










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAntipoda. Revista de Antropología y Arqueología
Print version ISSN 1900-5407
Antipod. Rev. Antropol. Arqueol. no.13 Bogotá July/Dec. 2011
TAFONOMÍA DE HUESOS DE AVES. ESTADO DE LA CUESTIÓN Y PERSPECTIVAS DESDE EL SUR DEL NEOTRÓPICO
Isabel Cruz*
* Doctora en Arqueología, Universidad de Buenos Aires, Argentina. isabelcruz55@yahoo.com.ar Universidad Nacional de la Patagonia Austral Santa Cruz, Argentina
RESUMEN
Las investigaciones en el sur del Neotrópico señalan que el impacto de los procesos tafonómicos sobre los restos de aves varía en relación con el tamaño corporal y la morfoanatomía de las mismas, así como en función del contexto ecológico y ambiental considerado. Los estudios futuros deberán enmarcarse en análisis comparativos para evaluar la acción de diversos procesos sobre los restos de diferentes vertebrados en un mismo contexto ambiental, y también deberán desarrollarse observaciones controladas para profundizar algunos temas.
PALABRAS CLAVE:
Tafonomía de vertebrados, aves, meteorización, representación anatómica, modificaciones por carnívoros.
BIRD BONES TAPHONOMY. STATE OF THE PROBLEM AND PERSPECTIVES FROM THE SOUTHERN NEOTROPICS
ABSTACT
Current studies on bird bones conducted at the Neotropics show that differences in the impact of taphonomic processes are mainly due to body size and anatomy, as well as to the role of environmental and ecological contexts. Future research must include comparative studies and controlled observations to generate specific models that will help to understand taphonomic histories of zooarchaeological remains in the region.
KEY WORDS:
Vertebrate Taphonomy, Birds, Weathering, Anatomic Representation, Carnivore's Modifications.
TAFONOMIA DE OSSOS DE AVES. ESTADO DA QUESTÃO E PERSPECTIVAS DESDE O SUL DO NEOTRÓPICO
RESUMO
As pesquisas no sul do Neotrópico sinalizam que o impacto dos processos tafonô micos sobre os restos de aves varia em relação ao tamanho corporal e a morfoanatomia das mesmas, assim como em função do contexto ecológico e ambiental considerado. Os estudos futuros deveriam moldar-se em análises comparativos para avaliar a ação de diversos processos sobre os restos de diferentes vertebrados num mesmo contexto ambiental e também deveriam desenvolver-se observações controladas para aprofundar alguns temas.
PALAVRAS CHAVE:
Tafonomia de vertebrados, aves, meteorização, representação anatô mica, modificações por carnívoros.
Introducción
Las aves presentan características que las convierten en instrumentos interesantes para analizar varios aspectos de la ecología humana. En primer lugar, constituyen un recurso que puede ser utilizado integralmente: su carne y sus huevos son un alimento importante, que contiene gran cantidad de proteínas y ácidos grasos. Además, sus plumas y huesos pueden ser utilizados como materia prima para la confección de diversos artefactos. En segundo lugar, las aves pueden ser importantes proxys en la determinación de variables paleoecológicas (Emslie, 1995; Morales Muñiz, 1993; Savanti, 1994, entre otros). Muchas aves están estrechamente asociadas a una comunidad vegetal o a un hábitat específico. La presencia de sus huesos en un depósito arqueológico puede ser una línea más en el análisis paleoambien-tal, que necesariamente debe integrar información de varios campos. Debido a que no todos los huesos de aves presentes en estos depósitos son producto de la predación humana, también permiten abordar aspectos relacionados con la formación del registro arqueológico, así como la distribución de especies presentes en un ambiente y su uso por los predadores no humanos.
Durante los últimos años ha aumentado el interés en desarrollar este potencial a partir del estudio de los restos de aves provenientes de depósitos zooarqueológicos en diversos lugares de la región Neotropical (Corona Martínez, 2001, 2002; Cruz, 2007a; deFrance, 2005; Higgins, 1999; Lefévre, 1989, 1992, 1997; Mameli, 2002; Martínez et al, 2009; Savanti, 1994; Tivoli, 2010, entre otros). A pesar de ello, todavía es poco lo que conocemos acerca de las aves mediante el registro arqueológico en la región. Uno de los factores que han impedido lograr avances es que sus huesos pueden ser muy escasos en gran parte de los sitios arqueológicos. En virtud de que es poco lo que sabemos acerca de la forma en que se preservan sus restos, es difícil dilucidar si esta escasez se debe a que eran poco cazadas o a que sus huesos fueron destruidos por procesos naturales antes de poder incorporarse a los registros arqueológicos.
La disciplina científica que se dedica a comprender cómo se conservan o destruyen los restos orgánicos es la tafonomía. Los estudios tafonómicos constituyen un paso previo al análisis de los restos zooarqueológicos, ya que brindan las herramientas interpretativas adecuadas para que las inferencias acerca del pasado sean correctas. El objetivo de este trabajo, por lo tanto, es sintetizar las investigaciones tafonómicas de restos de aves efectuadas hasta el momento en Patagonia (ver la figura 1), con el fin de evaluar el grado de conocimiento alcanzado y establecer las bases para las investigaciones futuras. La Patagonia se ubica en el sur del Neotrópico (región biogeográfica que abarca desde México hasta Tierra del Fuego), y muchas de las consideraciones presentadas aquí son de utilidad para toda la región. Sin embargo, tal como se enfatizará al final del trabajo, será necesario que en el futuro se desarrollen investigaciones específicas en distintos ambientes de esta región, que completen los estudios presentados aquí y muestren la diversidad ecológica que presentan los procesos tafonómicos en diferentes localidades de la región. Esto permitirá contar con un conjunto amplio de principios tafonómicos que podrá ser utilizado por todos aquellos que quieran abordar el estudio de las aves a partir de registros óseos (tanto arqueológicos como paleontológicos).
Zooarqueología y tafonomía
Desde sus inicios como disciplina científica, la zooarqueología ha estado íntimamente ligada al desarrollo de los estudios tafonómicos (Andrews, 1990; Binford, 1981; Brain, 1981; Blumenschine, 1986; Gifford, 1981; Lyman, 1994; Pickering et al, 2007, entre otros). La tafonomía es el estudio de los procesos de preservación y de la forma en que éstos afectan a la información que puede obtenerse a partir del registro fósil, tanto paleontológico como arqueológico (Behrensmeyer et al., 2000).
Como el registro arqueológico es el parámetro a partir del cual se determina la relevancia de las explicaciones acerca del pasado humano (Binford, 1981), gran parte de la tarea de los arqueólogos consiste en establecer la historia de formación de los depósitos. Una vez conocida esta historia -o, al menos, la parte de ella accesible a los investigadores-, entonces se puede proceder a evaluar la forma en que el registro en cuestión permite avalar o descartar las hipótesis acerca de la conducta humana. Desde esta perspectiva, el análisis zooarqueológico no sólo implica estudiar los depósitos en cuanto a los actores humanos, sino que también es necesario entender cuál pudo ser la contribución de otros actores y procesos en su formación. No hacerlo implica correr el riesgo de asignar contenido cultural a características de los restos óseos que son producto de procesos naturales.
En el marco de la zooarqueología, uno de los objetivos de los estudios tafonómicos es generar instrumentos interpretativos que permitan garantizar las inferencias sobre la conducta humana que se basan en los restos de animales recuperados en contextos arqueológicos. Las observaciones actuales son un medio fructífero para comprender la manera en que diversos actores y procesos afectan a los restos orgánicos, ya que permiten establecer cuáles son las relaciones que existen entre procesos geobiológicos específicos y los conjuntos de huesos que son contemporáneos a los mismos (Binford, 1981; Gifford, 1981, entre otros).
A lo largo de las últimas décadas se han multiplicado las observaciones tafonómicas en el sur del Neotrópico. Sin embargo, ni en esta región ni en otros lugares del mundo los estudios destinados a los restos de aves han sido tan exhaustivos como los dedicados a los mamíferos (Behrensmeyer et al, 2003; Bickart, 1984; Cruz, 2007b; Davis y Briggs, 1998; Livingston, 1989; Muñoz y Savanti, 1998; Oliver y Graham, 1994, entre otros). Las investigaciones tafonó-micas sobre huesos de aves en el sur del Neotrópico comenzaron a mediados de la década de 1990 y se desarrollaron fundamentalmente en ambientes de la Patagonia. Aunque todavía escasas, estas investigaciones han permitido conocer la forma en que diversos procesos afectan a los restos de estos vertebrados, así como algunos de sus efectos en la formación de los conjuntos óseos recuperados de depósitos arqueológicos.
El esqueleto de las aves y los procesos tafonómicos
La interacción entre huesos y procesos tafonómicos está mediada por varios factores, entre los cuales se destacan las propiedades específicas de los huesos de cada grupo de organismos. El esqueleto de las aves difiere del de otros vertebrados en varios aspectos, especialmente los relacionados con la principal adaptación locomotora del taxón: el vuelo. El modo de locomoción plantea constricciones anatómicas y, aunque no todas las aves son voladoras, muchos aspectos corporales de las aves reflejan los especiales requerimientos que demanda el vuelo (Livingston, 1989).
Los huesos de cualquier vertebrado tienen una función, que se relaciona con su "fuerza tafonómica" (Lyman, 1994). La función anatómica de un hueso determina sus propiedades mecánicas, las que a su vez median sobre los efectos que los procesos tafonómicos tendrán sobre ese hueso. Muchas de estas propiedades de los huesos pueden ser tafonómicamente significativas: el diseño y el tamaño corporal, así como la microestructura del tejido óseo, son factores que influirán en la destrucción o preservación de los restos de cada taxón (Andrews, 1990; Behrensmeyer, 1991; Blumenschine, 1987; Kidwell y Behrensmeyer, 1988; Lyman, 1994, entre otros). En el caso de las aves, entre los requerimientos para el vuelo que tienen correlatos importantes en el esqueleto se cuentan una reducción general del peso, la obtención de mayor rigidez corporal y una mayor resistencia para sobrellevar el stress que implica el vuelo (Bellairs y Jenkin, 1960; Feduccia, 1992; Gilbert et al., 1981). Debido a ello, varios investigadores han planteado la necesidad de efectuar estudios tafonómicos específicos para sus restos (Cruz, 2000a, 2007b; Davis y Briggs, 1998; Higgins, 1999; Lyman, 1994; Trapani, 1998, entre otros).
Como grupo, las aves exhiben pocas diferencias osteológicas entre sí (Bellairs y Jenkin, 1960), pero presentan gran variabilidad con respecto al tamaño. Éste es un factor que debe ser considerado al analizar el impacto de los procesos tafonómicos sobre sus huesos, porque se ha demostrado que procesos tafonómicos como la meteorización, la acción de carnívoros o el pisoteo varían según el tamaño corporal de los animales implicados (Andrews, 1990; Behrensmeyer, 1978; Behrensmeyer y Dechant-Boaz, 1980; Behrensmeyer et al, 2003; Blumenschine, 1987).
Por otro lado, en relación con el vuelo, muchos de los huesos del cráneo y del esqueleto poscraneano de las aves son huecos y contienen sacos de aire (Bellairs y Jenkin, 1960; Higgins, 1999). El grado de neumatización varía entre taxones: es mínimo en las aves más pequeñas, mientras que involucra la mayoría de los huesos de las aves grandes y activamente voladoras, como los albatros y las águilas. También entre los Ratites, aves no voladoras, los huesos presentan un alto grado de neumatización, característica posiblemente derivada de sus ancestros adaptados al vuelo. En cambio, los huesos de las aves buceadoras están muy poco neumatizados y sus esqueletos reflejan el equilibrio entre los requerimientos del vuelo y del buceo (Gilbert et al, 1981).
En síntesis, en función de las necesidades que plantea el vuelo, las aves han desarrollado un esqueleto más liviano y rígido, con menor cantidad de elementos individuales que el de otros vertebrados. Además, también la microestructura y la morfología de los huesos han sufrido modificaciones. Todos estos aspectos tienen particulares implicaciones para la tafonomía de sus huesos y permiten predecir que los procesos tafonómicos no los afectarán de la misma manera que a los de otros vertebrados.
aspectos metodológicos
Todos los estudios tafonómicos dedicados a los restos de aves efectuados en Patagonia se desarrollaron en el marco de lo que Marean (1995) denominó tafonomía naturalista. A diferencia de la tafonomía experimental, el enfoque naturalista permite conocer las condiciones variables en las que se conforman los registros óseos derivados de comunidades específicas, en ambientes concretos. Para ello, las investigaciones no se realizan en laboratorios o ambientes controlados sino que se basan en la observación de los restos depositados naturalmente. De esta manera, es posible establecer nexos entre los procesos tafonómicos y sus trazas en los restos.
Tal como lo efectuaron otros investigadores en diferentes ecosistemas de África (Behrensmeyer, 1993; Behrensmeyer y Dechant-Boaz, 1980; Behrensmeyer et al, 2003; Blumenschine, 1989; Bunn et al, 1991; Sept, 1994; Tappen, 1995, entre otros), la metodología utilizada en Patagonia consistió en el registro y posterior análisis de todas las características de los restos óseos actuales a lo largo de unidades espaciales amplias, por medio de la implementación de transectas en diferentes sectores del paisaje (Belardi, 1999; Cruz, 1999, 2000b, 2004, 2007b; Muñoz y Savanti, 1998, entre otros).
Las observaciones se centraron en aquellas aves de tamaño mediano a grande, que son las de posible utilidad económica para las poblaciones humanas del pasado (ver la figura 2). Con excepciones que luego se detallan, la mayor parte de estas aves están incluidas en lo que Behrensmeyer (1991) considera como microvertebrados, es decir, con un peso menor a 5 kg. Dentro del marco de este rango de tamaño corporal, las observaciones diferenciaron por lo menos tres grupos de aves en función de su morfoanatomía. El fundamento de esta diferenciación es que las características anatómicas tienen correlatos en la constitución del esqueleto y, por lo tanto, pueden ocasionar diferencias en el potencial de preservación de huesos correspondientes a distintas arquitecturas corporales.

En función de esta premisa, durante las observaciones efectuadas en Patagonia se consideraron las características que presentaban los restos de aves voladoras, de ñandúes (Rheidae) -aves no voladoras del grupo de las Ratites- y de pingüinos (Spheniscidae) -aves marinas no voladoras-. Las anatomías de estas aves presentan marcadas diferencias entre sí y sus pesos varían entre los 27 kg de los ñandúes, los 3,5-4 kg de los pingüinos y la diversidad de pesos (entre unos pocos cientos de gramos hasta los 4 kg) de las aves voladoras consideradas en la región.
Los resultados obtenidos
Las investigaciones desarrolladas tanto en el sur del continente (Belardi, 1999; Cruz, 1999, 2000b, 2007b, entre otros) como en la Isla Grande de Tierra del Fuego (Muñoz y Savanti, 1998) permitieron conocer diversos aspectos relacionados con la depositación, modificación y destrucción de huesos de aves por los procesos tafonómicos en diferentes ambientes (la costa marina, la estepa y el bosque andino-patagónico). Muchos de estos resultados son concordantes con los obtenidos en otros lugares del mundo (Behrensmeyer et al, 2003; Bic-kart, 1984; Davis y Briggs, 1998; Oliver y Graham, 1994) pero también mostraron diferencias en algunos aspectos (Cruz, 2007b, 2008).
Como en otros estudios actualísticos, las investigaciones patagónicas se centraron en dos aspectos relacionados con la preservación de restos orgánicos: la depositación y las modificaciones que se producen antes de que los mismos sean cubiertos por sedimentos (sensu Behrensmeyer et al., 2000). Esto tiene sentido si se considera que muchos de los factores que determinan las condiciones bajo las que se preservarán algunos organismos -o partes de los mismos- se definen en el momento de la muerte y/o en un corto lapso después de ella (Binford, 1981; Andrews, 1990; Behrensmeyer, 1991; Lyman, 1994, entre otros). Conocer la dinámica relativa a estos dos aspectos bajo condiciones ambientales particulares es un medio para establecer cuántos y cuáles podrán ser los restos que se preservarán o los que, por lo menos, tendrán posibilidades de ser cubiertos por los sedimentos.
Aunque los temas abordados por estos estudios han sido muchos, aquí se sintetizan los resultados alcanzados en relación con algunos de ellos: la depo-sitación de restos por causas naturales y su posible impacto sobre los registros arqueológicos de la región, la secuencia de desarticulación y la representación anatómica de aves, la meteorización y la acción de los carnívoros. Todos estos temas se consideran particularmente relevantes para una adecuada interpretación de los restos de aves contenidos en depósitos arqueológicos, para entender los contextos en los que los mismos se produjeron y como un instrumento más entre los varios necesarios para generar expectativas acerca de los registros óseos derivados de la actividad humana.
La depositación de huesos por causas naturales
Una de las formas más evidentes en que los procesos naturales pueden afectar a los registros arqueológicos es mediante la acumulación de restos en lugares recurrentemente utilizados por los humanos, que forman palimpsestos en los que se mezclan huesos depositados por distintos procesos y actores.
Binford (1981) define "integridad" como la homogeneidad relativa de los agentes causantes de la acumulación de los materiales presentes en un depósito arqueológico. Una herramienta útil para evaluar la integridad de los registros arqueofaunísticos en diferentes puntos del espacio y establecer las posibilidades de "contaminación" con huesos depositados por causas no humanas es conocer la dinámica que determina la depositación natural de restos en un ambiente dado. En un mismo paisaje existen variaciones en la depositación de restos (Behrensmeyer y Dechant-Boaz, 1980; Cruz, 2004, 2009a; Sept, 1994; Tappen, 1995, entre otros). La posibilidad de que exista superposición entre artefactos u otros restos arqueológicos y huesos derivados de muertes naturales variará en función de las características del registro arqueológico y de la forma que asuma la depositación natural de restos (Borrero, 2001).
En Patagonia, la mayor diferencia en este sentido se encuentra entre los hábitats costeros y los del interior (tanto sectores de estepa como de bosque y humedales). En los hábitats costeros se producen grandes concentraciones de restos durante el período de nidificación y cría de las aves marinas, cuando la mortalidad es generalmente muy alta (Cruz, 1999, 2007b, 2009a). Dadas las características que presenta el registro arqueológico en muchos sectores costeros de la región, las probabilidades de "contaminación" de depósitos arqueológicos por la muerte natural de aves son muy altas. Como se ha planteado para varios casos en Patagonia continental, es posible que todos o gran parte de los restos de aves relacionados con un registro arqueológico particular sean producto de la depositación natural, debido a la dinámica de las colonias de nidificación (Cruz, 1999, 2004, 2006, 2009a).
En cambio, en la estepa y el bosque característicos del sur de Patagonia las posibilidades de superposición o mezcla con restos arqueológicos son muy bajas, ya que, si se compara con lo que ocurre en la costa, son muy pocos los huesos de aves que se depositan, y, además, en virtud de la intensidad de los procesos tafonómicos, es factible que gran parte de ellos sean destruidos antes de ser cubiertos por sedimentos (Cruz, 2000b, 2007c, 2008, 2009a). La excepción detectada la constituyen los abrigos rocosos, que son sectores de gran acumulación natural por la acción de los carnívoros y por muertes naturales (Cruz, 2000a, 2000b, 2007a, 2009a). Los huesos de aves voladoras son frágiles y su destrucción puede ser rápida si no son cubiertos por sedimentos poco tiempo después de la depositación. Los abrigos rocosos constituyen puntos en el paisaje que generalmente presentan altas tasas de sedimentación o condiciones que atemperan la acción de los procesos destructivos, lo que facilita la conservación de huesos de aves. Como muchos de los depósitos arqueológicos en Patagonia continental se ubican en abrigos rocosos, el potencial para la mezcla con restos derivados de muertes naturales es muy alto en estos loci.
En síntesis, conocer la forma general que adopta la depositación natural de restos en un ambiente dado puede ser una herramienta útil para modelar la integridad de los depósitos arqueológicos y para generar los medios que permitan evaluarla en cada caso.
secuencia de desarticulacion
Otro de los temas al que se le ha prestado especial atención en la tafonomía de aves es la secuencia de desarticulación. Este interés ha sido importante en los estudios tafonómicos de otros vertebrados y se vincula con el hecho de que la representación de partes esqueletarias, utilizada frecuentemente para efectuar inferencias acerca del comportamiento humano, puede estar relacionada con esta secuencia. Si bien la desarticulación en sí misma no implica la destrucción o desaparición de los elementos, aumenta la vulnerabilidad de los huesos frente a la acción de otros procesos.
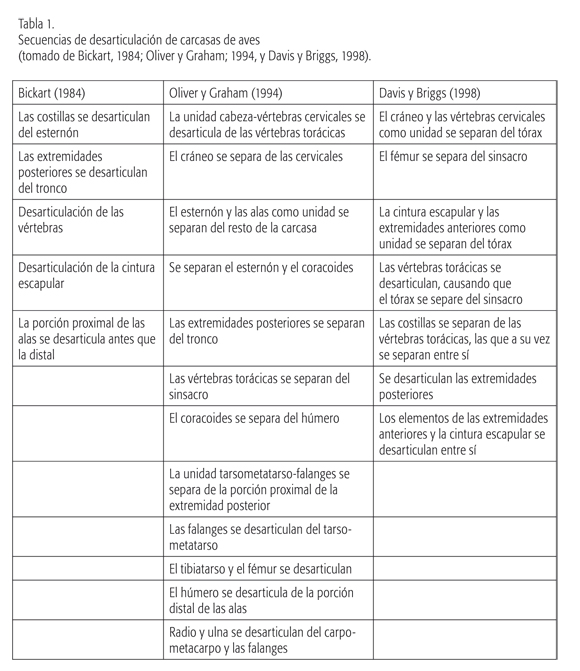
La secuencia de desarticulación de aves voladoras sigue un patrón que, en líneas generales, ha sido registrado mediante observaciones actuales en diferentes lugares del mundo (ver la tabla 1). Entre los aspectos más recurrentes, se ha observado que las extremidades posteriores se desarticulan de la carcasa antes que las anteriores. Asimismo, los elementos de las extremidades posteriores se desarticulan entre sí más rápidamente que los de las extremidades anteriores. Estas últimas permanecen más tiempo como una unidad, frecuentemente junto a coracoides, fúrcula, escápula y, en algunos casos, esternón. En ambas extremidades, las porciones proximales se desarticulan antes que las distales. Estos aspectos de la secuencia planteada para carcasas de aves voladoras son coincidentes con la detectada en Patagonia, y también se cumplen en los restos de pingüinos (Cruz, 2007b).
En las carcasas de ñandúes, en cambio, los primeros elementos en desarticularse son el cráneo, la cintura escapular, el esternón y las extremidades anteriores, que son rápidamente destruidos por los procesos tafonómicos. Vértebras y costillas pueden permanecer articuladas como una unidad en conjunto con la pelvis, pero se desarticulan y destruyen antes que las extremidades posteriores. Estas extremidades, especialmente la porción distal (tibiatarso, metatarso y dígitos), pueden permanecer articuladas por largo tiempo y sus elementos son los más resistentes a los procesos destructivos (Belardi, 1999; Cruz, 2007c; Cruz y Elkin, 2003; Cruz y Fernández, 2004).
Estas diferencias en las secuencias de desarticulación pueden correlacionarse con diferencias en la preservación de partes esqueletarias de las distintas aves patagónicas. En efecto, aquellos elementos que se desarticulan primero son los que posiblemente se destruyan primero. Los elementos correspondientes a las extremidades anteriores de las aves voladoras, que permanecen articulados por más tiempo, son los que posiblemente se conserven mejor (ver la figura 3) en la superficie terrestre. En el caso de los ñandúes, los elementos que perdurarán más tiempo articulados son los de las extremidades posteriores, por lo que serán los últimos en destruirse y los que, por lo tanto, tendrán más posibilidades de preservarse tanto en contextos naturales como arqueológicos.

Representación de partes esqueletarias
La frecuencia de partes esqueletarias es una de las variables que permiten discutir la historia tafonómica de un conjunto óseo (Behrensmeyer, 1991) y es uno de los medios más utilizados por los arqueólogos para abordar diversos aspectos de la subsistencia humana. Debido a ello, muchos de los estudios tafonómicos se han dedicado muy especialmente a establecer las diferencias que existen entre el patrón natural de representación anatómica y el derivado de la acción humana (Cruz, 2005; Ericsson, 1987; Livingston, 1989, entre otros).
Las investigaciones efectuadas dejaron en claro que la representación de partes esqueletarias de aves es el resultado de un conjunto complejo de procesos, que varía en función de gran número de variables (tamaño de las presas, tipo de predador implicado, si constituyen restos de comida o son restos regurgitados, etc.). Incluso, tal como ha planteado Bovy (2002) al respecto, la variabilidad puede extenderse a los conjuntos generados por actividades humanas, ya que pueden existir diferencias en función del tipo de aprovechamiento efectuado.
Entre los logros más importantes de las investigaciones en Patagonia, se cuenta el haber definido patrones naturales de representación anatómica diferentes para cada uno de los tipos de aves considerados. Entre los restos de aves voladoras predominan los elementos correspondientes a las extremidades anteriores (especialmente húmeros, radios y ulnas), las vértebras, así como los tibiatarsos (Cruz, 2005; Muñoz y Savanti, 1998). En el caso de los ñandúes, los elementos más representados son los de las extremidades posteriores (tibiatar-sos y tarsometatarsos) y las vértebras (Belardi, 1999; Cruz, 2000, 2005; Cruz y Elkin, 2003; Cruz y Fernández, 2004). Los restos de pingüinos más abundantes son los correspondientes a ambas extremidades (especialmente húmero, fémur y tibiatarso) y el coracoides (Cruz, 1999, 2005, 2007b; Emslie, 1995). La cintura pélvica es la porción anatómica menos representada en todos los casos.
En síntesis, la representación de partes esqueletarias en conjuntos naturales no es azarosa, sino que presenta patrones consistentes. Las diferencias detectadas no pueden atribuirse a las respectivas historias tafonómicas: en todos los casos estos patrones son el resultado de la acción conjunta de varios procesos tafonómicos (principalmente, meteorización y acción de carnívoros) que facilitan la destrucción diferencial de elementos. Tampoco reflejan un problema de muestreo, ya que se puso especial cuidado en relevar hábitats específicos de cada una de las aves consideradas. Contrariamente a lo planteado por algunos investigadores, las investigaciones patagónicas mostraron que estos patrones son un correlato de la morfoanatomía de cada una de las aves y no se relacionan con el agente causal de la depositación (Cruz, 2005).
El caso de los ñandúes constituye un ejemplo de aplicación de estos resultados: el patrón natural observado en los estudios tafonómicos es coincidente con la abundancia anatómica detectada en varios sitios arqueológicos de Patagonia y de todo el sur de Sudamérica en general. Las primeras interpretaciones de esta representación de partes esqueletarias proponían que la misma era el resultado del descarte de las extremidades posteriores en los sitios de caza, debido a que sólo se transportaban hasta los campamentos aquellas porciones con mayor contenido cárnico, que es variable entre elementos esqueleta-rios (Giardina, 2006). Sin embargo, esta representación anatómica también es característica de los conjuntos naturales y puede explicarse como el resultado de la destrucción mediada por la densidad (Cruz y Elkin, 2003; Fernández et al, 2001, entre otros). Por lo tanto, la abundancia anatómica de los conjuntos arqueológicos no debería atribuirse directamente a un comportamiento humano específico (el transporte de partes de una presa y el descarte de otras), ya que también puede ser el resultado de un gran número de procesos, en los que la destrucción mediada por la densidad mineral tiene el papel preponderante. Si bien las investigaciones acerca del aprovechamiento del ñandú por las poblaciones prehistóricas aún deben profundizarse, los estudios tafonómicos tuvieron un rol central para descartar una hipótesis y señalar nuevas líneas de investigación, las cuales deberán incluir explicaciones más complejas acerca de la representación de partes esqueletarias que presentan los restos de estas aves en los conjuntos arqueológicos de la región.
Meteorización
La meteorización es uno de los principales procesos que causan la destrucción de restos óseos en la superficie terrestre (Behrensmeyer, 1978; Lyman, 1994). Los huesos de distintos animales presentan diferentes tasas de meteorización en un mismo ambiente (Gifford, 1981), y el tamaño corporal es uno de los factores determinantes para que esto ocurra (Behrensmeyer, 1978). Debido a las características de los huesos de aves y a su tamaño corporal generalmente pequeño, el impacto de la meteorización sobre ellos es muy diferente al que sufren los huesos de otros animales, especialmente los mamíferos. Por otro lado, como la meteorización es un proceso ecológicamente mediado, son necesarias investigaciones específicas para comprender la forma en que puede afectar los depósitos arqueológicos en un ambiente particular.
Las primeras investigaciones en Patagonia dedicadas a los restos de aves fueron las de Muñoz y Savanti (1998) en la Isla Grande de Tierra del Fuego. A partir de sus observaciones, estos investigadores formularon varios estadios de meteorización para los huesos de aves que, en líneas generales, seguían los linea-mientos propuestos por Behrensmeyer (1978) para los huesos de mamíferos de más de 5 kg. Estudios posteriores efectuados en Amboseli (África) corroboraron que los estadios inicialmente propuestos para los mamíferos también son útiles para evaluar la meteorización en huesos de aves (Behrensmeyer et al., 2003).
A partir del perfil de meteorización de huesos de aves derivado de los resultados de sus observaciones, Muñoz y Savanti (1998) plantearon que el predominio de los estadios más bajos se debe a que las propiedades mecánicas de estos huesos facilitan su destrucción antes de alcanzar una meteorización avanzada. Estos resultados fueron confirmados por los estudios en el sector continental de Patagonia (Cruz, 2000a, 2007b, 2008, entre otros) y son acordes con los derivados de las investigaciones en Amboseli (Beherensmeyer et al, 2003). En todos los casos, la acción de la meteorización provocó que los huesos de aves desaparecieran más rápidamente que los de mamíferos, especialmente de ungulados. Existe un aspecto, sin embargo, en el que los resultados de Patagonia y Amboseli difieren: la destrucción de los huesos de aves producida por la meteorización es más rápida en Patagonia (Cruz, 2008). Esto es acorde con lo registrado para los huesos de ungulados en la región (Borrero, 2007), por lo que debe considerarse una consecuencia de las condiciones ambientales prevalecientes en el sur del continente.
Un último punto por destacar se relaciona con las diferencias registradas entre los huesos de los distintos grupos de aves patagónicas con respecto a la meteorización. La comparación entre los perfiles de meteorización de huesos de aves voladoras y pingüinos en Patagonia permite afirmar que las diferencias que pueden existir en la estructura de sus huesos no son importantes frente a la acción de este proceso tafonómico (Cruz, 2007b). En cambio, se registraron diferencias al respecto en el caso de los ñandúes, ya que todas las observaciones destacan que los elementos correspondientes a las extremidades posteriores de estas aves tienen mayor resistencia a los procesos tafonómicos, incluida la meteorización (Belardi, 1999; Cruz, 2007a; Cruz y Fernández, 2004). Esta resistencia puede atribuirse a dos factores: 1) los ñandúes son las aves de mayor tamaño corporal en Patagonia, y 2) estas aves están adaptadas para correr a grandes velocidades, por lo que los huesos de sus extremidades posteriores -especialmente tibiatarso, tarsometatarso y dígitos- poseen paredes gruesas y altos valores de densidad mineral (Cruz y Elkin, 2003). Como consecuencia, parte del esqueleto del ñandú (especialmente las extremidades anteriores, el cráneo y el esternón) comparte con el de las aves voladoras una rápida destrucción por la meteorización, mientras que las extremidades posteriores se asemejan a los huesos de ungulados, que poseen una mayor resistencia a este proceso (Cruz, 2007c).
Modificaciones y transporte por carnívoros
A diferencia de lo registrado en otros ecosistemas (Binford, 1981; Blumens-chine, 1989; Tappen, 1995, entre otros), los carnívoros patagónicos no tienen gran potencial para destruir huesos y carcasas de ungulados (Borrero, 1990, 2000; Cruz, 2008, entre otros). Sin embargo, en función de su pequeño tamaño corporal y su particular estructura ósea, las aves constituyen presas cuyos huesos y carcasas pueden ser profundamente afectados por la acción de predadores y carroñeros (Bickart, 1984; Oliver y Graham, 1994).
Las observaciones efectuadas en Patagonia permitieron establecer que el impacto de la acción de los carnívoros varía según las características de las comunidades de aves y las condiciones ecológicas imperantes en distintos ambientes de la región. Esto implica que para una adecuada evaluación del impacto que puede tener este proceso sobre los restos de aves hay que considerar los contextos en que se producen la predación y el carroñeo. La costa marina presenta marcadas diferencias con los otros ambientes en lo que respecta a sus comunidades de aves, lo cual determina el grado en el que las modificaciones producidas por los carnívoros influyen en la conformación del registro (Cruz, 2004, 2007b, 2008; Muñoz y Savanti, 1998). En la costa, la gran mortalidad de aves concentrada espacial y temporalmente se expresa en una gran deposita-ción de restos, que constituyen una importante fuente de alimento disponible para los carnívoros locales. Entonces, si bien podría esperarse una gran destrucción inicial en función del tamaño corporal de las aves, las condiciones de abundancia de carcasas permiten la preservación de gran parte de los restos. En la estepa y el bosque, en cambio, las aves son menos abundantes y las carcasas disponibles son escasas y están muy dispersas, por lo que los carnívoros las aprovechan más intensamente. Como parte de este aprovechamiento integral, se produce una mayor destrucción de las carcasas, y, además, el transporte de algunas partes anatómicas por los predadores afecta la localización de los huesos de aves (Cruz, 2000b, 2007c, 2008).
Los autores que han efectuado observaciones sobre la forma en que los carnívoros modifican los huesos y carcasas de aves concuerdan en que existen diferencias según se trate de predadores mamíferos o aves (Davis y Briggs, 1998; Oliver y Graham, 1994). Los mamíferos generalmente producen muchas modificaciones y destrucción y pueden transportar los restos fuera del sitio de muerte. En cambio, en general las aves carnívoras tienen una menor capacidad de modificación de los huesos y, si ocurre transporte, el mismo es de carne, vísceras o partes anatómicas segmentadas.
En Patagonia, aves y mamíferos también tienen diferentes roles en la modificación de huesos y carcasas (Cruz, 1999, 2004, 2007b). En la costa marina se ha registrado poca acción de los mamíferos terrestres sobre las carcasas de aves; en cambio, es importante la acción de las aves, que generalmente se centra en los tejidos blandos. Tal cual se observó en otros lugares del mundo, su papel como actores tafonómicos consiste fundamentalmente en contribuir a la desarticulación de las carcasas. Sin embargo, en un área de nidificación de cormoranes emplazada en una isla en la que no habitan mamíferos se registraron restos de aves con modificaciones en el esternón (Cruz, 2009b). Estas modificaciones, similares a los punctures producidos por mamíferos y descritos por Binford (1981), fueron efectuadas por gaviotas (Larus dominicanus) o eskúas ( Catharacta skua), e indican que en algunos casos las aves pueden producir efectos sobre los huesos.
En la estepa patagónica, en cambio, son los mamíferos quienes suelen destruir, modificar y transportar carcasas y huesos de aves (Belardi, 1999; Cruz, 2000a; Muñoz y Savanti, 1998). El transporte se produce principalmente a sus madrigueras de cría, que en este ambiente suelen ubicarse en abrigos rocosos (Cruz, 2000a; Fernández et al., 2010; Martín, 1998). No todos los huesos de aves en estas madrigueras presentan modificaciones claramente asignables a carnívoros, por lo que, si en el abrigo existen depósitos arqueológicos, estos restos pueden incorporarse a los mismos sin trazas que permitan establecer su procedencia (Fernández y Cruz, 2010).
Como en el caso de la meteorización, existen diferencias entre las distintas aves en lo que respecta a las modificaciones por estos actores tafonómicos. Los huesos de los ñandúes, en función de su tamaño y estructura, son más resistentes a la acción de los carnívoros. Los daños registrados no afectan a todo el esqueleto, sino que se ubican principalmente en las epífisis de huesos largos, el esternón y cintura pélvica (Belardi, 1999) (ver la figura 4). La destrucción de las carcasas de aves voladoras y pingüinos, en cambio, puede ser más completa, y los huesos con más probabilidades de sobrevivir son los correspondientes a las extremidades anteriores y la porción distal de las posteriores (Cruz, 2000a, 2005, 2007b).

Aplicación a casos zooarqueológicos
Robert Blumenschine (1998) planteó que la habilidad de los arqueólogos para "leer" el registro arqueofaunístico de manera confiable y verificable no está a la altura de la creatividad alcanzada por las explicaciones acerca del pasado. Para este investigador, el desarrollo desigual de estos dos aspectos de las investigaciones zooarqueológicas no se debe a la carencia de principios y modelos tafonómicos, ya que afortunadamente los mismos son muchos y variados. La causa de esta inexplicable falta de profundización de los aspectos tafonómicos involucrados en la interpretación arqueológica es que muchos zooarqueólogos todavía se niegan a aplicar los modelos de esta disciplina en sus interpretaciones de los conjuntos que describen (Blumenschine, 1998). En los comienzos de la segunda década del siglo XXI, esto continúa ocurriendo en gran parte de las investigaciones en la región Neotropical. Por lo tanto, además del ejemplo mencionado en relación con el aprovechamiento de Rheidos en el sur de Sudamérica (ver "Representación de partes esqueletarias"), aquí presentaremos brevemente dos casos que permiten apreciar el potencial de los modelos tafo-nómicos para una adecuada interpretación zooarqueológica.
El primer caso se vincula con la necesidad de evaluar si los restos de aves recuperados en varios sitios arqueológicos (CCP 5, CCP 7, ADG y AGV) en abrigos rocosos ubicados en el sector cordillerano del sur de Patagonia continental en Argentina derivan de la actividad humana o son el producto de procesos tafonómicos, como la acción de los carnívoros (Cruz, 2000a, 2007a). Teniendo en cuenta las altas posibilidades de superposición de huesos derivados de muertes naturales y restos arqueológicos en los abrigos rocosos (ver "La depositación de huesos por causas naturales" y "Modificaciones y transporte por carnívoros"), se planteó la necesidad de efectuar un análisis tafo-nómico que incluyese diversas líneas de evidencia para evaluar las hipótesis planteadas acerca de la explotación de aves en este sector cordillerano. El énfasis de este análisis estuvo puesto en la caracterización de la acción de los carnívoros en el área (Cruz, 2000a) y en las propiedades del registro óseo de los sitios arqueológicos de la localidad (Cruz, 2000b, 2007a). Por ejemplo, en varios de los conjuntos zooarqueológicos se registraron huesos de carnívoros sin modificaciones antrópicas, por lo que es posible pensar que estos animales murieron naturalmente en el abrigo y que debieron tener un papel en la acumulación y modificación de los restos recuperados (Cruz, 2000b). En el sitio CCP 5, se determinaron huesos de un cánido -posiblemente zorro extinto (Dusicyon avus)- y de puma, además de la presencia de marcas atribuibles a carnívoros en 3,4-6,6% de los especímenes de artiodáctilos identificados. En CCP 7, ADG y AGV también se registraron restos de carnívoros entre los materiales en estratigrafía. Estos restos incluyen tanto huesos como excrementos, y posiblemente corresponden a cánidos (Cruz, 2007a). Además, en la actualidad, las aves rapaces ocupan varios de estos abrigos en los que se emplazan sitios arqueológicos (CCP 5, CCP 7 y ADG), y su presencia en las secuencias arqueológicas ha sido demostrada a partir de egagrópilas y plumas, de restos de sus nidos y de sus propios huesos. En síntesis, la evidencia indica que los carnívoros (tanto mamíferos como aves) utilizaron los abrigos rocosos durante el período de formación de los depósitos arqueológicos. El análisis de las escasas modificaciones culturales en los restos de aves y la información contextual de los mismos en los conjuntos arqueológicos permitieron determinar que los dos actores más importantes en la conformación del conjunto avifaunístico son los carnívoros y los humanos. Tanto unos como otros contribuyeron a la formación del mismo, aportando y modificando los huesos de aves, lo cual fue tenido en cuenta en las interpretaciones arqueológicas. En síntesis, este caso mostró la necesidad de evaluar todos los posibles actores tafonómicos involucrados en la formación de los conjuntos óseos, para ajustar las interpretaciones y corroborar adecuadamente las hipótesis acerca de la forma en que los humanos predaron las aves en este sector de la cordillera de los Andes (Cruz, 2007a).
El segundo caso muestra los beneficios de comparar la representación natural de partes esqueletarias observada mediante estudios tafonómicos de huesos actuales de cormoranes (Phalacrocorax sp.) con la registrada en depósitos arqueológicos que incluyen restos de estas aves (Cruz, 2009b). A partir de las observaciones actuales, se determinó que el perfil de representación anatómica de los cormoranes se destaca por el predominio de elementos de las extremidades posteriores (mayoritariamente, fémures y tibiatarsos) y anteriores (especialmente, húmeros). Esta diferencia con el perfil anatómico de aves voladoras (ver "Representación de partes esqueletarias") se explica por la carencia de neumatización en muchos de los huesos de cormoranes y el papel de las extremidades posteriores de estas aves en la locomoción bajo el agua, por lo que sus huesos son más resistentes a los procesos tafonómicos que los correspondientes a los de otras aves voladoras (Cruz, 2009b). El análisis tafo-nómico estuvo destinado, entre otros temas, a comparar esta representación natural con la determinada en varios conjuntos zooarqueológicos que tienen una alta representación de los cormoranes. Tres de estos conjuntos provienen de Patagonia: Cabo Blanco 1 (Moreno et al, 1998), Punta Baja (Lefévre, 1989) y Túnel VII (Mameli, 2002); el restante, de la costa peruana: Quebrada Tacahuay (de France, 2005). La única similitud detectada en la representación anatómica de todos los conjuntos (los arqueológicos y el natural) fue que la cintura pélvica es la unidad menos representada. El perfil de Punta Baja se asemeja en líneas generales al natural, pero con una alta representación del esqueleto axial que no posee el conjunto natural (Cruz, 2009a). Los otros conjuntos tienen representaciones anatómicas muy diferentes a las registradas a partir de los estudios tafonómicos. Lo más interesante de esta comparación es que no fue posible aislar un patrón común a todos los depósitos arqueológicos. Lo mismo sucede con otros conjuntos de Patagonia, como los analizados por Lefévre (1992, 1997) y Savanti (1994). Esto no implica que el patrón detectado en cada uno de los depósitos no sea el resultado de la acción humana, lo cual debería determinarse en cada caso mediante un análisis tafonómico exhaustivo. Lo que estas diferencias señalan es que es posible una gran variabilidad en la representación de partes esqueletarias de cormoranes en los conjuntos zooarqueológicos, relacionada con la diversidad de actividades humanas que pueden involucrar a estas aves. Muchas de estas actividades no dejan una firma específica en los registros óseos y pueden asemejarse al perfil natural observado (tal como sucede con el conjunto de Punta Baja). Dada esta ambigüedad, el análisis tafonómico permitió concluir que los patrones de representación de partes esqueletarias no pueden considerarse un indicador claro de los actores y procesos involucrados, por lo que es poco aconsejable que se efectúen afirmaciones sobre la conducta humana utilizando exclusivamente estos perfiles anatómicos. Sin embargo, la representación de elementos constituye una de las variables que permiten discutir la historia de un conjunto óseo (Behrensmeyer, 1991), y su análisis en el marco de las investigaciones zooarqueológicas permitirá efectuar estudios tafonómicos más precisos en cuanto a los procesos involucrados.
Las investigaciones futuras
Las investigaciones tafonómicas efectuadas hasta el momento son un primer paso para comprender las particularidades del proceso de reciclado de huesos de aves en el sur del Neotrópico. Son varias las líneas de investigación sobre tafonomía de los restos de aves que quedan abiertas y que deberán desarrollarse para completar los resultados que presentamos aquí.
En primer lugar, es necesario profundizar las investigaciones acerca de la relación entre morfología esqueletaria y resistencia a los procesos tafonómicos, especialmente entre diferentes especies de aves voladoras. Esta categoría abarca un amplio rango de tamaños corporales y de variación anatómica, dentro del marco general del esqueleto básico de las aves. La anatomía de muchas de estas aves está también influenciada por aspectos independientes de la locomoción, e incluso por modos de locomoción alternativos. En Patagonia se ha avanzado en este aspecto, iniciándose investigaciones en colonias de nidificación de cormoranes (Phalacrocoracidae). Como se mencionó anteriormente, a partir de estos estudios, se pudo establecer que la representación de partes esqueletarias de estas aves, que vuelan y bucean, se asemeja más a la de los Spheniscidae que a la de otras aves voladoras (Cruz, 2009b). En el futuro, las investigaciones deberían explorar éste y otros aspectos en los restos de aves con diferencias anatómicas y tamaños corporales marcados.
En segundo lugar, los estudios tafonómicos de aves deben enmarcarse en investigaciones más amplias, destinadas a establecer el impacto que tienen diversos actores y procesos tafonómicos sobre los restos de distintos vertebrados en un mismo contexto ambiental. Esto permitirá evaluar el rol de las diferencias anatómicas más amplias en la preservación de restos de distintos organismos y puede constituir un instrumento adecuado para el análisis de conjuntos zooarqueológicos, generalmente conformados por restos de diferentes taxones.
Además, algunos aspectos requieren ser tratados mediante observaciones controladas. Entre ellos, se destacan la secuencia de desarticulación y el impacto de la meteorización. Estos temas han sido tratados por medio de las observaciones efectuadas, pero la rápida destrucción de los restos de aves en contextos naturales hace necesario que se implementen metodologías específicas que permitan un control más estricto de las variables que harán posible comprenderlos más acabadamente.
Por último, sería interesante contrastar los principios derivados de estas investigaciones tafonómicas en una escala más amplia, que permita aumentar los contextos ambientales en los que se depositan y modifican los restos de aves. La dinámica tafonómica de ambientes más cálidos, con otras relaciones ecológicas o con mayor diversidad de aves, seguramente es diferente a la registrada en el extremo sur del Neotrópico. Esta comparación más amplia permitirá una comprensión más acabada del comportamiento tafonómico de los restos de aves y será de utilidad para interpretar depósitos en ésta y otras regiones biogeográficas.
Consideraciones finales
Todo el conocimiento acerca del pasado está fuertemente mediado por la comprensión de los factores tafonómicos que han actuado para conformar los registros fósiles que son la base para las interpretaciones científicas (Behrensmeyer et al., 2000). El entendimiento amplio de la forma en que operan los mecanismos que promueven la destrucción y/o preservación de restos orgánicos permite generar los modelos e hipótesis que constituyen el corpus teórico a partir del cual se pueden interpretar los casos específicos.
El principal aporte de las observaciones tafonómicas en Patagonia, o en cualquier otra región del Neotrópico, radica en que pueden dar cuenta de condiciones diferentes a las consideradas por los modelos y principios que actualmente constituyen la base de la disciplina, formulados a partir de observaciones en otros lugares del mundo. Cuanto mayor sea la cantidad de ecosistemas y ambientes en los que se evalúen los modelos tafonómicos, mayor podrá ser su grado de generalidad, debido a que habrán sido puestos a prueba bajo gran variedad de condiciones. Al mismo tiempo, las investigaciones en diferentes regiones del mundo permitirán desarrollar modelos más específicos, en virtud de que los ya existentes podrán ser ajustados para dar cuenta de situaciones particulares. En este sentido, las observaciones patagónicas permitieron, por un lado, mostrar la persistencia de los patrones regulares propios de los procesos considerados, que se expresan independientemente de las distintas condiciones ambientales. Por otro, posibilitaron establecer cuáles son las particularidades propias de la región y, de esta manera, comprender cuáles son algunos de los aspectos que pueden incidir en las variaciones.
Como señaló Binford (1981), conocer la dinámica de las poblaciones humanas del pasado es imposible sin comprender el registro arqueológico. La tafonomía, como parte de las investigaciones arqueológicas, brinda la posibilidad de interpretar adecuadamente una porción del mismo -el registro arqueofaunístico-, al tiempo que permite abordar aspectos de los contextos en los cuales se generaron los depósitos. Dados la madurez de la disciplina y el desarrollo que la misma ha alcanzado en todo el mundo, esperamos que en los próximos años la tafonomía pueda incorporarse definitivamente en las investigaciones zooarqueológicas del Neotrópico.
Agradecimientos
Este trabajo es un resultado de los proyectos UNPA 29A/260-1 y PIP/CONI-CET 112-200801-00996. .
Referencias
Andrews, Peter
1990. Owls, Caves and Fossils. Chicago, The University of Chicago Press.
Behrensmeyer, Anna K.
1978. "Taphonomic and Ecological Information from Bone Weathering", Paleobiology 4, pp. 150-162.
1991. "Terrestrial Vertebrate Accumulations", en Taphonomy: Releasing the Data Locked in the Fossil Record, eds. P. A. Allison y D. E. G. Briggs, pp. 291-335. Nueva York, Plenum Press. [ Links ]
1993 "The Bones of Amboseli. The Taphonomic Record of Ecological Change in Amboseli Park, Kenya", en National Geographic Research & Exploration 9, pp. 402-421. [ Links ]
Behrensmeyer, Anna K. y Dorothy E. Dechant-Boaz
1980. "The Recent Bones of Amboseli Park, Kenya, in Relation to East African Paleoecology", en Fossils in the Making. Vertebrate Taphonomy and Paleoecology, eds. Anna K. Behrensmeyer y Andrew P. Hill, pp. 72-92. Chicago, University of Chicago Press.
Behrensmeyer, Anna K., Susan M. Kidwell y Roger Gastaldo
2000. "Taphonomy and Paleobiology", en Deep Time. Paleobiology's Perspective, Suplemento de Paleobiology, eds. Douglas H. Erwin y Scott L. Wing, 26, pp. 103-147.
Behrensmeyer, Anna K., C. Tristan Stayton y Ralph E. Chapman
2003. "Taphonomy and Ecology of Modern Avifaunal Remains from Amboseli Park, Kenya", Paleobiology 29, pp. 52-70.
Belardi, Juan Bautista
1999 "Hay choiques en la terraza. Información tafonómica y primeras implicaciones arqueofaunísticas para Patagonia", en Arqueología. Revista de la Sección Prehistoria 9, pp. 163-185.
Bellairs, A. D'A. y C. R. Jenkin
1960. "The Skeleton of Birds", en Biology and Comparative Physiology of Birds, ed. A. J. Marshall, Vol. 1, pp. 241-300. Nueva York, J. Wiley & Sons.
Bickart, Karl J.
1984. "A Field Experiment in Avian Taphonomy", Journal of Vertebrate Paleontology 4, pp. 525-535.
Binford, Lewis R.
1981. Bones. Ancient Men and Modern Myths. Nueva York, Academic Press.
Blumenschine, Robert J.
1986. Early Hominid Scavenging Opportunities. Implications of Carcass Availability in the Serengeti andNgorongoro Ecosystems. Oxford, BAR International Series 283.
1987. "Characteristics of an Early Hominid Scavenging Niche", Current Anthropology 28, pp. 383-407. [ Links ]
1989. "A Landscape Taphonomic Model of the Scale of Prehistoric Scavenging Opportunities", Journal of Human Evolution 18, pp. 345-371. [ Links ]
1998. "Comments to Musterian Large-Mammal Remains from Kobeh Cave: Behavioral Implications (C. Marean y S. Y. Kim)", Current Anthropology 39 (Suplemento), pp. 79-113. [ Links ]
Borrero, Luis A.
1990. "Taphonomy of Guanaco Bones in Tierra del Fuego", Quaternary Research 34, pp. 361-371.
2000. "Ten Years After: esquema para una tafonomía regional de la Patagonia Meridional y norte de Tierra del Fuego", en Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia, tomo I, pp. 183-193. Río Gallegos, Universidad Nacional de la Patagonia Austral. [ Links ]
2001. "Regional Taphonomy: Background Noise and the Integrity of the Archaeological Record", en Ethnoarchaeology of Andean South America. Contributions to Archaeological Method and Theory, ed. Lawrence A. Kuznar, pp. 243-254. Ann Arbor, Michigan. International Monographs in Prehistory. Ethnoarchaeological Series 4. [ Links ]
2007. "Longitudinal Taphonomic Studies in Tierra del Fuego, Argentina", en Taphonomy and Archaeozoology in Argentina, eds. María A. Gutiérrez, Gustavo Barrientos, Mónica Salemme, Laura Miotti y Guillermo L. Mengoni Goñalons, pp. 219-233. Oxford, BAR (British Archaeological Reports) International Series S1601. [ Links ]
Bovy, Kristine M.
2002. "Differential Avian Skeletal Part Distribution: Explaining the Abundance of Wings", Journal of Archaeological Science 29, pp. 965-978.
Brain, Charles Kimberlin
1981. The Hunters or the Hunted? An Introduction to African Cave Taphonomy. Chicago, The University of Chicago Press.
Bunn, Henry T., Ellen M. Kroll y Lawrence E. Bartram
1991. "Bone Distribution on a Modern East African Landscape and its Archaeological Implications", en Cultural Beginnings. Approaches to Understanding Early Hominid Life-ways in the African Savanna, pp. 33-54. Bonn, Dr. Rudolf Habelt GMBA.
Corona Martínez, Eduardo
2001. "La relación hombre-ave en el siglo XVI. Crónicas y arqueozoología en el centro de México", Arqueología 25, pp. 109-120.
2002. Las aves en la historia natural novohispana. México, Instituto Nacional de Antropología e Historia. [ Links ]
Cruz, Isabel
1999. "Pingüinos de Cabo Vírgenes (Santa Cruz). Aspectos tafonómicos e implicaciones arqueológicas", en Actas delXIII Congreso Nacional de Arqueología Argentina, tomo 4, pp. 95-108. Córdoba, Universidad Nacional de Córdoba.
2000a. "Líneas tafonómicas y ecológicas para evaluar la explotación prehistórica de aves acuáticas en la zona cordillerana (Prov. de Santa Cruz)", en Desde el país de los gigantes. Perspectivas arqueológicas en Patagonia, tomo I, pp. 202-217. Río Gallegos, Universidad Nacional de la Patagonia Austral. [ Links ]
2000b. "Los restos de aves de los sitios arqueológicos del Parque Nacional Perito Moreno (Santa Cruz, Argentina)", en Anales del Instituto de la Patagonia, Serie Ciencias Sociales (Chile), 28, pp. 305-313. [ Links ]
2004. "Tafonomía de huesos de aves en Punta Medanosa (Depto. Puerto Deseado, Santa Cruz, Argentina)", en Contra viento y marea. Arqueología de la Patagonia, eds. María Teresa Civalero, Pablo Marcelo Fernández y Ana Gabriela Guraieb, pp. 455-468. Buenos Aires, Instituto Nacional de Antropología y Pensamiento Latinoamericano y Sociedad Argentina de Antropología. [ Links ]
2005. "La representación de partes esqueléticas de aves. Patrones naturales e interpretación arqueológica", Archaeofauna 14, pp. 69-81. [ Links ]
2006. "Los huesos de pingüinos (Spheniscidae) de los sitios de Cabo Blanco (Santa Cruz, Patagonia Argentina). Análisis tafonómico y perspectivas arqueológicas", Intersecciones en Antropología 7, pp. 15-26. [ Links ]
2007a. "La explotación de las aves en la zona cordillerana del sur de Patagonia (Santa Cruz, Argentina)", Werken 11, pp. 107-126. [ Links ]
2007b. "Avian Taphonomy: Observations at Two Magellanic Penguin (Spheniscus magellanicus) Breeding Colonies and Their Implications for the Fossil Record", Journal of Archaeological Science 34, pp. 1252-1261. [ Links ]
2007c. "The Recent Bones of the Río Gallegos Basin (Santa Cruz, Argentina) and Their Preservation Potential", en Taphonomy and Archaeozoology in Argentina, eds. María A. Gutiérrez, Laura Miotti, Gustavo Barrientos, Guillermo L. Mengoni Goñalons y Mónica Salemme, pp. 161-170. Oxford, BAR (British Archaeological Reports) International Series S1601. [ Links ]
2008. "Avian and Mammalian Bone Taphonomy in Southern Continental Patagonia. A Comparative Approach", Quaternary International 180, pp. 30-37. [ Links ]
2009a. "Tafonomía en escalas espaciales amplias: el registro óseo de las aves en el sur de Patagonia", en Temas de arqueología 2: estudios tafonómicos y zooarqueológícos, eds. Alejandro Acosta, Daniel Loponte y Leonardo Mucciolo, pp. 15-34. Buenos Aires, Instituto Nacional de Antropología y Pensamiento Latinoamericano. [ Links ]
2009b. "Tafonomía de huesos de cormoranes en la costa patagónica. Primeros resultados", en Arqueología de la Patagonia. Una mirada desde el último confín, pp. 729-741. Ushuaia (Argentina), Editorial Utopía. [ Links ]
Cruz, Isabel y Dolores C. Elkin
2003. "Structural Bone Density of the Lesser Rhea (Pterocnemia pennata) (Aves: Rheidae). Taphonomic and Archaeological Implications", Journal of Archaeological Science 30, pp. 37-44.
Cruz, Isabel y Pablo M. Fernández
2004. "Comprendiendo el pasado: avances en tafonomía de Rheidos", en Actas del Io Congreso Latinoamericano sobre Conservación y Cría Comercial de Ñandúes, CD-ROM. Buenos Aires, INTA-Rheacultura.com.ar-Secretaría de Agricultura, Ganadería, Pesca y Alimentación de la República Argentina.
Davis, Paul G. y Derek E. G. Briggs
1998. "The Impact of Decay and Disarticulation on the Preservation of Fossil Birds", Palaios 13, pp. 3-13.
deFrance, Susan
2005. "Late Pleistocene Marine Birds from Southern Peru: Distinguishing Human Capture from El Niño-induced Windfall", Journal of Archaeological Science 32, pp. 1131-1146.
Emslie, Steven D.
1995. "Age and Taphonomy of Abandoned Penguin Rookeries in the Antarctic Peninsula Region", Polar Record 31, pp. 409-418.
Ericsson, Per G.
1987. "Interpretations of Archaeological Bird Remains: A Taphonomic Approach", Journal of Archaeological Science 14, pp. 65-75.
Feduccia, Alan
1992 "Osteología de las aves", en Anatomía de los animales domésticos, eds. Samuel Sisson, J. B. Grossman y Robert Getti, tomo I, pp. 1960-1972, México, Editorial Salvat.
Fernández, Pablo Marcelo, Isabel Cruz y Dolores C. Elkin
2001 "Densidad mineral ósea de Pterocnemia pennata (Aves: Rheidae). Una herramienta para evaluar frecuencias anatómicas en sitios arqueológicos", Relaciones, Revista de la Sociedad Argentina de Antropología 26, pp. 243-260.
Fernández, Pablo M., Isabel Cruz y Ana Forlano
2010. "Sitio 37: una madriguera de carnívoro en el Norte de la Patagonia Andina (Cholila, Provincia de Chubut, Argentina)", en Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio, eds. María A. Gutiérrez, Mariana De Nigris, Pablo M. Fernández, Miguel Giardina, Adolfo F. Gil, Andrés Izeta, Gustavo Neme y Hugo D. Yacobaccio, pp. 409-417. Buenos Aires, Ediciones del Espinillo.
Fernández, Pablo M. e Isabel Cruz
2010. "The Fox, the Hare, and the Sheep. Bone Modifications in a Modern Carnivore Den at the Deciduous Andean Forest (Cholila, Argentina)", póster presentado en la Reunión Internacional del ICAZ (International Council of Archaeozoology). París (Francia).
Giardina, Miguel A.
2006. "Anatomía económica de Rheidae", Intersecciones en Antropología 7, pp. 263-276.
Gifford, Dianne P.
1981. "Taphonomy and Paleoecology: A Critical Review of Archaeology's Sister Disciplines", en Advances in Archaeological Method and Theory, ed. Michael B. Schiffer, 4, pp. 365-438. Nueva York, Academic Press.
Gilbert, B. Miles., Larry D. Martin y Howard G. Savage
1981. Avian Osteology Laramie, Modern Printing Co.
Higgins, Jude
1999. "Túnel: A Case Study of Avian Zooarchaeology and Taphonomy", Journal of Archaeological Science 26, pp. 1449-1447.
Kidwell, Susan y Anna K. Behrensmeyer
1988 "Overview: Ecological and Evolutionary Implications of Taphonomic Processes", en Palaeogeography, Palaeoclimatology and Palaeobiology 63, pp. 1-13.
Lefèvre, Christine
1989. "Les oiseaux", en Ethno-archéologie dans les archipels de Patagonie: les nomades marins de Punta Baja, ed. Dominique Legoupil, pp. 99-113. París, Editions Recherche sur les Civilisations, Mémoire No. 84.
1992. "Punta María 2. Los restos de aves", Palimpsesto, Revista de Arqueología 2, pp. 71-98. [ Links ]
1997. "Sea Bird Fowling in Southern Patagonia: A Contribution to Understanding the Nomadic Round of the Canoeros Indians", International Journal of Osteoarchaeology 7, pp. 260-270. [ Links ]
Livingston, Stephanie D.
1989. "The Taphonomic Interpretation of Avian Skeletal Part Frecuencies", Journal of Archaeological Science 16, pp. 537-547.
Lyman, Richard L.
1994. Vertebrate Taphonomy. Cambridge, Cambridge Universtiy Press.
Mameli, Laura
2002. "Bird Management in America's Extreme South during the 19th Century", en Acta zoologica cracoviensia 45, pp. 151-165.
Marean, Curtis W.
1995. "Of Taphonomy and Zooarchaeology", Evolutionary Anthropology 4, pp. 64-72.
Martín, Fabiana M.
1998. "Madrigueras, dormideros y letrinas: aproximación a la tafonomía de zorros", en Arqueología de la Patagonia Meridional (Proyecto Magallania), comp. Luis Alberto Borrero, pp. 73-96. Concepción del Uruguay, Ediciones Búsqueda de Ayllu.
Martínez, Ismael, Isabel Cartajena y Patricio López M.
2009. "Presence of Late-Pleistocene Rheidae in the Oasis of Calama (Second Region, Chile)", Current Research in the Pleistocene 27, pp. 33-35.
Morales Muñiz, Arturo
1993. "Ornithoarchaeology: The Various Aspects of the Classification of Bird Remains from Archaeological Sites", Archaeofauna 2, pp. 1-13.
Moreno, Eduardo, Alicia Castro, Karina Martinelli y Andrés Abello
1998. "El material faunístico de Cabo Blanco 1", Anales del Instituto de la Patagonia, 26, pp. 169-180.
Muñoz, A. Sebastián y Florencia Savanti
1998 "Observaciones tafonómicas sobre restos avifaunísticos de la costa noreste de Tierra del Fuego", en Actas y Memorias delXI Congreso Nacional de Arqueología Argentina (8° Parte), Revista del Museo de Historia Natural de San Rafael Tomo XX 1/2: 107-121.
Oliver, James S. y Russell W. Graham
1994. "A Catastrophic Kill of Ice-traped Coots: Time-Averaged Versus Scavenger-specific Disarticulation Patterns", Paleobiology 20, pp. 229-244.
Pickering, Travis, Kathy Schick y Nicholas Toth (eds.)
2007. Breathing Life into Fossils: Taphonomic Studies in Honor of C. K. (Bob) Brain. Gosport, Stone Age Institute Publication Series Number 2.
Savanti, Florencia
1994. Las aves en la dieta de los cazadores-recolectores terrestres de la costa fueguina. Temas de Arqueología. Buenos Aires, CONICET/PREP.
Sept, Jean M.
1994. "Bone Distribution in a Semi-arid Riverine Habitat in Eastern Zaire: Implications for the Interpretation of Faunal Assemblages at Early Archaeological Sites", Journal of Archaeological Science 21, pp. 217-235.
Tappen, Martha
1995. "Savanna Ecology and Natural Bone Deposition. Implications for Early Hominid Site Formation, Hunting, and Scavenging", Current Anthropology 36, pp. 223-260.
Tívoli, Angélica M.
2010. "Exploitation of Bird Resources among Prehistoric Sea-nomad Societies of the Beagle Channel Region, Southern South America", Before Farming 2, artículo 3.
Trapani, Joshua
1998. "Hydrodynamic Sorting of Avian Skeletal Remains", Journal of Archaeological Science 25, pp. 477-487.

