











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink1. Introduction
The knowledge of Lower Cretaceous pliosaurids has recently increased with the description of several specimens from the Berriasian-Hauterivian of western Russia (Fischer et al., 2015; Fischer et al., 2017; Zverkov et al., 2018), the Barremian-Aptian of Colombia (Gómez-Pérez & Noè, 2017; Páramo-Fonseca et al., 2016, 2018, 2019), the Aptian-Albian of Australia (Holland, 2018) and the Albian of Europe (Bardet et al., 2016; Madzia 2016; Madzia & Machalski, 2017). The Colombian pliosaurids are currently represented by four species, Acostasaurus pavachoquensis Gómez-Pérez and Noè (2017), known through a three-dimensionally preserved skull and some poscranial elements from the lower Barremian (Gómez-Pérez & Noè, 2017); Sachicasaurus vitae Páramo-Fonseca, Benavides-Cabra and Gutierrez (2018) known from a gigantic nearly complete skeleton from the upper Barremian (Páramo-Fonseca et al., 2018); Stenorhynchosaurus munozi Páramo-Fonseca, Gómez-Pérez, Noè and Etayo-Serna (2016) known from a nearly complete skeleton and a skull with some articulated cervical vertebrae from the upper Barremian (Páramo-Fonseca et al., 2016; 2019); and "Monquirasaurus" boyacensis (Hampe, 1992) represented by a nearly complete skeleton from the upper Aptian. This last species, previously assigned to Kronosaurus by Hampe (1992), has recently been attributed to a new genus Monquirasaurus Noè and Gómez-Pérez (2022); however, we consider that most of the characters used by these authors to differentiate this species from S. vitae are based on traits that are poorly preserved in the only known specimen of "M." boyacensis and are not obvious enough to warrant a generic distinction. Therefore, we prefer to use the name "Monquirasaurus" (within quotes) until new characters, such as the morphology of the palate, as discussed by Páramo-Fonseca et al. (2018), are available to be compared.
The specimens of Stenorhynchosaurus munozi (the holotype and the specimen MP050310-1) have the anterodorsal end of the snout lost or damaged (Páramo-Fonseca et al., 2016; 2019). A well-preserved fragment of the anterior end of a snout, housed in the Museo Paleontológico of the Universidad Nacional de Colombia at Villa de Leyva, was recently prepared. Based on the presence of diagnostic characters the specimen is referred to S. munozi. Herein, we describe this specimen and provide hitherto unknown features of the anterior end of the snout of S. munozi.
2. Materials and methods
The specimen described herein (MPVL-667) is an anterior end of a snout comprising occluded upper and lower jaws, which was collected by the staff of the Museo Paleontológico of the Universidad Nacional de Colombia at Villa de Leyva in the 1990s. It was collected as a trundled concretion found in the riverbed of La Yuca creek, in the municipality of Sutamarchán, Boyacá, Colombia, a municipality adjacent to Villa de Leyva. The discovery site is located approximately 8 km north-west of Villa the Leyva, (Fig. 1A). We visited the La Yuca creek and examined the concretions transported by the stream, both at the finding site and in the nearest 150 m upstream. Based upon a generic identification of ammonoids collected from the concretions, we propose the stratigraphic origin of the specimen MPVL-667. The ammonoid were identified at generic level by one of the authors (CDB-C).
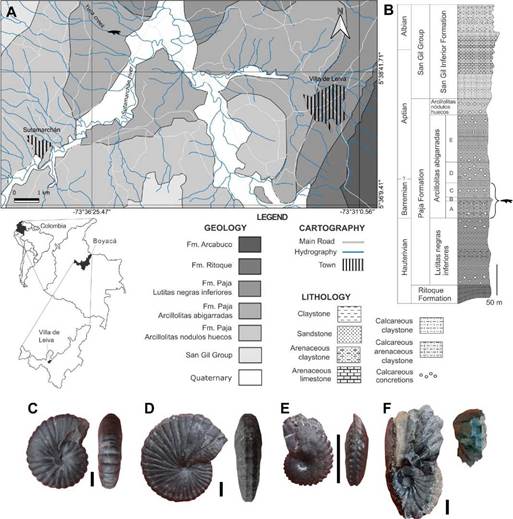
Figure 1 Geographic and stratigraphic provenance of the specimen MPVL-667. A, Geological map showing the finding site (pliosaurid silhouette) (modified from Etayo-Serna 1968). Cartographic base: Instituto Geográfico Agustín Codazzi. B, Generalized stratigraphic section of the Paja Formation cropping out around Villa de Leyva and Sutamarchán, showing the stratigraphic origin of the specimen (modified from Etayo-Serna, 1968). C-F, Lateral and ventral views of the ammonoid specimens representing the identified genera: C, Nicklesia Hyaat, 1903 (MP081022-1a), D, Pulchellia Uhlig, 1883 (MP081022-2), E, Heinzia sensu Bürgl (1956) (MP081022-7c) and F, Gerhardtia sensu Hyatt (1903) (MP081022-12). Scale bars C-F = 10 mm.
The specimen was embedded in micritic matrix, so chemical treatment was the selected preparation method. Successive immersions of 5-6 hours in formic acid at 3% concentration followed by a minimum of 12 hours of water rinsing and subsequent drying were performed. Three water baths of 24 hours each were performed at the end of the preparation to ensure the elimination of acid residues.
Given that the new specimen exhibits many morphological similarities with Stenorhynchosaurus munozi, the anatomical comparisons were focused on this species and specifically on specimen MP050310-1, which although damaged, preserves the same region of the snout. To corroborate the affinity of the new specimen with S. munozi, new observations were made directly on specimen MP050310-1. The anatomical description of the dentition follows the terminology proposed by Zverkov et al. (2018).
A 3D photogrammetric model of the specimen is provided as support of the description (see Supplemental Data 1). The model was obtained from raw 18 megapixels images using high-resolution structure from motion techniques. Each fragment was photographed with a Canon EOS rebel 4000D camera and processed individually using 275 pictures for the upper piece and 225 for the down one. Photos were taken surrounding each piece at three different angles to ensure the maximum possible resolution and coverage. Each model has been scaled using two scale bars located in each image. The correct three-dimensional position of both 3D models of the snout fragments was achieved manually, having single pictures of the right location of both pieces. Agisoft Metashape Professional 1.5.1, Meshlab v2022.02, and Blender 3.3.1 programs were used to obtain the 3D models.
A phylogenetic analysis was performed to determine the position of S. munozi within the Plesiosauria, based on our enhanced anatomical understanding of the species. The analysis was based on the data set of Páramo-Fonseca et al. (2019) which is a modified version of the data set of Fischer et al. (2017) and that of Benson and Druckenmiller (2014). The dataset was compiled using the Mesquite software 3.51 (Maddison and Maddison, 2018). The resulting data matrix has 102 OTUs and 271 characters (Supplemental Data 2). The Cladistic analysis was carried out using TNT (Goloboff and Catalano, 2016). All characters were treated as unordered and were equally weighted. An initial exploration for the shortest-length tree islands was performed using the new technology search options (Ratchet + Drift + Tree Fusing) with 200 ratchet iterations and 99,999 trees limit. Five replicates, one hit on the shortest tree, and seven trees held per replicate were obtained. The resulting trees were then used as the starting point for a tree bisection reconnection (TBR) branch swapping algorithm. A strict consensus was obtained, and the unstable OTUs were identified by applying the iterPCR algorithm (Pol and Escapa, 2009). The unstable OTUs were pruned from the consensus cladogram. The consistency (CI) and retention (RI) indices (Farris, 1989) were calculated, and Bremer support values (Bremer, 1994) were computed for the nodes in the consensus tree using TNT tools.
3. Geologic and biostratigraphic settings
The specimen was found in the riverbed of La Yuca creek, in a trundled concretion in which no ammonoids, except for a minute unidentifiable shell, were preserved. We collected a matrix sample and analyzed it for calcareous nannofossils and palynomorphs, but the analysis showed no microfossil content. Although no outcrops were found in the La Yuca creek, the stream cuts the La Yuca hill where the almost complete stratigraphic sequence of the Arcillolitas abigarradas Member of the Paja Formation outcrops (Figs. 1A-B). The topography and the geology of the finding region allowed us to establish with certainty that the specimen MPVL-667 comes from the Arcillolitas abigarradas Member of the Paja Formation.
Abundant trundled concretions bearing ammonoids were found in the riverbed of La Yuca creek. In some of the concretions we identified lower Barremian ammonoid genera, such as Nicklesia Hyaat, 1903 and PulchelliaUhlig, 1883 (Patarroyo, 2000; 2020) (Figs. 1C-D), and in other concretions we identified the genera Gerhardtia sensu Hyatt (1903) and Heinzia sensu Bürgl (1956) (Figs. 1E-F) which are indicators of the upper Barremian in Colombia (Patarroyo, 2000; 2020). We did not find any Aptian ammonoids on the creek, which suggests that the specimen came from Barremian rocks. Furthermore, in the nearest outcrops in the La Yuca hill we found the same ammonoid genera. In this context, we strongly assert that the specimen MPVL-667 originated within Barremian Beds of the Arcillolitas abigarradas Member that can be correlated with the segments A, B, and C of Etayo-Serna (1968) (see Fig. 1B). A more accurate stratigraphic origin of the specimen within the Barremian sequence is not possible.
4. Systematic paleontology

Type Specimen: VL17052004-1, Paleontological collection of the Servicio Geológico Colombiano. A nearly complete skeleton lacking part of the left anterior paddle, the complete right posterior paddle, and the tail (Páramo-Fonseca et al., 2016, Figs. 2-4).

Figure 2 MPVL-667, Stenorhynchosaurus munozi. The complete specimen before separation of the jaws, in A, left lateral, B, anterior and C, right lateral views. Abbreviations: d, dentary; mx, maxilla; pmx, premaxilla; 1d, first dentary tooth; 1pmx, first premaxillary alveolus. Scale bar = 50 mm.
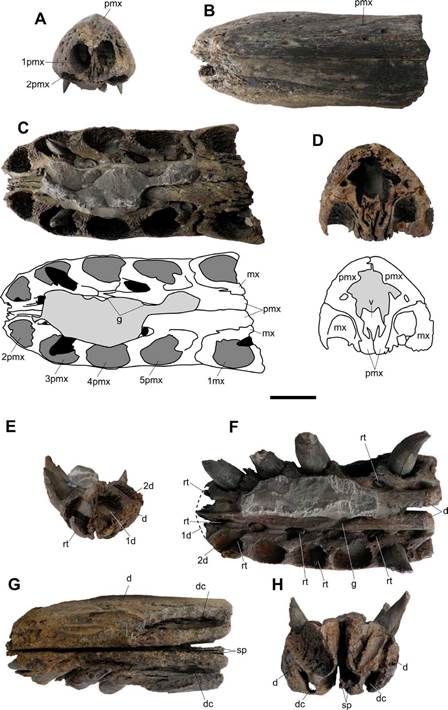
Figure 3 MPVL-667, Stenorhynchosaurus munozi. A-D, Upper jaw in A, anterior, B, dorsal, C, ventral (with interpretative scheme) and D, posterior (with interpretative scheme) views. Light gray: sediment; dark gray: alveoli; black: replacement teeth. E-H, Lower jaw in E, anterior, F, dorsal, G, ventral and H, posterior views. Abbreviations: d, dentary; dc, dentary canal; g, groove; mx, maxilla; pmx, premaxilla; rt, replacement tooth; sp, splenial; v, vomer; 1d, 2d, first, second dentary alveolus; 1mx, first maxillary alveolus; lpmx, 2pmx, ..., first, second, ... premaxillary alveolus. Scale bar = 30 mm.

Figure 4 MPVL-667, Stenorhynchosaurus munozi. Teeth. A-D, Replacement tooth into the third left premaxillary alveolus in A, labial, B, mesial, C, distolingual and D, apical views. E-G, sixth right mandibular tooth (incomplete by erosion) in E, lingual, F, mesial and G, distal views. H, cross section of the replacement tooth for the second left premaxillary position. Abbreviations: c, crown; ca, carina; r, root. Scale bar: A-G = 10 mm; H = 5 mm.
Referred specimen: MP050310-1, Paleontological collection of the Servicio Geológico Colombiano. A skull and the first 10 cervical vertebrae (Páramo-Fonseca et al., 2019, Figs 2-4)
New material: MPVL-667, Museo Paleontológico de Villa de Leyva, Universidad Nacional de Colombia. Anterior end of a snout with articulated mandible (Fig. 2).
Geographic and stratigraphic occurrence: Alto Ricaurte region (Villa de Leyva, Sáchica and Sutamarchán), Boyacá, Colombia; Barremian beds of the Arcillolitas abigarradas Member of the Paja Formation (Etayo-Serna, 1968; Páramo-Fonseca et al., 2016; 2019)
Emended Diagnosis: The diagnosis of Stenorhynchosaurus munozi follows Páramo-Fonseca et al. (2019) except for the following features, which are observed in the new specimen. Premaxilla with five (not four) functional alveoli, the anterior one procumbent and smaller than other premaxillary teeth. The autapomorphy concerning the vomer is specified as follows: vomer extending dorsally above the palatal processes of the premaxillae, but in ventral view ending posterior to the palatal premaxilla-maxilla suture.
5. Description
Preservational features
The specimen is three-dimensionally preserved and exhibits accurate detail of bone textures. It consists of the anterior end of a snout with the occluded mandible, extending to the level of the first maxillary alveolus (Fig. 2). The upper jaw was preserved slightly turned to its left side (Fig. 2B). The external surfaces of the first alveolus in the left premaxilla as well as the anterior tip and the right lateral side of the mandible are eroded. Most of the functional teeth are preserved in place on the right side of the mandible, whereas the remaining alveoli in the upper and lower jaws are either empty or they lodge replacement teeth.
6. Anatomical features
Premaxilla and maxilla. The anteriormost portion of the premaxillae, forming the anterior end of the snout, and a small portion of the anterior end of the maxillae are present in the specimen (Figs. 3A-D). The preserved portion of the snout is narrow with near parallel borders in dorsal view (Fig. 3B). Only a very slight narrowing of the snout at the level of the fifth alveolus and a faint widening at the level of the fourth alveolus are observed. The anterior tip of the snout is rounded and bears two procumbent alveoli (Figs. 2B; 3A-B). The suture between the premaxillae is straight. The external surface of the preserved portion of premaxillae is pitted by numerous vascular foramina, more densely arranged in the anterior end of the snout (Figs. 2A,C; 3B). The preserved portion of the lateral premaxilla-maxilla suture, near the alveolar margin, is interdigitated (Figs. 2A,C). Ventrally, the premaxilla-maxilla suture crosses the alveolar surface on a diastema of 19 mm in length that separates the fifth and sixth functional alveoli (Fig. 3C).
In ventral view, the palatal process of the premaxilla extends posterior to the premaxilla-maxilla suture (Fig. 3C). This condition indicates that the anterior end of the vomer exposed in palatal view is located posterior to the palatal premaxilla-maxilla suture. This trait is an autapomorphy of Stenorhynchosaurus munozi (Páramo-Fonseca et al., 2016). The palatal process of the premaxilla diverges dorsally forming a medial longitudinal lamella that contacts the vomer (Fig. 3D). Medial to the alveolar row, a narrow longitudinal canal runs, joining the concavities into which the replacement teeth erupt (Fig. 3C). This canal is interrupted in the diastema. At the level of the fourth alveolus an oblique posteromedial groove cross from the longitudinal canal to the midline where the two premaxillae meet (Fig. 3C).
Each premaxilla has five alveoli. The first alveolus is the smallest and the fourth alveolus is the largest (Fig. 3A, C). The first alveolus is practically horizontal and opens anteriorly on a vertical plane above the second alveolus, indicating a procumbent condition (Fig. 3A). The procumbent alveolus from both premaxillae is very close to each other; there is a thin bone wall of 0.26 mm separating them. The procumbent alveolus is vertically oval in outline, 19 mm in dorsoventral diameter and 13 mm in mesiodistal diameter (Fig. 3A). The subsequent premaxillary alveoli are circular in outline and anterolaterally oriented. Their mesiodistal diameter measure 22 mm (in the second alveolus), 23 mm (in the third), 25 mm (in the fourth), and 23 mm (in the fifth). The interalveolar spaces range from 6 to 8 mm. In the maxilla, only the right first maxillary alveolus is complete (Fig. 3C), it is circular in outline and its mesiodistal diameter is 22 mm.
Vomer. The vomers are fused forming an elongated bar that anteriorly extends dorsal to the palatal processes of the premaxillae (Fig. 3D). This dorsal extension of the vomers was not previously known in S. munozi; therefore, it is specified in the diagnosis of the taxon. The bar is trifurcated ventrally, forming three ventral lamellae that intercalate with the medial lamellae from the premaxillae, in such a way that, in cross-section, the joint between the vomers and the premaxillae is M-shaped (Fig. 3D). The dorsal surface of the fused vomers is concave and is textured by numerous longitudinal ridges.
Mandible. The preserved mandible includes the anterior portion of the dentaries and the anteriormost ends of the splenials, forming the anterior part of the mandibular symphysis (Figs. 3E-H). Most of the right external surface of the preserved mandible is eroded, but the left lateral surface is well preserved and demarcates the alveolar margins. As the rostrum, the mandible does not exhibit noticeable lateral expansion nor a marked increase in the size of the functional alveoli. The symphysis is not tightly closed; both mandibular rami run nearly parallel to each other, slightly further apart in the back (5 mm) than in the front (2 mm) (Figs. 3E, G-H). As in the upper jaw, a narrow longitudinal canal running medial to the alveolar row joins the concavities into which the replacement teeth erupt (Fig. 3F). Coinciding with the upper jaw, at the level of the fourth alveolus an oblique posteromedial groove cross from the longitudinal canal to the midline where both dentaries meet (Fig. 3F).
Each mandibular ramus preserves six complete and a seventh incomplete functional alveoli. The first functional alveolus is the smallest and the fifth is the largest (Fig. 3F). The first alveolus is anteriorly directed and nearly horizontal but opens in an anterodorsal plane. Differing from the first alveolus in the premaxilla, which opens directly above the second one, the anterior alveolus in the mandible does not open under the second alveolus, but anteroventral to it (Fig. 3E). The described condition suggests the first mandibular tooth was not procumbent. Except for the first alveolus, the mandibular functional alveoli are inclined slightly anterolaterally. The fifth and sixth mandibular teeth positions agree with the diastema of the upper jaw, without having an intercalated tooth. The measurements of the mesiodistal diameter of the mandibular alveoli are: 17 mm (anterior alveolus), 23 mm (second alveolus), 24 mm (third alveolus), 22 mm (fourth alveolus), 26 mm (fifth alveolus) and 22 mm (sixth alveolus). The space between successive alveoli in the mandible is slightly shorter than in the premaxilla; it ranges from 5 to 7 mm.
Teeth. Only replacement teeth are present in the premaxillae and maxillae (Fig. 3C). On the mandible, in addition to some replacement teeth, there are five functional teeth, four in the right mandibular ramus and one in the left (Fig. 3F). Replacement teeth emerge in small cavities placed within a canal that extends medially to the alveolar row. These cavities, named the replacement alveoli by Edmund (1960), are positioned distal-lingual to the marginal (functional) alveoli, and become elongated when the replacement tooth migrates to the marginal alveolus (Edmund, 1960; Sassoon et al., 2015). The replacement tooth of the first premaxillary position is located ventrally to the functional alveolus and the first replacement tooth in the mandible is placed dorsally to the marginal alveolus (Figs. 3C, F). In both premaxillae, the third replacement tooth is the largest; their crown is practically emplaced into the alveolus, but their root is still short.
The root in the second and third functional teeth of the right mandibular ramus is visible due to erosion (Fig. 2). They are 59 mm and 55 mm in heigh respectively and protrude 15 to 17 mm from the alveolar rim. The root height is about twice that of the crown. The root is slightly bulbous, circular in cross-section, and its surface is smooth.
The crowns of all functional teeth is poorly preserved and labially eroded (Fig. 2). However, in the right sixth functional tooth the lingual surface is visible and in the largest replacement teeth the crowns are well preserved. The crown of the largest replacement teeth is 31 mm in height. The crown is nearly conical, with a flat labio-mesial surface delimitated by two sharps smooth carinae, one mesial and one distal-labial, running the entire height of the crown (Figs. 4A-C). This morphology gives a sub-trihedral cross-section of the crown (Fig. 4H), as defined by Benson et al. (2013) for P kevani. An additional distal apicobasal ridge appears toward the apex of the crown forming a trihedral tip of the tooth (Fig. 4D), as in the holotype of Stenorhynchosaurus munozi (Páramo-Fonseca et al., 2016). The flat surface of the crown only bears a few fine ridges at its base, whereas on the remaining enameled surface there are coarse, abundant, and equidistant longitudinal ridges reaching the lower half of the crown (Fig. 4E-G). For every one or two ridges, a shorter inserted ridge is intercalating. These crown features have been described for the specimen MP050310-1 (Páramo-Fonseca et al., 2019).
7. Phylogenetic analysis
Based on the anatomical input provided by the described specimen we modify the scoring of character 131 of Stenorhynchosaurus munozi from state 0 (four premaxillary teeth) to 1 (five premaxillary teeth). Furthermore, we added one character to the matrix: 271- Direction towards which the anteriormost premaxillary alveolus faces. 0. The alveolus faces ventrally or weakly anteroventrally; 1. The alveolus faces directly anteriorly (procumbent). The scoring of the new character [271] can be found in Supplemental Data 3.
The strict consensus of 99,999 most parsimonious trees with 1620 steps (CI = 0.244 and RI = 0.670), obtained after TBR branch swapping, shows Stenorhynchosaurus munozi within a large polytomy that includes all pliosaurids and other plesiosaurs (Fig. 6A). After applying the IterPCR algorithm, 15 unstable OTUs were identified: Pistosaurus skull, Pliosaurus patagonicus, Makhaira rossica, Pliosaurus almanzaensis, QM_f51291, 'Polyptychodon interruptus'., Pliosaurus irgisensis, Pliosaurus brachydeirus, Pliosaurus brachyspondylus, Eromangasaurus australis, Stratesaurus taylori, Anningasaura lymense, Eoplesiosaurus antiquior, Eurycleidus arcuatus, and Atychodracon megacephalus. After pruning these 15 unstable OTUs from the consensus, the reduced strict consensus (Fig. 6B) shows S. munozi within a resolved Brachaucheninae. The results obtained here shows no changes with respect to those previously obtained by Páramo-Fonseca et al. (2019), even after including the new character. Nonetheless, the close relationship between S. munozi and L. itilensis is reflected in the obtained results and now it is supported by an additional character, the presence of procumbent anteriormost premaxillary teeth.

Figure 5 Stenorhynchosaurus munozi, specimen MP050310-1. A-B, Anterior tip of the snout in A, anterolateral and B, anterior views. C, detail of the anterior end of the snout showing the two concentric structures (broken teeth) found in the broken anterior surface. Scale bar = 30 mm.
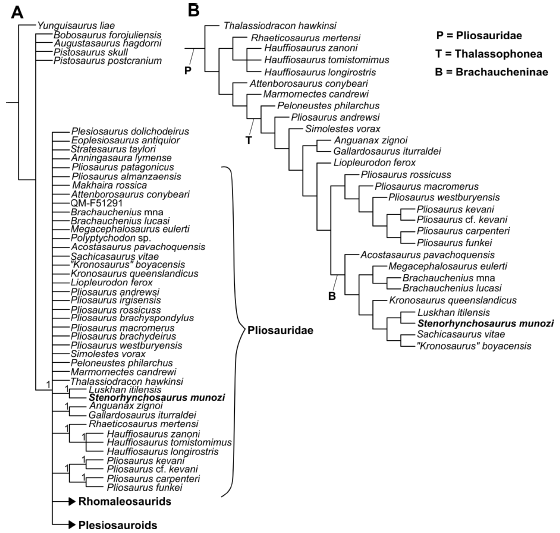
Figure 6 A, B, consensus trees of 99,999 trees of 1,620 steps based on data set of Plesiosauria with 102 OTUs and 271 characters (see Materials and Methods). A, strict consensus tree, with Bremer support indicated in some nodes. B, reduced consensus found after pruning the unstable OTUs.
The expected early diverging branch grouping Stenorhynchosaurus munozi, Makhaira rossica and Luskhan itilensis, does not appear in the cladogram, probably due to the unstable position of M. rossica. The node of S. munozi and L. itilensis does not appear as early-diverging as expected by their shared characteristics with the Jurassic thalassophoneans, instead it appears in a distant position to the ancestral node Brachaucheninae. This might be due to the unordered treatment of the characters related to the tooth morphology
8. Discussion
One of the two autapomorphies defining Stenorhynchosaurus munozi concerns the anterior region of the snout and is present in the new specimen: The anterior external exposure of the vomer is located posterior to the palatal premaxilla-maxilla suture in palatal view (Páramo-Fonseca et al., 2016; 2019). Moreover, the morphological features of the new specimen, including the size and shape of the snout, the size and orientation of the alveoli, and the shape and ornamentation of the teeth are consistent with the morphology previously described for Stenorhynchosaurus munozi (Páramo-Fonseca et al., 2016; 2019). The dimensions of the studied specimen agree with the size of the holotype of S. munozi, which was considered a juvenile individual (Páramo-Fonseca et al., 2016). In both specimens, the symphysis is not fused indicating that they represent juvenile individuals. The new specimen is thus reliably referred to as a juvenile individual of S. munozi.
The ventral exposure of the anterior end of the vomer is not observable in the only adult specimen of S. munozi known so far (Páramo-Fonseca et al., 2019). The anterior dorsal extension of the vomer observed in the new specimen raises the possibility that the shorter ventral exposition of the vomer in young specimens is the result of ontogenetic allometry. However, this doubt cannot be clarified until the ventral exposure of the vomer in the palate of an adult individual is known.
Stenorhynchosaurus munozi was erected for a nearly complete skeleton (VL17052004-1, the holotype) whose skull lacks the anterior tip of the snout (Páramo-Fonseca et al., 2016). A second specimen (MP050310-1), a skull with some articulated cervical vertebrae, was later described by Páramo-Fonseca et al. (2019). The dorsal region of the anterior end of the snout in the second specimen is damaged (Páramo-Fonseca et al., 2019: Figure 3C; Figs 5A-B). Therefore, the anterior dorsal end of the snout was unknown in S. munozi. In the published descriptions of these specimens, the authors proposed the number of premaxillary teeth of S. munozi as four based on the preserved material (Páramo-Fonseca et al, 2016; Páramo-Fonseca et al., 2019). The specimen herein described has five premaxillary teeth, the first being a dorsal procumbent tooth smaller than the other premaxillary teeth. This allows us to infer that S. munozi had five premaxillary teeth and not four as previously proposed.
New and detailed observations made on the dorsal end of the snout of the specimen MP050310-1, allowed us to identify two circular concentric structures on the broken surface that this specimen has above the alveolar margin (Fig. 5B-C). The morphology of these structures agrees with traces of two broken anterior procumbent teeth. Nevertheless, these structures seem to be in a somewhat higher position than that occupied by the procumbent alveoli in the new specimen. This discrepancy does not imply a taxonomic difference, since: 1- the broken surface where the concentric traces are registered in MP050310-1 is placed at a considerable distance from the frontal alveolar margin, so the traces could be showing the posterior portion of alveoli of the two slightly inclined procumbent teeth; and 2- the MP050310-1 represents an adult individual and the new specimen comes from a juvenile individual, so it could be expected an ontogenetic difference between the two specimens, with a more robust snout and procumbent teeth more separated from the alveolar margin in the adult.
Based on the above discussion, we can affirm that Stenorhynchosaurus munozi had five premaxillary teeth, the anterior one procumbent, nearly horizontal, and smaller than the other premaxillary teeth. The presence of procumbent premaxillary teeth is rare in thalassophonean pliosaurids; it has only previously been reported for the brachauchenine pliosaurids Makhaira rossicaFischer, Arkhangelsky, Stenshin, Uspensky, Zverkov, and Benson, 2015 and Luskhan itilensis Fischer, Benson, Zverkov, Soul, Arkhangelsky, Lambert, Stenshin, Uspensky, and Druckenmiller, 2017, both from the upper Hauterivian (Lower Cretaceous) of Russia (Fischer et al., 2015; Fischer et al., 2017). In addition, L. itilensis, M. rossica, and S. munozi share the elongated rostrum without lateral expansions, the isodont dentition, and the subtrihedral crown with trihedral apex (Fischer et al., 2015; Fischer et al., 2017; Páramo-Fonseca et al., 2019), a feature that differentiates these taxa from the other brachauchenine pliosaurids, which have a subcircular tooth cross-section (Zverkov et al., 2018). The differences between S. munozi and the Russian Hauterivian taxa have already been discussed by Páramo-Fonseca et al. (2019). Here we give only a few remarks on the number of premaxillary teeth (five in the S. munozi), which differs from that of the Russian taxa (six in M. rossica and seven or probably six in L. itilensis) (Fischer et al., 2015; Fischer et al., 2017; Zverkov, personal communication) and from that of all other brachauchenines (four functional teeth) (Schumacher et al., 2013; Albright et al., 2007; White, 1935; Gómez-Pérez and Noè, 2017; Páramo-Fonseca et al., 2018). The thalassophonean pliosaurids sharing five (or five-six) premaxillary teeth are the Jurassic taxa Liopleurodon feroxSauvage 1873, Pliosaurus macromerus (Phillips, 1871), Simolestes voraxAndrews, 1909 and Eardasaurus powelliKetchum & Benson, 2022 from England and Pliosaurus rossicusNovozhilov, 1948 from Russia. Nevertheless, all these taxa differ from S. munozi by having a constricted rostrum with large teeth near the premaxilla-maxilla suture and short symphysis (Andrews, 1909; 1913; Halstead, 1971; Benson et al., 2013; Fischer et al, 2015, appendix data set; Ketchum & Benson, 2022). The combination of features in the premaxillary dentition of S. munozi is therefore unique among thalassophonean pliosaurids. The new traits are therefore added to the diagnosis of the species.
The presence of anterior procumbent teeth in the premaxilla of the Barremian taxon S. munozi from Colombia has significant implications in the temporal distribution of this trait in the history of long-snouted brachauchenines. It is noteworthy that within the thalassophonean pliosaurids the procumbent teeth in front of the upper jaw only occur in the long and narrow snouted taxa of the Early Cretaceous. These taxa have morphological affinities with the Cretaceous brachauchenine pliosaurids but retain traits of the dental crown morphology found in the Jurassic thalassophonean pliosaurids. These long and narrow snouted taxa with procumbent teeth formed, early in the history of the brachauchenines, a derived evolutionary lineage with a particular ecological adaptation that successfully expanded across the Tethys Sea from Eurasia to northern Gondwana during the Early Cretaceous.
The apparent temporal discontinuity of the ecological adaptation of a narrow snout with procumbent front teeth on premaxilla among brachauchenines could be due to a collection bias, but also to ecological changes. It would not be unreasonable to suggest that the polycotylid plesiosaurs, having elongated snouts, relatively isodont dentition, and frequent teeth with carinae (Fischer et al., 2018), which begin their history in the Aptian-Albian (Druckenmiller, 2002; Kear, 2003) and become diverse during the late Cretaceous (Druckenmiller and Russel, 2009; Fischer et al., 2018; Sachs et al., 2020), could have occupied the ecological niche of the Early Cretaceous long-snouted brachauchenines.
9. Conclusions
We identify the specimen MPVL-667 as a juvenile individual of Stenorhynchosaurus munozi by presenting juvenile traits in a morphology consistent with that described for this species and showing the anteriorly exposed end of the vomer in palatal view located posterior to the palatal premaxilla-maxilla suture, which is one of the autapomorphies that define S. munozi. The study of the new specimen reveals information about traits that were previously unknown or were interpreted differently for the species, such as the dorsal anterior extension of the vomer, the presence of five premaxillary teeth (previously interpreted as four) and the existence of a procumbent anteriormost premaxillary tooth. This last feature is corroborated by new observations in the specimen MP050310-1. The procumbent anterior premaxillary teeth and the subtrihedral crown with trihedral apex is shared by S. munozi, Luskan itilensis, and Makhaira rossica and differentiate these taxa from the other brachauchenines. These shared traits show a close relationship between the Hauterivian brachauchenines from Russia and the Barremian brachauchenines from Colombia. Moreover, our cladistic analysis supports the phylogenetic proximity of S. munozi and L. itilensis by 11 synapomorphies, including the new character of the procumbent anteriormost premaxillary teeth. We suggest that the niche occupied by the Early Cretaceous long-snouted brachauchenines (S. munozi, L. itilensis, and M. rossica) could have been later occupied by the polycotylid plesiosaurs, which also have elongated snouts, relatively isodont dentitions and frequent teeth with carinae.

