaterally to contact the quadrate.
The bones of the braincase are only partially exposed on the right and ventral sides of the skull, and it is not possible to say much about their morphology (Figures 3B and 4B). The dorsoventral compression of the cranium caused the skull roof bones to be in contact with those of the palate. Only a small fraction of the external surfaces of the supraoccipital, prootic and exoccipital-opisthotic can be seen in dorsolateral view. In ventral view the basioccipital is barely exposed; it is ventrally covered by the pterygoids. The basioccipital condyle articulates with the atlas-axis complex, so its morphology remains unknown. A small part of the anterior end of the basisphenoid and two short fragments of a narrow parasphenoid are visible through the posterior interpterygoid vacuity; their morphology remains unclear (Figure 3B).
The ventral part of the palate is well exposed; its anterior region is well preserved, but its anterior end is obscured by the mandibular symphysis (Figure 3B). The internal nares are located halfway between the anterior end of the snout and the anterior margin of the posterior interpterygoid vacuity. They are small and oval in outline and are enclosed by the vomers and maxillae. The vomers are fused in a single element that forms the medial border of the internal nares. The posterior end of the vomer is divided into two rami by a medial anterior prolongation of the pterygoids, in the same way as inM. eulerti (Schumacher et al., 2013). Laterally, these two vomer rami contact the palatines. The palatines are separated medially by the pterygoids and thus, do not meet in the midline (Figure 3B), differing from B. lucasi (Williston, 1903; Schumacher et al., 2013), Liopleurodon feroxSauvage, 1873 (Noe, 2001) and probably K. queenslandicus (McHenry, 2009). The exposed ventral surface of the palatine is lateromedially narrow. It contacts the pterygoid medially in a long straight suture that becomes zigzagging in the middle. Laterally, the palatine limits the maxilla in a straight suture. Posteriorly, the palatines contact the ectopterygoids, but the suture between them is not clear.
The pterygoids are medial broad plates located between the palatines. They enclose an anterior interpterygoid vacuity, as in M. eulerti (Schumacher et al., 2013) and S. munozi (Páramo-Fonseca et al., 2016), that is small and oval in outline (Figure 3B). The pterygoid expands laterally to contact the ectopterygoid. It borders laterally and posteriorly the posterior interpterygoid vacuity, forming a lateral flange. The flanges of both pterygoids are crushed, but it can be seen that they unite in the midline behind the posterior interpterygoid vacuity. The distance from the posterior border of the interpterygoid vacuity to the posterior end of the pterygoids in the midline is short. The pterygoids join anterior to the posterior interpterygoid vacuities, covering the parasphenoid, as in most of the brachauchenine pliosaurids (Williston, 1903; Schumacher et al., 2013; Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016). The pterygoids cover ventrally the basicranium. Here, the evidence of a short suture in the posterior border, lateral to the midline suggests the pterygoids slightly separate posteriorly, exposing a small portion of the basioccipital (Figure 3B).
Both the ectopterygoids are incomplete; their lateral end is broken. The ectopterygoid forms, together with the pterygoid, a laterally curved flange (Figure 3B) and contacts anteriorly the palatine. The presence of a suborbital fenestra cannot be established because the ectopterygoids are fractured in this region. The hyoids are slightly broken and displaced from their anatomical position. The right one is located ventrally over the posterior interpterygoid vacuity and the left one is moved to the right edge of the cranium (Figure 3B). Differing from K. queenslandicus (Kear, 2003), the hyoids are long and slender.
Mandible (Figures 3 and 4)
The mandibular rami are robust and high and are partially exposed lateral to the snout in dorsal view (Figures 3, 4A and B). In ventral view the mandibular symphysis tapers anteriorly, as in Hauffiosaurus (O'Keefe, 2001; Benson et al., 2011) and brachauchenine pliosaurids (Williston, 1903; Schumacher et al., 2013; Páramo-Fonseca et al., 2016; Fischer et al., 2017). The symphysis is particularly short, being 1/9 of the total mandible length (table 1) and ends in the middle of the fourth dentary tooth. The dentary is exposed laterally in the anterior region of the mandibular ramus. As the jaws are occluded, the anterior end of the dentary is partially covered by the premaxilla and the maxilla. The dentary does not have a ventral elaboration along the mandibular symphysis, differing from non-brachauchenine thalassophoneans and from Hauffiosaurus longirostris Benson, Ketchum, Noé and Gómez-Pérez, 2011 (Benson et al., 2011). Posteriorly, the dentary contacts the surangular apparently in a V-shaped suture with its vertex pointing backwards. The contact between the surangular and the articular is covered taphonomically by the squamosals and quadrates. The angular forms most of the ventral surface of the mandibular rami. It extends anteriorly to the level of the first maxillary tooth and posteriorly to the retroarticular processes (Figures 3B and 4B). The retroarticular processes are preserved, but the bones forming them are not identifiable. They are posteromedially inflected, as in other brachauchenine pliosaurids (Williston, 1903; 1907; Schumacher et al., 2013; Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016; Fischer et al., 2017). Medially, the splenial contacts the dentary and participates in the mandibular symphysis (Figure 3B). The splenial is a long bone that extends posteriorly contacting the angular. Dorsal to the splenial, the coronoid and the prearticular form the medial wall of the mandibular ramus, but their limits are difficult to identify. Only the anterior end of the coronoid is recognized; it forms the posterior edge of a small mandibular opening (Figure 3B).
Teeth (Figure 4)
The description of the tooth morphology follows the terminology proposed by Zverkov et al. (2018). The dentition is anisodont (heterodont), as in K. queenslandicus (Kear, 2003; McHenry, 2009), "K." boyacensis (Hampe, 1992) and in non-brachauchenine pliosaurid (Andrews, 1913; O'Keefe, 2001; Benson et al., 2011; Sassoon et. al., 2012; Benson et al., 2013; Benson and Druckenmiller, 2014; Gómez-Pérez and Noé, 2017). The premaxillary and anterior dentary alveoli are large, but the fourth dentary and the first maxillary are the largest alveoli (Figures 4A, B and C). This condition differs from what is found in other heterodont pliosaurids, in which the largest maxillary alveoli are generally two in each mandibular ramus and they are in a more posterior position (e.g. A. pavachoquensis, L. ferox, Simolestes vorax Andrews, 1909, Pliosaurus kevani Benson et al., 2013, K. queenslandicus) (Noé, 2001; McHenry, 2009; Benson et al., 2013; Gómez-Pérez and Noé, 2017). All upper and lower teeth are intercalated; the teeth succession in the anterior snout is: Pmx1, D1, Pmx2, D2, Pmx3, D3, Pmx4, D4, Mx1, D5, Mx2. That differs from K. queenslandicus were there are two contiguous dentary teeth (the largest teeth) followed by two adjacent maxillary teeth (the largest ones) (D4, D5, Mx1, Mx2) (McHenry, 2009). There are four teeth of similar size in each premaxilla. Differing from some thalassophoneans, there are no small teeth in the front of the snout (Figure 4C) (Andrews, 1913; Noé, 2001; Benson et al., 2013; Schumacher et al., 2013; Gómez-Pérez and Noé, 2017; Fischer et al., 2017). The mesiodistal length of the premaxillary alveoli from first to fourth are: 50 mm, 60 mm, 80 mm and 85 mm. Different than M. eulerti and L. itilensis (Schumacher et al., 2013; Fischer et al., 2017), in MP111209-1 the alveoli are narrowly spacing, as it is in almost all pliosaurids. the distance measured from the midpoint of contiguous premaxillary alveoli are 105 mm from the first to the second, 100 mm from the second to the third, 110 mm from the third to the fourth and 150 mm from the fourth to the first maxillary alveoli. In the left maxilla the largest tooth is in anatomical position and exhibits its complete crown. This crown is 90 mm in height and 70 mm in basal diameter. The large maxillary tooth of the right side is displaced out of its alveolus (Figure 4B). It has a long root, with a smooth surface. Its enameled crown is distally fractured. The height of this tooth including its preserved portion of crown and its root is 300 mm; the diameter of the root, measured at the limit with the crown, is 90 mm.
Almost all the alveoli of the dentaries can be observed either with or without teeth. In both sides of the mandible, the distal teeth are the smaller; the distalmost ones are not clearly differentiable. Mesial to the first maxillary tooth there are four dentary teeth positions and distal to it, 12 to 13 dentary teeth positions can be counted. Adjacent to the first maxillary teeth the dentaries are covered by the maxillae and here the dentary alveoli cannot be seen laterally or dorsally. However, in a diagonal fracture that passes through this area in the left side of the cranium, teeth are observed in section and only one dentary tooth is covered laterally by the maxilla. Therefore, the estimated number of dentary teeth is 17 or 18.
Many of the tooth crowns show transverse fractures (Figures 4D, E and F) and, in some, the apex is slightly displaced and bended. All of the tooth crowns are conical and have a circular cross section (Figure 4D, E and F) as in early pliosaurids and in brachauchenines except L. itilensis (Benson and Druckenmiller, 2014; Cau and Fanti, 2016; Gómez-Pérez and Noé, 2017; Fischer et al., 2017). The surface of the crowns has several prominent ridges but lacks carinae. It is ornamented with ridges even in the newly emplaced teeth (Figure 4D). Some of the ridges reach the tip of the crown, but the others only reach halfway through the crown height. Close to the apex, there are several ridglets on the crown surface. (Figure 4D).
Axial Skeleton (Figures 2 and 5; Table 2)
The vertebral column is well preserved, with some disarticulated vertebrae (Figure 2). The atlas-axis complex is partially fused and a vestige of the suture between them can be seen ventrally. The atlas has a bulged anterior surface. Only the axis has a fused rib. There are 12 cervical vertebrae including the atlas and axis (Figure 5A), as there are in "K." boyacensis (Hampe, 1992), B. lucasi (Williston, 1907), and K. queenslandicus (Kear, 2003) (although for McHenry, 2009 the number of cervical vertebrae in K. queenslandicus is 13), and fewer than in L. itilensis (14) and S. munozi (17) (Páramo-Fonseca, et al., 2016; Fischer et al. , 2017). The cervical centra are articulated. They are wider than high and much higher than long (Figures 5B, C and D; table 2) as in all other brachauchenines (Williston, 1903; Romer and Lewis, 1959; Fischer et al., 2017). Their articular surface is subcircular and nearly flat. The ventral surface of the cervical centra does not have ridges but has paired foramina located near its ventrolateral borders; these foramina are longitudinally oval (Figure 5B). Ventral foramina in cervical centra are also present in other Cretaceous pliosaurids as L. itilensis and A. pavachoquensis (Fischer et al., 2017; Gómez-Pérez, 2001) but are absent in K. queenslandicus, B. lucasi and S. munozi (Williston, 1903; McHenry, 2009; Páramo-Fonseca et al., 2016). The dorsal surface of the cervical centra lacks vascular foramina (Figure 5C), as in A. pavachoquensis (Gómez-Pérez, 2001) but unlike in B. lucasi, S. munozi and L. itilensis (Williston, 1903; Páramo-Fonseca et al., 2016; Fischer et al., 2017). The cervical rib facets are single-headed and are located laterally as in brachauchenine pliosaurids (Williston, 1903; Romer and Lewis, 1959; Benson and Druckenmiller, 2014; Páramo-Fonseca et al., 2016; Fischer et al., 2017) excluding M. eulerti which possesses double-headed ribs (Schumacher et al., 2013). They have a rectangular outline and occupy almost the entire length of the cervical centrum (Figure 5D), corresponding to the same arrangement of "K." boyacensis (Hampe 1992). The rib facets of the posterior cervical centra are in a more dorsal position (Figure 5A) and they are no divided into anterior and posterior portion as it is seen in "K." boyacensis (Hampe, 1992). The cervical ribs are only preserved in the axis. The cervical neural arches are articulated with each other and disarticulated from the centra. They are preserved at the right side of the cervical centra (Figure 5A). The neural canal is not exposed in any of the vertebrae, so its shape and size cannot be established. The neural spine is straight and inclined posterodorsally as in K. queenslandicus and L. itilensis (Romer and Lewis, 1959; Fischer et al., 2017). The prezygapophyses and postzygapophyses have their articular surface ventromedially inclined but its medial relationship cannot be seen.
Table 2 Measurements (in cm) of the vertebral centra dimensions of Sachicasaurus vitae gen. et sp. nov, holotype MP111209-1.
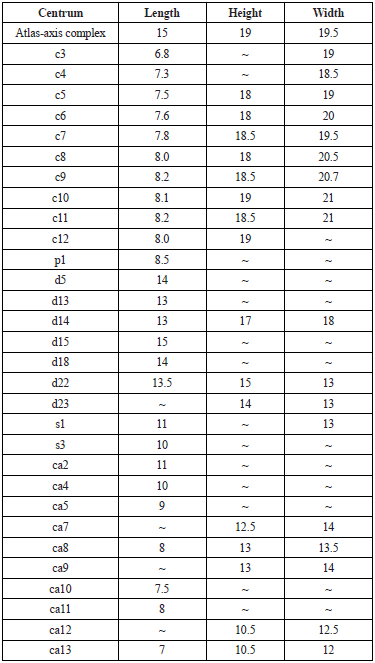
Cervical (c), pectoral (p), dorsal (d), sacral (s), and caudal (ca) centra.
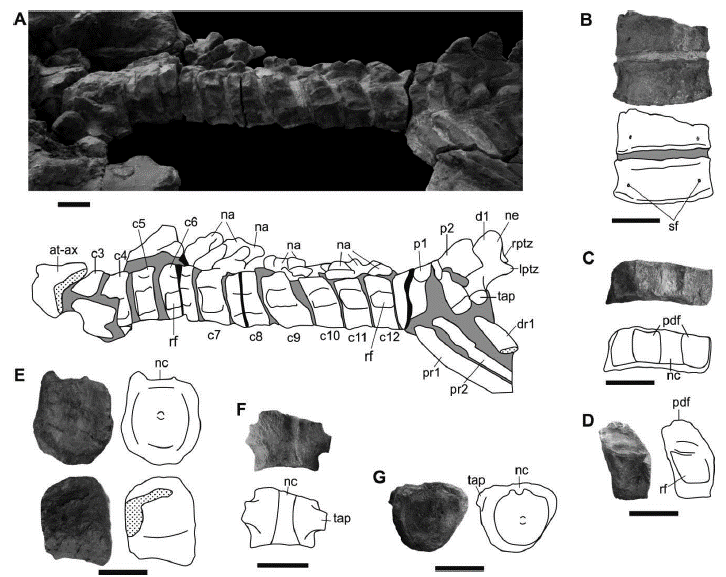
Figure 5 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. Photographs and interpretive schemes of axial skeleton elements. A, cervical and pectoral vertebrae series; B, seventh cervical centrum and a fragment of the sixth one in ventral view; C, seventh cervical centrum in dorsal view; D, 11th cervical centrum in left lateral view; E, 22th dorsal vertebrae in posterior and left lateral views; F, fourth caudal centrum in dorsal view; G, ninth caudal centrum in anterior view. Dotted surfaces: broken bones; in gray: sediment; in black: empty spaces. Scale bars=100 mm.
There are two pectoral vertebrae. Their neural arches and those of the anteriormost dorsal vertebrae are not fused and are slightly displaced from their anatomical position (Figure 5A). The transverse processes of the pectoral vertebrae are robust and have subcircular articular surfaces. There are at least 23 dorsal vertebrae. The dorsal centra are longer than the cervical ones (table 2) and they are longer than they are wide. In anterior dorsals the transverse processes project laterally at the same level of the prezygapophyses but ventral to the postzygapophyses. The most posterior dorsal centra are higher than wide and their articular facets are almost flat (Figure 5E). After the 7th dorsal vertebra the neural arches are poorly preserved and the vertebral centra are rotated 90 degrees to the right side. The last dorsal neural arches (17-19 dorsal vertebrae) clearly show zygapophyses.
There are three sacral vertebrae that are articulated but slightly displaced backwards, being separated from the dorsal series. Their neural arches are crushed over the centra, so the sacral vertebrae are not clearly differentiated. There are 13 caudal vertebrae preserved, the anterior four are articulated with the sacral. Although no other posterior caudal vertebrae were found during the excavation, it seems that the caudal series is incomplete, because the preserved centra do not show a decrease in size (table 2). The posterior caudal vertebrae are disarticulated but form a lineal series, which indicates that they are close to their anatomical position (Figure 2). The caudal centra have a subtriangular shape in anterior and posterior view; they are as high as wide and have nearly flat articular surfaces (Figure 5G; table 2). In these vertebrae, the articular ribs facets are located on the centra (Figure 5F). There are no vestiges of neural or haemal arches in the preserved caudal vertebrae.
The ribs are slightly displaced from their in vivo position, so that their proximal extremities are not in direct articulation with the vertebrae. Two rib fragments, plastically deformed, lie on the distal end of the right humerus (Figure 2). The rib cage is compressed towards the vertebral column in both sides of the specimen (Figure 2). The pectoral ribs, as well as the first and last dorsal ribs, are shorter than the other dorsal ribs. The dorsal ribs are curved in their proximal region and become straight towards their distal region. The ribs are robust and oval in cross section. The largest proximal diameter is equivalent to 0.06 times the length of each rib. There are a few poor preserved slender bone fragments lying near the right ribs, which could represent gastralia remains. (Figure 2).
Appendicular Skeleton (Figures 2, 6 and 7; Table 3)
The girdles are well preserved and are covered medially by the axial skeleton (Figures 2 and 6A). Both scapulae are present. The left scapula is found in its anatomical position but partially covered by the axial skeleton while the right one is incomplete and was found off site, separated from the skeleton, a few meters from the excavation site. A complete ventral plate is not observable in any of the scapulae. In the left scapula the dorsal blade is long as in S. munozi and P. philarchus (Andrews, 1913; Páramo-Fonseca et al., 2016) but unlike in L. itilensis (Fischer et al., 2017) and has its distal end mediolaterally expanded as in S. munozi and L. itilensis (Páramo-Fonseca et al., 2016; Fischer et al., 2017) but unlike in P. philarchus (Andrews, 1913). The glenoid ramus is short and thickens distally to articulate with the humerus (Figures 6D and E). The right scapula is broken and preserves the dorsal blade, which is more laterally oriented than that ofthe left scapula (Figure 6D; table 3). The dissimilar orientation of the dorsal blade of both scapulae seems to be due to taphonomical deformation, so the original orientation of this blade cannot be established with certainty. The coracoids are almost entirely covered by the axial skeleton, so their morphology and their medial relationship cannot be determined. It can only be said that it is a greatly elongated plate with a weakly developed posterolateral cornu as in Hauffiosaurus zanoniO'Keefe, 2001 and Anguanax zignoiCau and Fanti, 2016 (Vincent, 2011; Cau and Fanti, 2016) but unlike in L. itilensis where the cornu is present and extends lateral to the coracoid glenoid ramus (Fischer et al., 2017).
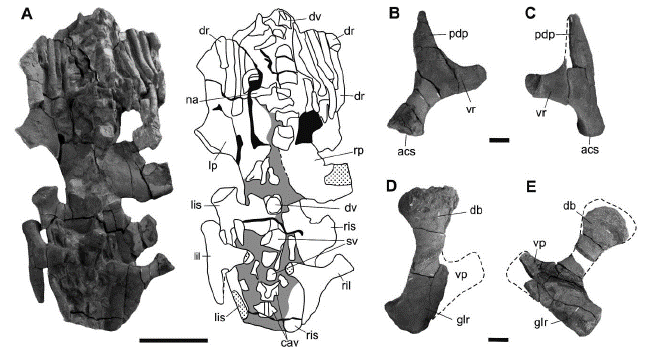
Figure 6 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. A-C, pelvic girdle. A, Photograph and interpretive scheme of pelvic girdle as preserved, in dorsal view; B, right and C, left ilium in anterodorsal view. D-E, scapulae; D, left an E, right scapula in dorsomedial view. Dotted lines: interpretive limits; dotted surfaces: broken bones; in gray: sediment; in black: empty spaces. Scale bars: A=500 mm; B-E=100 mm.
Table 3 Measurements (in cm) of scapular rami and propodials of Sachicasaurus vitae gen. et sp. nov, holotype MP111209-1.

The pelvic girdle is well preserved, but its medial portion is covered by the axial skeleton (Figure 6A). The right bones are slightly displaced backwards with respect to the left ones. Therefore, the precise joint shape between the bones of both sides cannot be established. The ilia lie over the ischia, slightly displaced and rotated backwards (Figures 2 and 6A). Each ilium is a triradiate bone with three rami disposed in the same plane. The dorsal expansion is large, as in P. philarchus (Andrews, 1913) and presents a very elongated posterodorsal process (Figures 6B and C), as it was illustrated by Fischer et al. (2017) for L. itilensis, but it differs from the shorter process described or illustrated for P. philarchus, H. zanoni and Marmornectes candrewi Ketchum and Benson, 2011b (Andrews, 1913; Ketchum and Benson, 2011b; Vincent, 2011). The acetabular ramus is thick and wide. The medial ramus is short and sub-tabular in form. The posterodorsal process is longer than the medial ramus and tapers distally. The pubes are broad, anteroposteriorly elongated plates with a mediolateral dorsal concavity (Figure 6A). They contact each other in the midline, but the join shape is obscured by the axial skeleton. The lateral border of the pubis, anterior to the acetabulum, is concave. Its anterior border is convex, and its posterior border is slightly concave. Both ischia are well preserved and meet medially at least in their anterior portion. The ischium is a very elongated plate with a short and robust acetabular ramus (Figure 6A). The length to width ratio of the ischium (2.02) is similar in K. queenslandicus (1.98) but it is greater than in some basal pliosaurids (Benson and Druckenmiller, 2014). The anterior border of the ischium is concave, and its posterior end is convex and thickened. The right ischium is almost flat, while the left one, as well as the left pubis, are taphonomically distorted with their lateral portions very curved dorsally (Figure 6A).
Both left fins are articulated with the girdles while the right ones were removed during the mining, and only the propodials were found on the surroundings of the excavation site (Figure 2). The left limbs are plastically deformed, especially the hind limb (Figures 7C and D). The profile of these limbs looks wavy in anterior and posterior view. The propodials are thick and wide, dorsoventrally compressed bones. They are slender and pendulous; their distal extremity is fan-shaped. The propodials shaft is relatively robust compared with that of the "K." boyacensis. The femur is longer than the humerus, and in
both, the posterior border is more concave than the anterior one (Figures 7A and B; table 3). The ratio of humerus to femur length (0.87) is greater than in "K." boyacensis (0.82, calculated from the measurements given by Hampe, 1992), and most thalassophoneans (Benson and Druckenmiller, 2014; Fischer et al., 2017) exceptH. zanoni (0.92) and Hauffiosaurus tomistomimus (0.97) Benson, Ketchum, Noé and Gómez-Pérez, 2011 (calculated from the figures of Benson et al., 2011 and Vincent, 2011).
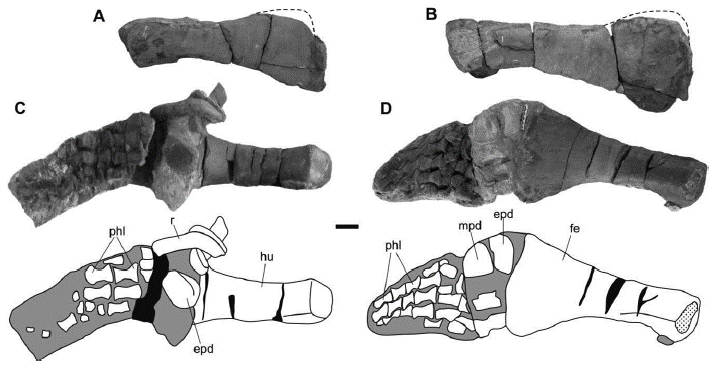
Figure 7 Sachicasaurus vitae gen. et sp. nov., holotype MP111209-1. A-B right propodials; A, humerus and B, femur in dorsal view. C-D, photographs and interpretive schemes of left limbs; C, forelimb and D, hindlimb in dorsal view. Dotted lines: interpretive limits; dotted surfaces: broken bones; in gray: sediment; in black: empty spaces. Scale bar=100 mm.
The humerus length versus width ratio is between 2.3 and 2.7, as in non-thalassophoneans pliosaurids and P. kevani (Benson and Druckenmiller, 2014; Benson et al., 2013). The proximal end of the humerus have a subrectangular outline, that is higher than it is long. The humerus becomes longer than high at the middle of its proximodistal length, and in its distal end it has an elongated oval outline, being much longer than high. The length to distal width ratio of the femur is 2,1, which falls between the range of most pliosaurids. The proximal end of the femur is higher than long; it has two marked articular facets, one dorsal and another ventral. The dorsal facet has a straight dorsal border, ventrolaterally inclined lateral sides and a curved ventral border. The ventral facet is squared in outline. At its proximodistal mid-length, the femur becomes longitudinally oval in cross-section, and distally, it turns significantly flattened dorsoventrally.
The epipodials and mesopodials are poorly preserved (Figures 7C and D), so their morphology cannot be established. The metapodials are relatively well exposed in the forelimb (Figure 7C). Here, four elements that have the shape of elongated phalanges are identified. Five digits can be counted in the hind limb; the best preserved contains five phalanges (Figure 7D). All the preserved phalanges are hourglass shaped.
DISCUSSION
Morphological Comparisons
The new specimen MP111209-1 is considered a member of Pliosauridae because it presents most of the morphological traits of the family as defined by Brown (1981), Druckenmiller and Russell (2008) and Ketchum and Benson (2010). Even though Benson et al. (2013) and Benson and Druckenmiller (2014) recently carried out phylogenetical analyses of Plesiosauria, they did not introduced changes to the previous diagnosis. MP111209-1 shares with other pliosaurids a large skull, premaxilla no participating in the margin of the external nares, coronoid present with a long lingual process, broad and strong teeth, ornamented with longitudinal ridges, reduced number of cervical vertebrae with short cervical centra and ventral surface nearly flat, anteroposteriorly relatively long ischia, and femur larger than the humerus. MP111209-1 differs from the diagnosis of Pliosauridae in having 17-18 dentary teeth, contrasting with the 25 to 40 found in other pliosaurids (Brown, 1981).
The Barremian pliosaurid genera known in Colombia, Stenorhynchosaurus and Acostasaurus, are smaller in size and clearly differ from the new specimen MP111209-1: Stenorhynchosaurus has a more elongated snout, isodont dentition, and longer mandibular symphysis (Páramo-Fonseca et al., 2016); and Acostasaurus has a shorter preorbital rostrum, large nasal present in contact with the parietal, anterior interpterygoid vacuity absent, longer mandibular symphysis, different arrangement of the heterodont dentition (Gómez-Pérez and Noé, 2017), cervical rib facets in ventrolateral position and double-headed cervical ribs (Gómez-Pérez, 2001).
MP111209-1 presents most of the diagnostic characters established by Benson and Druckenmiller (2014) for Brachaucheninae. However, in MP111209-1 the cervical centra possess ventral foramina. This feature is generally present in the non-brachauchenines pliosaurids according to the character matrix of Benson and Druckenmiller (2014). Nevertheless, Fischer et al. (2017) described the presence of ventral foramina in the cervical centra of the early brachauchenine L. itilensis. Furthermore, in MP111209-1 the rostrum is laterally expanded posterior to the level of the premaxilla-maxilla suture, accommodating the first large maxillary tooth, which differs again from the condition found in the brachauchenine pliosaurids (Williston, 1903; Schumacher et al., 2013; Benson and Druckenmiller, 2014).
MP111209-1 also differs from most brachauchenines (and most pliosaurids) in having the pineal foramen in a very anterior position (similar in L. itilensis), an extremely short mandibular symphysis, and a particular morphology of the ilium, with a remarkable posterior dorsal projection, greater than that illustrated by Andrews (1913) for P. philarchus. Unfortunately, the morphology of the ilium is unknown in most brachauchenines, and in L. itilensis the ilium, apparently similar in morphology, is incomplete (Fischer et al., 2017).
MP111209-1 shares traits with the upper Hauterivian brachauchenine genus Luskhan that are not found in other Pliosauridae or in Brachaucheninae, as mentioned above, but also have considerable differences with it, mostly in the number and morphology of the premaxillary teeth. In Luskhan there are seven premaxillary teeth, the mesialmost teeth (1st premaxillary alveoli) being procumbent and separated from the other alveoli (Fischer et al., 2017) while in MP111209-1 there are four premaxillary teeth similar in size.
The other Cretaceous genera Makhaira, Brachauchenius and Megacephalosaurus are also smaller than MP111209-1 and differ from it in having an isodont dentition and a longer symphysis (Williston, 1903; 1907; Schumacher et al., 2013, Fischer et al., 2015). Moreover, Brachauchenius differs from the new specimen in having the palatines united in the midline (Williston, 1903) and Megacephalosaurus differs in having double-headed cervical ribs (Schumacher et al., 2013).
Comparison with Kronosaurus
The genus Kronosaurus was first proposed by Longman (1924) to name, as K. queenslandicus, a fragment of a large mandible from the Albian of Australia. He found this mandibular fragment different from the other plesiosaurs because of its large size and the distinct "contours of the teeth and mandible" (Longman, 1924). Then, Longman (1930) studied some Albian fragments of propodials and White (1935) described cranial fragments of two Aptian specimens. They both assigned these fragments to K. queenslandicus.Molnar (1991) suggested that the specimens studied by White represent a different species from K. queenslandicus based on the age difference of the fragments. K. queenslandicus is now known from numerous fragments from Australia, many of them described but still unpublished (McHenry, 2009).
A second Kronosaurus species, "K." boyacensis, was proposed by Hampe (1992) to include an Aptian skeleton from Colombia. He clarified that the inclusion of this new species in the genus is provisional because the definition of the genus Kronosaurus required a "mandatory revision". The holotype (and only known specimen) of "K." boyacensis, can only be observed dorsally, its skull is badly preserved due to intense crushing and the loss of bone surface, and its preparation is still insufficient. Probably for this reason Hampe's description of the skull is general and imprecise. However, Hampe (1992) justifies its provisional inclusion of the specimen in the genus Kronosaurus based on the large size of the skeleton, the short neck with 12 vertebrae, the expanded opisthotic, the small and unpaired supraoccipital, and the cervical rib facets extending through the entire length of the centra.
Kear (2003) provided "preliminary" diagnoses for the genus Kronosaurus and for the species K. queenslandicus. Both of the diagnoses include significant characters than cannot be observed in "K." boyacensis": in the definition of Kronosaurus, Kear (2003) includes premaxilla with four large caniniform teeth, anterior interpterygoid vacuity absent, ectopterygoid and pterygoid forming lateral flanges united in a short dished contact ventrolateral to the posterior interpterygoid vacuity, and mandibular symphysis extended back to sixth tooth position; and in the definition of K. queenslandicus he includes the presence of up to three dorsal longitudinal ridges in the interorbital region, pterygoids extensively underlapping basicranium and with interpterygoid vacuity situated beneath rather than anterior to the basicranium; paroccipital process contacting quadrate flange of pterygoid at lateral articulation only, and robust hyoids. All these characters are unknown in "K." boyacensis, so the inclusion of the holotype of "K." boyacensis within the genus Kronosaurus cannot be granted under the current knowledge of the specimen. Therefore, Kronosaurus is a genus that should be restricted to its type species, K. queenslandicus.
Unlike "K." boyacensis, the skull of the specimen MP111209-1 is well-exposed ventrally and can be compared with that ofK. queenslandicus. MP111209-1 shares with K. queenslandicus the large size, premaxilla with four caniniforn teeth, conical teeth without carinae, and 12 cervical vertebrae with single articular facets for ribs (Kear, 2003). Nevertheless, MP111209-1 differs from this species in a series of characters. It has a very short mandibular symphysis, ending in the middle of the fourth mandibular tooth, while in K. queenslandicus the symphysis encompasses six teeth (Kear, 2003); it exhibits an anterior interpterygoid opening in the palate, absent in K. queenslandicus; the three dorsal longitudinal ridges in the interorbital region mentioned by Kear (2003) are not present in MP111209 1; the hyoid bones, robust in K. queenslandicus (Kear, 2003), are slender in MP111209-1; the posterior dorsal vertebrae of MP111209-1 exhibit zygapophyses, which according to Kear (2003), do not seem to be present in K. queenslandicus. The anterior interpterygoid opening in MP111209-1 is small and could be related to a submature ontogenetic state, like Ketchum and Benson (2011a) established for Peloneustes. Instead, the difference in the number of teeth in the mandibular symphysis is a real trait of taxonomic value (Tarlo, 1960; Ketchum and Benson, 2011a; Knutsen, 2012; Benson et al. , 2013). Furthermore, the pineal foramen is located in a more posterior position in K. queenslandicus than in M111209-1, with its anterior end near the middle length of the orbits (Molnar, 1991; McHenry, 2009); the teeth ornamentation is marked in MP111209-1, even in the replacement teeth (Figure 4D), whereas in K. queenslandicus it is thinner and absent in the replacement teeth (Longman, 1924; While, 1935; McHenry, 2009); the dental sequence described by McHenry (2009) for K. queenslandicus, with two contiguous large dentary teeth followed by two adjacent large maxillary teeth (D4, D5, Mx1, Mx2) differs considerably from the condition seen in MP111209-1, where the dental sequence shows only two non-contiguous large teeth (D4, Mx1).
According to the reconstruction of K. queenslandicus presented by McHenry (2009), this species shows the suture between the premaxilla and the maxilla reaching the alveolar margin at the level of the fifth dentary alveolus, while this suture occurs at the level of the fourth dentary alveolus in MP111209-1. In addition, in the K. queenslandicus the vomer is not bifurcated posteriorly as it is in MP111209-1, the palatines join medially, while in MP111209-1 they are separated by the pterygoids, the palatal surface of the maxilla is reduced compared to the broad palatal surface of the maxilla exhibited in MP111209-1, the internal nostrils are in a more advanced position than they are in MP111209-1, and the palate is narrower anteriorly than in MP111209-1.
The new specimen and "K." boyacensis share a series of features. They share the conical teeth morphology, with circular cross section and with ornamented crowns, the short symphysis (although it seems to be shorter in MP111209-1), the number of cervical vertebrae and the long ischia. Although Hampe's (1992) diagnosis mentions five premaxillary teeth in "K." boyacensis (even though the suture between the premaxilla and the maxilla cannot be located with certainty), the location of the largest maxillary tooth in the snout of the holotype is similar to that of the first maxillary tooth of MP111209-1, suggesting that in fact "K." boyacensis may have four premaxillary teeth as in MP111209-1. Both specimens differ from the diagnosis given by Kear (2003) for Kronosaurus and K. queenslandicus in having shorter mandibular symphysis, ornamented tooth crowns and distinct dental sequence. Thus, considering the marked differences between K. queenslandicus and the new specimen, we propose to restrict Kronosaurus to its type species and erect a new genus for MP111209-1.
A comparison of MP111209-1 with "K." boyacensis is important given their morphological similarities and the fact that the genus attribution of the latter is dubious. Of the characters proposed by Hampe (1992) to define "K." boyacensis, several are not found in MP111209-1. These include the total number of teeth in each mandibular ramus, that in "K." boyacensis is approximately 24, while in MP111209-1 is not greater than 18 (although the number of mandibular teeth may differ among individuals of the same species (Ketchum and Benson, 2011a)); the diastema is broader in "K." boyacensis than in MP111209-1; the rib facets of the posterior cervical vertebrae are divided anteroposteriorly into two areas in "K." boyacensis, but not in MP111209-1, and the propodials are slender in "K." boyacensis compared with those of MP111209-1(table 2).
Furthermore, MP111209-1 is larger than "K." boyacensis; the length from the anterior tip of the rostrum to the femur insertion is 737 cm in MP111209-1 versus 630 cm in "K." boyacensis. However, "K." boyacensis has a larger skull in relation to the body than the new specimen. In fact, the length from the anterior tip of the rostrum to the femur insertion is 2.7 times the skull length measured in the midline in the holotype of "K." boyacensis, while it is 3.3 times in MP111209-1. The skull of "K." boyacensis is more elongated than that of MP111209-1; thus, the ratio between the length measured in the mid line and the maximum width measured at the level of the quadrates is more than 1.9 in "K." boyacensis (despite being crushed in its posterior region) while it does not exceed 1.7 in MP111209-1. The total number of pre-sacral vertebrae is 34 in "K." boyacensis (12 cervicals, 3 pectorals and 19 dorsals) (Hampe, 1992), and 37 in MP111209-1 (12 cervicals, 2 pectorals and 23 dorsals). The MP111209-1 scapulae are more robust than those of "K." boyacensis, according to the measurements given by Hampe, (1992); the glenoid ramus is notoriously shorter in MP111209-1 and the dorsal ramus is longer. Besides, in MP111209-1 the acetabular ramus of the ischium is short or poorly projected from the bone, while in the holotype of "K." boyacensis this ramus is appreciably longer. The ratio of humerus to femur length is greater in MP111209-1.
The morphology of the girdle bones and the propodials of MP111209-1 are more robust than those of "K." boyacensis. Although these differences may be related to sexual dimorphism, they may also be related to differences in ecological adaptations. Furthermore, the two specimens were found in different geological stages (MP111209-1: upper Barremian and "K." boyacensis: upper Aptian), and this, together with the combination of differences in the number of mandibular teeth, in the cranial proportions, and in the number of pre-sacral vertebrae, allows defining MP111209-1 as a taxon distinct from "K." boyacensis, probably belonging to the same genus.
In conclusion, the preceding comparisons show MP111209-1 as a brachauchenine pliosaurid with distinctive features that separate it from the other brachauchenine or pliosaurid genera. The new specimen shows sufficient morphological particularities to assign it to a new genus and species: Sachicasaurus vitae gen. et sp. nov. This new taxon represents the third pliosaurid genus defined for the Barremian of Colombia.
Phylogenetic analysis
The strict consensus of 20000 most parsimonious trees obtained after (TBR) branch swapping show S. vitae within a large polytomy that include all thalassophonean pliosaurids plus Marmornectes candrewi (Figure 8A). After applying the iter PCR algorithm, 13 unstable OTUs were identified: Pistosaurus skull; Stratesaurus taylori; Eoplesiosaurus antiquior; Meyerasaurus victor; QM_f51291; Acostasaurus pavachoquensis; Anguanax zignoi; Gallardosaurus iturraldei; Simolestes vorax; Liopleurodon ferox; Pliosaurus brachyspondylus; Pliosaurus irgisensis and Eromangasaurus australis. The unstable A. pavachoquensis was found jumping into different positions within suboptimal trees.
The resulting consensus tree obtained after removing the 13 unstable OTUs shows an unresolved Pliosaurus polytomy and a partially resolved monophyletic Brachaucheninae clade (Figure 8B). The Brachaucheninae node is supported by 10 synapomorphies, seven of which are present in MP111209-1: the parasphenoid is covered by the pterygoids, anterior to the posterior interpterygoid vacuities [86.0]; the palatine does not participate in the rim of the internal naris as seen in ventral view [93.1]; the pterygoid tapers anteriorly [95.0]; the mandibular symphysis tapers anteriorly in ventral view [113.0]; the long axis of the retroarticular process is inflected slightly posteromedially [123.1]; there is only one rib facet in the anterior-middle cervical vertebrae [160.3]; the cervical rib facets are located laterally [162.1]. The three other synapomorphies defining Brachaucheninae [163.1; 83.1 and 232.0] are unknown in MP111209-1.
The obtained tree shows the clade ((S. vitae + "K." boyacensis) + K. queenslandicus) separated from the Upper Cretaceous brachauchenines by having the anterior cervicals neural spines taller than long [171.0]. Within this clade, S. vitae was recovered as the sister taxon of "K." boyacensis, supporting the idea that both species could belong to the same genus, other than Kronosaurus, as suggested in the results of the morphological comparison. These two taxa have a reduced number of dorsal vertebrae [179.1], a synapomorphic character that separates them form K. queenslandicus.
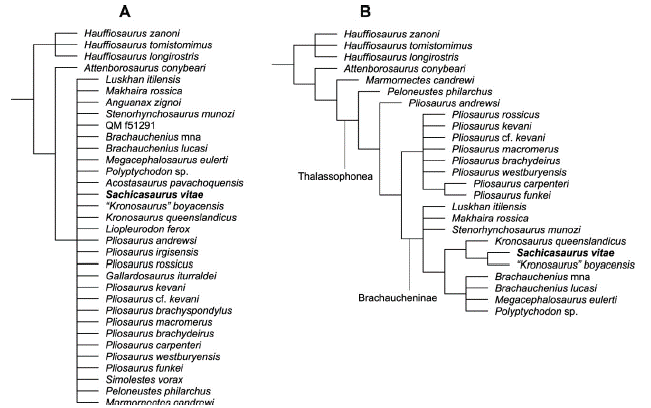
Figure 8 Pliosauridae branch of the phylogenetic tree obtained from the analysis of the complete Plesiosauria data set (see methods). A, Strict consensus tree of 20000 most parsimonious trees, 1579 steps, obtained after (TBR) branch swapping. B, reduced strict consensus tree after pruning the unstable OTUs (see text).
S. munozi, the other Colombian brachauchenine pliosaurid was recovered within an unresolved polytomy with L. itilensis and M. rossica in a basal position within Brachaucheninae. S. vitae and the other derived brachauchenines are distinguished from this basal polytomy by the absence of a postorbital-squamosal contact [42.1], the squamosal anterior extension significantly posterior to the postorbital bar [57.1], the parasphenoid terminating just anterior to the basisphenoid-basioccipital contact [84.1], the absence of foramina between the maxilla and the vomer, anterior to the internal naris [90.0]; four premaxillary teeth [131.0]; teeth with a rounded or sub-rounded cross-section [139.0]; and the presence of 15 or fewer cervical vertebrae [152.0].
Conclusions
The discussed comparisons and the results of the cladistic analysis show the specimen MP111209-1 as a derived brachauchenine pliosaur distinguished from others brachauchenines by a unique combination oftraits. The very anterior location of the pineal foramen, the extremely short symphysis (containing only three and a half alveoli), the low number of mandibular teeth, the anisodont dental sequence showing the first maxillary and the fourth dentary teeth as the largest of the series, the presence of ventral foramina and absence of dorsal ones on the cervical centra, and the number of presacral vertebrae (37), characterize MP111209-1 as a new genus and species, Sachicasaurus vitae gen. et sp. nov. Sachicasaurus represents the third pliosaurid genus defined for the Barremian of Colombia. Its discovery highlights the great diversity that pliosaurids attained during the Early Cretaceous and draw special attention to the study of Colombian Lower Cretaceous pliosaurids in the resolution of the phylogeny of Cretaceous pliosaurids. The occurrence of different genera of pliosaurids in the Barremian beds of Colombian suggests that the environmental conditions of the Colombian sea during the Early Cretaceous facilitated the development of sufficient marine life to sustain a diverse group of predators. More research is needed to reveal such a high diversity in the Cretaceous of Colombia.

