










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAntipoda. Revista de Antropología y Arqueología
Print version ISSN 1900-5407
Antipod. Rev. Antropol. Arqueol. no.13 Bogotá July/Dec. 2011
ARQUEOLOGÍA SUBACUÁTICA Y TAFONOMÍA: RECIENTES AVANCES EN EL ESTUDIO DE SITIOS FINIPLEISTOCÉNICOS SUMERGIDOS EN LA COSTA PACÍFICA DE CHILE CENTRAL
Isabel Cartajena F.* , Patricio López M.**, Diego Carabias A.***, Carla Morales **** y Gabriel Vargas*****
* Doctora en Arqueología Americana (Altamerikanistik, Freie Universität, Berlín). icartaje@uchíle.cl Departamento de Antropología, Universidad de Chile, Chíle.
** Doctor en Antropología. Programa de Doctorado en Antropología, Universidad Católica del Norte, IIAM, Chile. patriciolopezmend@yahoo.es Universidad Católica del Norte, IIAM, Chile.
*** Arqueólogo, Universidad de Chile, Chile. dcarabias@arkachile.cl ARKA Consultores S. A., Chile.
**** Doctora, Historia del Arte, Restauración y Conservación, Universidad Internacional S.E.K., Chile. cmorales@arkachile.cl ARKA Consultores S. A., Chile.
***** Doctor en Oceanografía y Paleoceanografía, Universidad de Bordeaux I, Francia. gvargas@ing.uchile.cl Departamento de Geología, Universidad de Chile, Chile.
RESUMEN
Se presentan los resultados del análisis tafonómico del sitio GNLQ1 (32° S - 71o O, Chile) descubierto a 13 m de profundidad bajo el mar. El sitio corresponde a un contexto primario conformado por una serie de restos de fauna continental extinta (familias Camelidae, Cervidae, Cricetidae, Artiodactyla, Equidae, Mylodontidae, Canidae, así como del Orden Xenarthra) recuperados en concentraciones dispuestas sobre una unidad estratigráfica asignada al Pleistoceno final. En este trabajo se abordan la determinación taxonómica y representación anatómica, además de las características superficiales del material, con el fin de conocer las condiciones paleoecológicas y delimitar los factores tafonómicos en contextos terrestres y subacuáticos.
PALABRAS CLAVE:
Arqueología subacuática, tafonomía, fauna extinta, Pleistoceno final, Chile central.
SUB-AQUATIC ARCHEOLOGY AND TAPHONOMY: RECENT ADVANCES IN THE STUDY OF SUBMERGED PHINIPLEISTOCENICS IN THE PACIFIC COAST OF CENTRAL CHILE
ABSTRACT
This paper presents the results of a taphonomic analysis of the site GNLQ1 (32 ° S - 71° W Chile) discovered at 13 m of depth below sea level. The site can be characterized as a primary context, conformed by diverse remains of extinct continental fauna (families Camelidae, Cervidae, Cricetidae, Artiodactyla, Equidae, Mylodontidae, Canidae and the Xenarthra Order) recovered in concentrations arranged on a stratigraphic unit also assigned to the Late Pleistocene. We deal with taxonomic determination, skeletal units' representations, and surface modifications of the material. We aim to understand paleo-ecological conditions and identify taphonomic factors in terrestrial and sub aquatic contexts.
KEY WORDS:
Underwater Archaeology, Taphonomy, Extinct Fauna, Late Pleistocene, Central Chile.
ARQUEOLOGIA SUBAQUÁTICA E TAFONOMIA: RECENTES AVANÇOS NO ESTUDO DE LUGARES FINIRLEISTOCÊ NICOS SUBMERSOS NA COSTA PACÍFICO DO CHILE CENTRAL
RESUMO
Apresentam-se os resultados da análise tafonô mica do lugar GNLQ1 (32° S - 71° O, Chile) descoberto a 13 cm de profundidade baixo o mar. O lugar corresponde a um contexto primário conformado por uma série de restos de fauna continental extinta (famílias Camelidae, Cervidae, Cricetidae, Artiodactyla, Equidae, Mylodontidae, Canidae e assim como da ordem Xenarthra) recuperados em concentrações dispostas sob uma unidade estratigráfica atribuída ao pleistoceno final. Neste trabalho abordam-se a determinação taxonô mica e representação anatô mica, além das características superficiais do material com o fim de conhecer as condições paleoecológicas e delimitar os fatores tafonô micos em contextos terrestres e subaquáticos.
PALAVRAS CHAVE:
Arqueologia subaquática, tafonomia, fauna extinta, pleistoceno final, Chile central.
Introducción
La arqueología subacuática es una disciplina que ha comenzado a ocupar su propio espacio dentro de los estudios arqueológicos en Chile, ya sea mediante el desarrollo de proyectos de investigación con objetivos particulares y claramente definidos, o bien, en el marco de estudios de impacto ambiental (Carabias, 2001, 2005). Las dificultades de trabajar en un medio diametralmente distinto al terrestre, unidas a procesos de formación disímiles a los tradicionalmente estudiados, han llevado a la arqueología subacuática a formular sus propios marcos teóricos y metodológicos y adaptar otras técnicas a partir de los referentes clásicos de la arqueología de campo. No obstante, la excavación en un contexto subacuático no difiere en demasía de la de sitios sobre tierra, siendo similares en el registro visual, las técnicas de excavación, la recolecta de muestras y el registro planimétrico, aunque cambian las condiciones y los tiempos de trabajo, debido a las limitantes del medio.
Los avances de la arqueología subacuática en las últimas décadas han posibilitado que esta disciplina haya ido consolidando metodologías propias en el estudio de restos de relevancia arqueológica que actualmente se encuentran sumergidos tanto en ambientes marinos como de agua dulce (Dean et al., 2000; Petriaggi y Davidde, 2007; Masters y Flemming, 1983). Si bien por tradición la arqueología subacuática y marítima ha enfatizado el estudio de contextos históricos, los trabajos en sitios prehistóricos son cada vez más frecuentes y se han enmarcado en problemáticas relacionadas con el uso de rutas costeras dentro de procesos de poblamiento inicial de una zona, el uso de antiguas embarcaciones, la aplicación de técnicas de pesca y recolección de recursos marinos y sus correlatos materiales (Blanton, 1996; Clausen et al., 1979; Dunbar et al., 1992; Faught, 1996, 2004a, 2004b; Faught y Donoghue, 1997; Flemming, 2004; Galili et al, 1993; Gusick y Faught, 2011). Así mismo, el estudio de contextos paleontológicos y arqueológicos prehistóricos ha permitido recolectar una gran cantidad de información geoarqueológica relacionada con el aumento del nivel del mar, o bien, la inundación de sectores de gran potencial para estudios paleoambientales y geomorfológicos (Dunbar et al, 1992; Faught, 1996; Faught y Donoghue, 1997; Faught, 2004a, 2004b; Flemming, 2004).
Dentro de la arqueología americana, las investigaciones subacuáticas han adquirido cierta relevancia en el estudio del poblamiento inicial del continente, debido al posible uso de la ruta costera (Dillehay, 2009; Goebel et al., 2008), cuyos sitios se encontrarían actualmente sumergidos, producto del avance marino durante el Pleistoceno final, y que suponen un poblamiento previo al denominado "Clovis First" (Bradley y Stanford, 2004; Dillehay, 2009; Faught, 1996, 2004a, 2004b; Goebel et al, 2008; Hoffecker et al, 1993). No obstante, la hipótesis de entrada por el Pacífico (Pacific Rim o Coastal-entry) es un modelo complejo que involucra diversas propuestas. Si bien la ruta por el Pacífico es lógicamente atractiva, la evidencia es escasa. Desde el Último Glacial Máximo, el nivel del mar ha subido hasta 120 m, inundando paisajes costeros y estuarios ribereños donde se podrían haber localizado los sitios. A excepción de Josen-hans et al. (1997), que han logrado localizar sitios en la costa de Queen Charlotte Islands, se han realizado escasos esfuerzos de búsqueda a lo largo de las líneas costeras sumergidas (Bonnichsen et al., 2005: 321). Si bien la evidencia arqueológica señala la presencia de adaptaciones costeras, también cabe esperar la presencia de cazadores terrestres, no en el sentido planteado por Bon-nichsen et al. (2005) en cuanto a la reubicación de asentamientos y un acercamiento a los recursos costeros ante un escenario cambiante, sino en el marco de la utilización de quebradas y estuarios por parte de grupos interiores, cuyas evidencias quedaron cubiertas posteriormente por el mar.
El reconocimiento de yacimientos en este contexto no involucra únicamente la recolección de datos paleoambientales, geomorfológicos o evidencias artefactuales y/o ecofactuales, sino que además, como todo sitio arqueológico temprano, requiere una contrastación rigurosa de la antigüedad real de las ocupaciones humanas, así como de la integridad de las asociaciones esperadas para estos eventos ocupacionales (Stewart, 1999; Waters, 1992). Al respecto, uno de los problemas recurrentes a lo largo de las investigaciones de sitios del Pleistoceno final se centra en una evidencia poco diagnóstica para discriminar eventos naturales y/o culturales en la generación de un yacimiento determinado, de manera específica, entre las evidencias osteofaunísticas de mamíferos actualmente extintos producidas por agentes naturales versus la acción humana. Este problema ha sido abordado con profusión, y la tafonomía constituye un referente metodológico y teórico clave para el entendimiento e interpretación de sitios donde los agentes naturales y culturales se mezclan, o son difíciles de separar (Bonnichsen, 1982; Borrero, 2003; Frison, 1982; Hill y Behrensmeyer, 1985).
Los proyectos orientados al estudio de sitios tempranos se caracterizan por el trabajo conjunto de numerosos especialistas, con el fin de entender y comprender los conjuntos recuperados. Disciplinas como la geología, geoar-queología, botánica, palinología, botánica y zooarqueología se congregan comúnmente para el estudio de dichos yacimientos. Mediante este trabajo conjunto, no sólo se busca la colaboración y compartir experiencias sino abrir nuevas perspectivas de análisis.
Reflexiones en torno a una tafonomía en contextos subacuáticos
Dentro de los estudios tafonómicos, los depósitos de mamíferos de talla mediana a grande han sido abordados en detalle, tanto en el marco de trabajos experimentales (Binford, 1981; Blumenschine, 1995), etnoarqueológicos (Bunn et al., 1988; Bunn, 1993; O'Connell et al., 1990), como mediante observaciones actualísticas en ensambles faunísticos modernos (Beherensmeyer, 1978; Borrero y Martin, 1996; De Ruiter y Berger, 2000; Domínguez-Rodrigo, 1999; Haynes, 1983a, 1983b). No obstante, los agentes y procesos tafonómicos estudiados se enmarcan en contextos terrestres, y escasamente han sido pensados para sitios sumergidos (Stewart, 1999; Stright, 1995; Waters, 1992). Dentro de estas reflexiones destacan las de Dunbar y colaboradores (1989), que proponen, a partir del caso del sitio de Aucilla River (Florida, EE. UU.), al menos cinco tipos de modificaciones que deben ser consideradas en contextos finipleistocénicos sumergidos con restos de fauna, a saber: 1) modificaciones culturales en huesos frescos sincrónicas a la muerte del animal (por ejemplo, huellas de corte), 2) modificaciones naturales previas a la depositación de los huesos (por ejemplo, meteorización, marcas de carnívoros), 3) modificaciones naturales posdeposita-cionales (mineralización, degradación por acción del suelo), 4) modificaciones naturales dentro de la interfaz sedimento-agua (por ejemplo, oxidación, abrasión) y 5) alteraciones posexcavación producto del cambio de ambiente de los huesos. Esta alteración, sin embargo, es controlable a partir de un debido trabajo de conservación y restauración de las osamentas (Bouzas y Laborde, 2003).
A estas alteraciones agregamos aquellas que se producen en sitios con alta frecuencia de tránsito de barcos de distintos calado como muelles y puertos, además de zonas aledañas a pueblos y ciudades costeras. En el primer caso, los muelles y puertos, como la bahía de Quintero, constantemente movilizan y arrastran anclas, cadenas, o "muertos" que corresponden a bloques de concreto que alteran subsuperficialmente el fondo marino. En el caso de las poblaciones humanas asentadas en las cercanías de la costa, éstas producen alteraciones, debido a la incorporación de basura y tubos de desagüe.
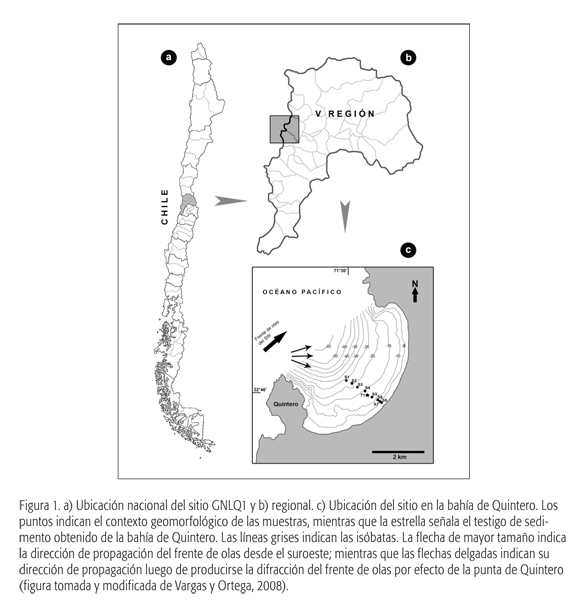
En el caso del sitio a estudiar, GNLQ1, ubicado en localidad de Quintero (Latitud: 32° 46' 59" S; Longitud: 71° 31' 59" O), en el litoral de Chile central, se encuentra aproximadamente a 13 m de profundidad bajo el mar y presenta características únicas en la costa del Pacífico sur. El sitio GNLQ1 corresponde a un contexto primario conformado por una serie de restos de fauna continental extinta del Pleistoceno final, depositados en sedimentos terrígenos actualmente sumergidos. Dentro de un área de 1 m2 de excavación, se registraron diversos restos del esqueleto apendicular y axial de especies extintas pertenecientes a las familias Camelidae, Cervidae, Cricetidae, Artiodactyla, Equidae, Mylodontidae, Canidae y restos del Orden Xenarthra, que fueron recuperados en concentraciones discretas dispuestas sobre una unidad estra-tigráfica caracterizada por una grava arcillosa en una matriz de arena fina a limo y arcillas con presencia de lentes de carbón. Los antecedentes faunísticos, geomorfológicos y sedimentológicos sugieren que el contexto durante el Pleistoceno final correspondía a un ambiente tipo estuario-lagunar, con un período de somerización-disecación, a uno de tipo de anteplaya superior, inundado por el mar.
Las características del emplazamiento permiten, de acuerdo con Bishop (1980), adscribirlo a aquellos paleoambientes fosilíferos en regímenes continentales, que permiten la fosilización y conservación. Entre ellos se encuentran las aguas poco profundas de estuario, los deltas y depósitos lagunares asociados a las transgresiones marinas (Bishop, 1980: 23). Además, en un eje temporal, el contexto terrestre representa una escala temporal corta, en la cual es posible observar cambios rápidos en el ambiente, que ocurren y se perciben en el nivel sedimentario. Por otra parte, en su paso a un contexto subacuático, es posible adscribirlo a un ambiente instantáneo ("Instant environment" sensu Bishop, 1980: 22), definido como eventos a corto plazo, que permiten un conocimiento acabado de un evento catastrófico de diversa magnitud. En este medio acuoso, por su parte, el conjunto no presenta las particularidades de restos transportados o expuestos al movimiento de las aguas (Hanson, 1980).
A través del presente trabajo se busca caracterizar el conjunto faunístico recuperado del sitio GNLQ1, tanto al nivel de la representación de unidades anatómicas como de la diversidad taxonómica, poniendo especial hincapié en el análisis taxonómico, para lo cual se consideraron las características de las superficies, integridad y articulación de unidades. Si bien este análisis es preliminar, mediante los resultados obtenidos se busca conocer las condiciones paleoecológicas y delimitar los factores tafonómicos en contextos terrestres y subacuáticos (Behrensmeyer y Hill, 1980). Así mismo, a partir de este análisis se busca generar expectativas para el desarrollo de los análisis zooarqueológicos y tafonómicos en estrecha relación con programas de investigación que intenten formalizar la búsqueda de yacimientos arqueológicos prehispánicos sumergidos y su potencial para comprender y caracterizar de mejor manera procesos de poblamiento humano hacia el Pleistoceno final.
El sitio GNLQ1
Como se mencionó anteriormente, el sitio GNLQ1 se ubica en la bahía de Quintero, emplazada al norte de Valparaíso, V Región de Chile (ver las figuras 1a, 1b y 1c). Ésta corresponde a una bahía somera, con una profundidad máxima de 55 a 60 m, abierta hacia el noroeste. En ella descargan algunas quebradas con escorrentía de permanente a perenne, que generan lagunas y/o humedales entrampados por barras de arena asociadas a la acción de la hidrodinámica litoral (Vargas y Ortega, 2008). El perfil batimétrico muestra una mayor pendiente en los primeros ca. 250 m desde la línea de costa y una menor pendiente hasta los 1.600 m alejados de ésta. El sitio fue descubierto durante las labores de prospección y excavación arqueológica, en el marco de un proyecto relacionado con una terminal de gas.
En este contexto se realizó una serie de sondeos subsuperficiales, en puntos distribuidos a lo largo de una transecta trazada dentro de un área de 40 x 25 m (Carabias, 2008). A partir de estas evidencias, se excavó una unidad de 1 x 1 m (Unidad K8) delimitada sobre una concentración de huesos que asomaban del fondo marino (erosionado por las actividades realizadas en el muelle cercano) y que presentaban potencial estratigráfico, de acuerdo con los sondeos realizados previamente. En el caso de la excavación del sitio GNLQ1, se procedió mediante niveles artificiales de 10 cm, registrando todo tipo de evidencias en planta, extrayendo el sedimento mediante dragado para ser harneado.

A partir del análisis del testigo se determinaron tres unidades estratigráficas con características sedimentológicas distintas, que por orden de antigüedad son: Unidad 1, Unidad 2 y Unidad 3. La Unidad 1 está constituida por arena fina de buena selección, con un sedimento de color café. La Unidad 2 está constituida por grava arcillosa clasto-soportada de aglomerados de arena fina y arcilla, y eventualmente calcedonia, de alta dureza, de color naranja y gris, algunas con pátinas de carbón, en una matriz de arena fina a limo y arcillas. En la parte superior e inferior de la unidad se intercalan lentes milimétricos de carbón. La Unidad 3 está constituida por arena fina arcillosa de color café anaranjado (Vargas y Ortega, 2008).
Las evidencias óseas fueron encontradas en la Unidad 2, distribuidas horizontalmente sobre una extensa área, directamente asociadas a lentes de carbón presentes en los primeros 5 cm de la Unidad 2 (Vargas y Ortega, 2008; ver las figuras 2a y 2b). Las muestras, luego de extraídas, fueron desalinizadas mediante lavados progresivos en agua desmineralizada. Al respecto, durante la excavación de los sondeos del sitio GNLQ1, los materiales recuperados fueron extraídos manteniendo gran parte del sedimento o matriz que los rodeaba. Este levantamiento "en bloque" permitió rigidez y soporte al hueso y sus fragmentos, y minimizó la pérdida de información y deterioro físico que pudiera ocasionar la extracción. Considerando la fragilidad de algunos de los restos, se puso especial cuidado en no afectarlos mecánicamente durante la intervención arqueológica. El primer daño que pueden sufrir los materiales orgánicos, cualquiera que éstos sean, es la pérdida de humedad. Para evitarla, se dispuso de recipientes individuales con agua salada para ir introduciendo los objetos a medida que son extraídos del fondo, impidiendo que se produzca la deshidratación de éstos, y así no romper el equilibrio que han logrado estando en el fondo marino. Los materiales óseos, así como la mayor parte de los materiales recuperados de sitios arqueológicos en tierra o bajo el agua, son vulnerables también a los cambios bruscos de temperatura. Para controlar ese tipo de daño los huesos se fueron almacenando, según se iban levantando, en una zona protegida de los rayos solares, en tanques de almacenamiento realizados especialmente para este proyecto.
Una vez despejado, el material óseo se depositó en contenedores individuales con agua salada proveniente del mismo sitio. Allí se dio inicio al segundo proceso, quizás el más importante, del que dependen el éxito de la estabilización de los materiales y su posterior conservación. Este proceso consistió en bajar los niveles de sal que contienen, primero cambiando de manera gradual el agua que los contenía por agua libre de minerales. Este reemplazo se realizó periódicamente, con cambios del porcentaje de agua salada versus agua destilada una vez cada diez días, hasta que el 100% fuera agua dulce.
Después de 50 días comenzó el tratamiento de rinsing de los huesos. Éste consistió en un lavado delicado con agua corriente utilizando una escobilla suave para acelerar la limpieza, teniendo cuidado de no rayar la superficie de la pieza. Mientras se realizaban estos lavados, los materiales fueron depositados en contenedores con agua desmineralizada renovada periódicamente en un lugar oscuro y fresco para evitar el desarrollo de microorganismos, como hongos o bacterias. Al igual que en el proceso anterior, se buscó liberar al material óseo de la sal que tenía en su interior; en este caso en particular, los lavados se realizaron una vez por semana durante cuatro meses, hasta que los controles de acidez fueran bajos y pudiera iniciarse el último de los tratamientos, el secado. Estos controles se realizaron midiendo el pH durante cada cambio de agua, y correspondieron a uno de los procesos más importantes durante el tratamiento de conservación (Carabias, 2008).
Ahora bien: para determinar la antigüedad del sitio fueron enviadas dos muestras de hueso de paleocamélidos para su datación por C14 (AMS), pero no presentaban colágeno suficiente. De acuerdo con el ambiente de sedimentación de las Unidades 3 y 2, el sitio GNLQ1 habría sido un estuario (Vargas y Ortega, 2008), y teniendo en cuenta la variación global del nivel del mar, junto a un despreciable alzamiento tectónico para la zona en los últimos miles de años, se podría tentativamente sugerir una edad superior a los 7.000 años AP, para la formación de la Unidad 2, puesto que el nivel del mar se encontraba alrededor de 10-20 m más bajo que su posición actual, entre los 7.000-9.000 años AP, respectivamente (Lambeck et al, 2002). Lo anterior ha quedado ratificado a partir de una muestra de sedimentos de la parte inferior de la matriz en donde se encontraban los restos (Unidad 2) bajo los restos fósiles, la cual aportó una fecha por C14 de 13.640±40 años AP sin calibrar (UGAMS#9194).
Resultados Características del conjunto óseo
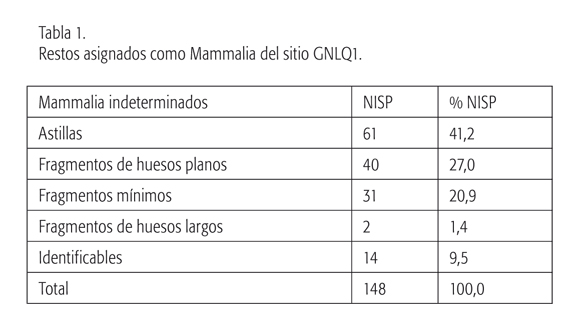
Se analizó un total de 224 especímenes óseos (ver la tabla 1), que actualmente se encuentran depositados en Departamento de Antropología de la Universidad de Chile. Los restos provienen mayoritariamente de la cuadrícula K8-2 (Nivel 1, Concentraciones 1, 3 y 6), y, como se mencionó anteriormente, previo a cualquier análisis fueron sometidos a largos procesos de estabilización y conservación producto del cambio brusco de ambiente (Carabias, 2008).
El conjunto óseo se encuentra en muy buenas condiciones de conservación. No obstante, algunos especímenes evidencian estados iniciales de meteo-rización como agrietamientos longitudinales en fragmentos de diáfisis, lo que sugiere una exposición a condiciones subáreas previa a su enterramiento final. Esta meteorización podría haber favorecido la fragmentación del conjunto, en especial astillas (27% de la muestra total) y fragmentos de huesos planos (17% de la muestra total). Pese a esto, un porcentaje no menor pudo ser remontado, pese a la erosión de los bordes de los huesos producto de la constante acción del agua y sedimentos abrasivos propios del ambiente de depositación del sitio.
Determinación taxonómica
Para la determinación taxonómica de los restos se utilizaron colecciones de referencia albergadas en el Departamento de Antropología de la Universidad de Chile correspondientes a équidos, camélidos y cérvidos actuales. Así mismo, se utilizó la colección paleontológica del sitio Kamac Mayu (II Región, Chile) depositada en el Laboratorio de Estudios de Arqueología y Patrimonio (LEAP) de la Universidad Internacional SEK-Chile. Debido a las cercanías y similitud faunística con el sitio GNLQ1, se utilizaron las colecciones del sitio Quereo (IV Región), que están actualmente depositadas en el Museo Arqueológico de La Serena.
Para los restos de Canidae fueron utilizadas colecciones particulares de Pseudalopex griseus y Lycalopex culpaeus recolectadas en las cercanías al área de estudio, y que tenían por objeto caracterizar las diferencias de tamaño y morfología. Finalmente, en el caso de los restos de xenartros, el material paleontológico disponible en Chile para fines comparativos es escaso; sin embargo, existe en el Museo Nacional de Historia Natural de Santiago material bastante completo de Megatherium medinae y Glossotherium, que fue utilizado para la identificación anatómica de los restos. Debido a la relevancia del sitio, la fase de asignación taxonómica del material de GNLQ1 aún se encuentra en proceso, por lo que en el presente trabajo se discutirán y presentarán datos generales relacionados con este tema.
Clase Mammalia
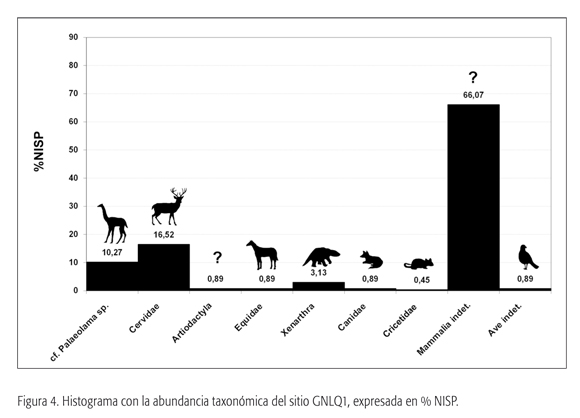
Esta categoría taxonómica es la que presenta la mayor representación de restos dentro del conjunto (ver la figura 4). La mayor parte de los especímenes corresponden a astillas, fragmentos de huesos planos (muchos de ellos restos de costillas), fragmentos mínimos y algunas unidades anatómicas identificables como fragmentos de vértebras y epífisis humeral y femoral (tabla 1). Los restos corresponderían a grandes mamíferos, encontrándose tanto fragmentos pertenecientes a individuos adultos como juveniles (no fusionados).
Clase aves
Estos restos fueron registrados en una cuadrícula diferente (K7), en la cual la presencia de huesos era mínima. Se recuperaron un fémur y una tibia de ave correspondientes a un individuo joven, lo que dificulta su asignación taxonómica. Sin embargo, la morfología y tamaño es compatible con especímenes de la familia Phasanidae.
Orden Artiodactyla
Se recobró un premolar deciduo de tipo selenodonte propio de un artiodáctilo. De igual forma, se registró un fragmento de cráneo (meato auditivo). Ambos restos, ante la ausencia de otros rasgos diagnósticos, se asignaron a esta categoría taxonómica.
Familia Camelidae
cf. Palaeolama sp.
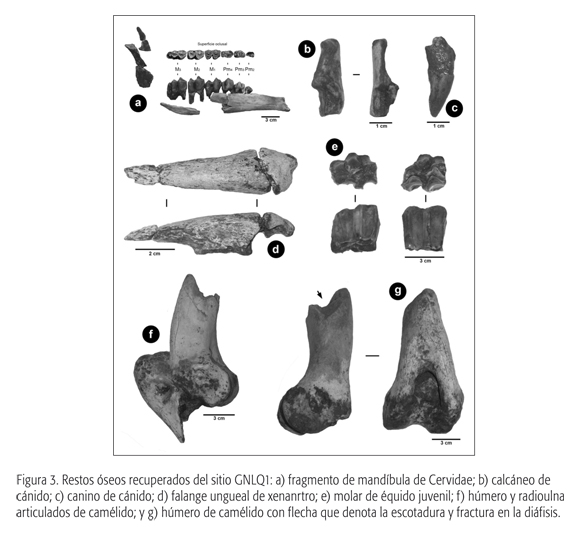
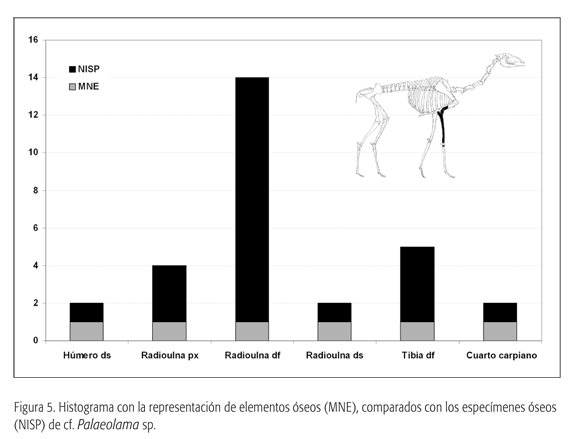
El total de restos recuperados es de 23 especímenes. Éstos corresponden principalmente a la extremidad anterior superior (húmero distal y radioulna), un cuarto carpiano y fragmentos de tibia (ver las figuras 3f, 3g y 5). La conservación de la epífisis permitió la obtención de medidas, que, comparadas con especímenes de guanaco (Lama guanicoe) y Palaeolama sp. provenientes de depósitos del Pleistoceno final de la zona aledaña de Los Vilos (31° S), denotan un camélido de gran tamaño, comparable con los géneros Palaeolama y Hemiauchenia.
En el sitio GNLQ1 no fueron recuperados materiales que permitieran una asignación certera al género Palaeolama. Los restos recuperados (húmero distal y radioulna) son similares en tamaño y morfología al material recobrado del sitio de Quereo (López et al., 2004). De acuerdo con esto, se ha clasificado el material de Quintero como cf. Palaeolama, en espera de un registro más completo.
Familia Cervidae
El material analizado corresponde a fragmentos de la mandíbula izquierda con la serie dental completa, es decir, Pm2, Pm3, Pm4, M1, M2 y M3, además del I3, Pm2, Pm3, Pm4 derechos de un mismo individuo (ver la figura 3a). En comparación con Hippocamelus antisensis (taruca del norte o huemul), los restos de Quintero son de mayor tamaño, acercándose a géneros como Antifer y Blastocerus. Ahora bien, dados los escasos antecedentes sobre la sistemática de cérvidos extintos en territorio chileno (Casamiquela, 1984, 1999; Labarca y Alcaraz, 2011), se ha optado por asignar el material en estudio como Cervidae gen. et sp. indet.
Familia Equidae
El material estudiado corresponde a un P2 y un P3, ambos derechos, de un animal juvenil de aproximadamente dos años (figura 3e). La superficie oclusal, en el caso del P3, prácticamente no presenta desgaste y comienza a denotar el protocono, aunque con un pliegue caballuno casi inexistente. Si bien la forma del protocono tiende a ser ovalada, el escaso desarrollo de diente impide asignarlo con certeza al género Hippidion. Debido a esto, han sido asignadas ambas piezas como Equidae gen. et sp. indet.
Orden Xenarthra
El registro fósil de xenartros recobrados del sitio GNLQ1 corresponde a un osteodermo, la secuencia completa de los molariformes de la mandíbula derecha aparentemente de un mismo individuo y una falange ungueal (figura 3d).
En el caso de los molariformes, las superficies oclusales por forma son afines a especies de la Subfamilia Scelidotheriinae. Por su parte, la ausencia de rasgos diagnósticos de la falange ungueal impide por el momento asignarlo a una Familia específica. Por su parte, los osteodermos están presentes tanto en especies de la Familia Mylodontidae como en algunos ejemplares de Glyptodon (Rinderknecht, 2000). Este material se encuentra en proceso de estudio, y, de acuerdo con resultados preliminares, la totalidad del registro ha sido clasificada como Xenarthra gen. et sp. indet., en espera de mayores comparaciones.
Familia Canidae
El material de Quintero corresponde a un calcáneo y a un canino cuya morfología y cuyo tamaño son similares al actual Lycalopex culpaeus. No obstante, debido a lo escaso del material, se ha asignado el registro del sitio GNLQ1 como Canidae gen. et sp. indet. (figuras 3b y 3c).
Familia Cricetidae
Se recuperó un incisivo de un roedor de un Cricetidae de baja diagnosis taxonómica.
Tafonomía
Alteraciones posdepositacionales del sitio
Tal como se mencionó en párrafos anteriores, el sitio GNLQ1 se encuentra emplazado en una bahía somera que ha funcionado a lo largo de su historia como puerto natural para embarcaciones de distinto calado, y en la actualidad es un muelle artificial usado como terminal para el ingreso de gas licuado al país. A causa de ello, recalan en las cercanías del sitio embarcaciones de gran tamaño, lo que ha motivado a definir un área de exclusión alrededor del sitio para su debida protección (Carabias, 2008). Sin embargo, en el momento de descubrirse el yacimiento GNLQ1 se observó una serie de alteraciones menores, que afectaron mediante erosión los sedimentos superiores que sellaban el sitio y, por tanto, han acelerado la exposición superficial de algunas concentraciones de restos fósiles. Esta erosión se ha producido por la acción mecánica sobre el fondo marino de trabajos submarinos desarrollados en el marco de la construcción del muelle producto del traslado de "muertos', bloques de cemento y cadenas que son llevados en remolques mediante embarcaciones en superficies (Carabias, 2008). Otro proceso de alteración que tiene una larga data en la bahía es el fondeo de anclas que constantemente remueven el fondo del muelle. Si bien estas alteraciones son continuas y han afectado mediante la erosión ciertos sectores del sitio, no han movilizado mayormente restos óseos, por cuanto se ha observado en dichas concentraciones cierta integridad anatómica, debido a que unidades de un mismo individuo que se articulan entre sí se disponen espacialmente en un mismo espacio acotado.
Otras alteraciones posdepositacionales corresponden a la incorporación de basura actual sobre la capa superior del sitio. Estas incorporaciones han sido discriminadas sin problemas, debido a que corresponden a desechos históricos absolutamente diacrónicos con el depósito pleistocénico.
Alteraciones tafonómicas en el registro óseo
Abrasión
Entre las alteraciones tafonómicas comunes para la totalidad de la muestra, o buena parte de ésta, se observa un leve grado de abrasión de la superficie de los huesos. Esta abrasión es homogénea y se manifiesta como un pulimento parejo tanto en los bordes como en las superficies planas de las osamentas. Esta alteración es esperable, debido a las condiciones de enterramiento de los huesos en un ambiente acuoso y de sedimentos abrasivos, aunque no se afectaron de forma muy agresiva, puesto que es posible distinguir rasgos como marcas producidas por distintos agentes biológicos (carnívoros y roedores), y aún pueden remontarse sin problemas astillas pertenecientes a un mismo elemento óseo, tal como se señaló anteriormente.
Alteraciones químicas
Una segunda alteración común a todos los restos es la presencia de manchas de oxidación producto de la interacción de los huesos con una interfaz de sedimentos de grava-arcillosa y agua (ver alteración química en la tabla 2). Este tipo de alteración es frecuente en ambientes similares a los del sitio GNLQ1 (Dunbar et al., 1989; Noakes et al., 2009), por lo que se la debe considerar a futuro como una expectativa dentro de las alteraciones tafonómicas. Por su parte, la presencia de sal en el agua marina y, por ende, en los huesos fue controlada mediante lavados continuos con agua desmineralizada, como se mencionó anteriormente.
La meteorización afectó al 87,5% de la muestra (ver la tabla 2). Esta alteración es leve y se concentra en los estadios 1 a 2, según la escala de Behrensmeyer (1978). Así mismo, indica que los restos quedaron expuestos a condiciones subaéreas, previo a su enterramiento y posterior inundación. Cabe destacar que los restos de cf. Palaeolama presentan un estadio de meteorización más avanzado que el resto de la muestra (Estadio 2), que no sólo indica tiempos más prolongados de exposición, sino que también sugiere que en el sitio GNLQ1 se formó a partir de distintos eventos de enterramiento.

Roedores
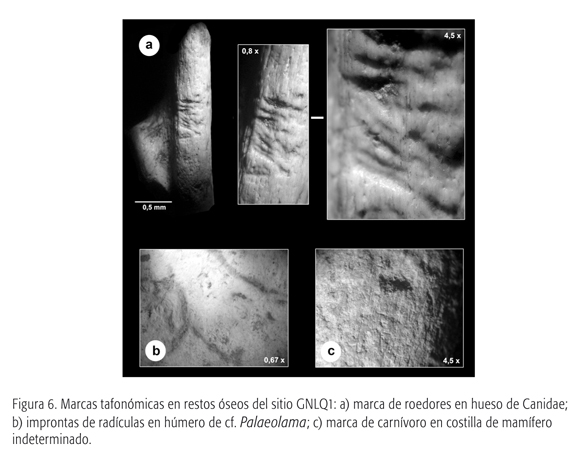
Las únicas marcas producidas por roedores se concentran en el calcáneo de cánido (ver la figura 6a). Esta evidencia presenta características propias del paso de incisivos de roedores dentro de una zona acotada en la diáfisis del calcáneo, a modo de líneas delgadas, cortas y paralelas.
Radículas
La mayor parte del registro óseo presenta en su superficie evidencias de improntas de raíces (ver la tabla 2 y la figura 6b). La presencia de estas improntas en los restos no sólo indica el crecimiento de una cubierta vegetacional sobre los depósitos fósiles previa a la transgresión marina, sino que además es un buen indicador que sugiere cierta estabilidad del sitio. Al respecto, las improntas se distribuyen marcadamente sobre la superficie superior de las osamentas y decrecen en la superficie inferior, e indican que los huesos no se movilizaron dentro del estrato portador, previa a la inundación del sitio.
Carnívoros
La acción de carnívoros se observó en el 5,8% de la muestra total. Pese a lo bajo de la incidencia de carnívoros en la alteración del conjunto óseo, estos agentes son significativos, ya que no sólo aportan información referente a los procesos de formación del sitio sino que además sirven para una reconstrucción paleoecológica del ensamble faunístico.
Ahora bien, de la totalidad del conjunto, los especímenes de cf. Palaeo-lama sp. presentan la mayor cantidad de marcas producidas por carnívoros. Por un lado, la epífisis distal de húmero presenta dos perforaciones circulares, que podrían ser atribuidas a la acción de un carnívoro de gran tamaño, y que muestran características similares a las observadas en restos de mamíferos extintos de diversos sitios registrados en la costa de la zona aledaña de Los Vilos (López, 2007). En la diáfisis de este hueso se observa una fractura transversal, que guardaría relación con la acción del carnívoro, puesto que en la superficie dorsal de la diáfisis y en el borde de la fractura se observan dos escotaduras cuyo agente podría ser el mismo carnívoro de gran tamaño, ya que presentan una distancia (ca. 35 mm) similar a las perforaciones registradas en la epífisis.
Tal como se señaló en el párrafo anterior, este tipo de marcas presentes en especies de similar tamaño y peso que Palaeolama en distintos sitios paleontológicos y arqueológicos de la zona de Los Vilos fueron atribuidas por López (2007) a un carnívoro de gran tamaño, específicamente a Panthera onca mesembrina. En el caso del sitio GNLQ1, no se ha identificado con claridad el agente específico de las alteraciones producidas en los restos de cf. Palaeolama, aunque parecen indicar un predador cuya talla supera a la de los cánidos, y se relacionan más bien, por morfología y tamaño, con las de un gran carnívoro (Borrero y Martin, 1996). Estas marcas fueron observadas también en la epífisis distal del radio de cf. Palaeolama, que por su tamaño y proximidad espacial parece corresponder al mismo individuo al que pertenece el resto del material asignado a este camélido extinto.
Dentro de los daños producidos por carnívoros se identificaron en fragmentos óseos y cuerpos vertebrales no identificados marcas de menor tamaño comparadas con las descritas para los restos de cf. Palaeolama sp., y que dan cuenta de la acción de pequeños carnívoros. Éstas son compatibles con la morfología y características de las modificaciones de cánidos, cuyos restos están presentes en el sitio GNLQ1 (ver la figura 6c).
Evidencias culturales
En los restos de cf. Palaeolama sp., el húmero distal presenta una fractura sobre hueso fresco. No obstante, esta fractura se asocia con perforaciones producidas por la acción de un gran carnívoro, por lo que es probable que este agente haya sido el causante de los daños observados en el húmero (Borrero y Martin, 1996; Haynes, 1983a). Si bien, este tipo de fracturas fue considerado propio de las actividades humanas, diversos análisis determinaron el grado de ambigüedad de este rasgo, ya que también aparecen en contextos formados por agentes naturales (Haynes, 1983a, 1983b; Myers et al, 1980). Cabe destacar, no obstante, el notable parecido de este espécimen con un húmero distal proveniente del cercano sitio de Quereo, donde se observa una fractura muy similar y que ha sido interpretada como cultural (López et al., 2004). Pese a esto, las diferencias observadas en el caso de la muestra de Quereo tienen relación con la morfología y los rasgos asociados al borde de fractura, puesto que se observaron escotaduras y negativos de impacto producidos por el golpe directo sobre la diáfisis del fósil de Quereo, que en el húmero del sitio GNLQ1 no se observan con claridad y además se asocian a las marcas de carnívoros descritas anteriormente.
La totalidad de la evidencia ósea no permite atestiguar de manera clara la acción antrópica en el sitio GNLQ1. No obstante, el análisis sedimentológico de la Unidad 2, es decir, la capa portadora del registro fósil, evidenció lentes milimétricos de carbón cuyo origen podría deberse eventualmente a combustiones locales producidas en una ambiente de sedimentación de tipo somero a estuarial-lagunar (Vargas y Ortega, 2008).
Ahora bien: en las superficies óseas se observaron manchas producidas por la oxidación propia de la interfaz del sedimento con el agua, aunque en algunos ejemplares estas manchas parecían mezclarse en algunos huesos con pátinas carbonosas difíciles de separar macroscópicamente. Análisis mediante Microscopio Electrónico de Barrido (MEB) y Espectroscopía de Energía Dispersiva (EDS) indican que la totalidad de estas manchas son producto de alteraciones diagenéticas, debido a la presencia de óxidos de hierro.
Discusión y conclusiones
La información geomorfológica, sedimentaria y paleontológica del sitio GNLQ1 sugiere la presencia de un contexto primario con escasas alteraciones posteriores al depósito de los restos óseos. La unidad estratigráfica portadora de los restos fósiles, denominada Unidad 2, sugiere un ambiente estuarial-lagunar en proceso de desecación, el cual dio paso a un ambiente de anteplaya superior inundada por el mar. En relación con las dataciones del evento de enterramiento del conjunto fósil, si se consideran la variación global del nivel del mar y el alzamiento tectónico de la zona en los últimos miles de años, se sugiere una edad superior a los 7.000 años AP para la formación de la Unidad 2, puesto que el nivel del mar se encontraba alrededor de 10-20 m más bajo que su posición actual, entre los 7.000-9.000 años AP, respectivamente (Vargas y Ortega, 2008). Esta observación es consistente con el fechado de C14 obtenido de una muestra de la matriz sedimentaria sobre la cual se encuentra depositado el conjunto óseo.
Del análisis tafonómico se desprenden diversos agentes que alteraron el registro previo, durante y posterior a su ingreso a la litósfera. Al respecto, se identificó la acción de al menos dos tipos de carnívoros sobre los restos, denotados por el tamaño y distancia de las punturas presentes. Uno de los agentes modificadores correspondería a un carnívoro de gran tamaño, como lo indican las marcas dejadas en huesos largos de cf. Palaeolama sp. Por otra parte, se observaron punturas más pequeñas compatibles con una arcada similar a la dejada por cánidos. En menor medida, la acción de roedores se manifiesta claramente en las marcas observables en un calcáneo perteneciente a un carnívoro. Por su parte, la presencia de improntas de raicillas indica el crecimiento de una cubierta vegetal, lo que denota que los restos, una vez expuestos y antes de la transgresión marina, habrían sido cubiertos por sedimentos, lo que favoreció la preservación e integridad del registro.
Dentro de las alteraciones producidas después del ingreso de los huesos en los sedimentos, la más elocuente corresponde a las manchas oxidación producto de la interacción de los huesos con una interfaz de sedimentos de grava-arcillosa y agua, que pueden ser consideradas como propias de ambientes depositacionales similares a los del sitio GNLQ1.
Por último, si bien no se identificaron restos culturales artefactuales ni en los sondeos ni en superficie, ni claros rastros de consumo humano de la fauna estudiada, no debe descartarse la acción del hombre en parte de la conformación del sitio. En relación con esta hipótesis, las investigaciones desarrolladas en la costa del norte semiárido chileno señalan una alta densidad de sitios con mamíferos extintos distribuidos en ambientes lagunares y ecosistemas estua-rinos que se formaron a partir de eventos de mortandad natural de especies actualmente extintas, ya sea mediante la incorporación de carcasas y huesos dejados por carroñeros y carnívoros, o bien, por la acción predadora de las primeras poblaciones humanas asentadas en la zona (Jackson et al., 2005; López et al., 2004; López, 2007). El resultado de la intervención de estos tres agentes se encuentra muchas veces conjuntamente en un mismo sitio, como en el caso del sitio de Quereo y en otros sitios subsuperficiales del semiárido, observándose incluso posibles evidencias de carroñeo humano sobre restos dejados por grandes carnívoros (López, 2007; Núñez et al., 1994). A esto se suma que la evidencia artefactual es mínima y se complementa o prioriza el uso de huesos como artefactos expeditivos para el procesamiento de las carcasas (Jackson et al., 2007; López, 2007; Núñez et al., 1994).
Estos antecedentes proponen un escenario que debe evaluarse en futuras investigaciones y sugieren que las evidencias del sitio GNLQ1 no son excluyentes de la posible acción antrópica dentro del conjunto óseo, aspecto que dependerá de un mayor cúmulo de evidencias.
Sin lugar a dudas, este hallazgo es único por su naturaleza y estado de conservación; sin embargo, su similitud con otros yacimientos hace necesario ahondar los análisis, puesto que habría una serie de indicadores como la diversidad taxonómica, la concentración de los restos, la presencia de fracturas en hueso frescos, entre otros, que nos llevan a indagar su posible asociación con ocupaciones humanas.
Agradecimientos
El presente trabajo fue financiado por GNL Quintero S. A. y Arka Consultores. Los autores desean agradecer al arqueólogo Joaquín Vega por la información y ayuda aportada. Especial agradecimiento a los organizadores del Primer Encuentro Latinoamericano de Zooarqueología, en particular a la doctora Elizabeth Ramos, por la ayuda y facilidades brindadas. Por último, agradecemos a los evaluadores anónimos, que ayudaron a mejorar el presenta manuscrito. Los errores de este trabajo son de exclusiva responsabilidad de los autores. .
Referencias
Behrensmeyer, Anna Kay
1978. "Taphonomic and Ecologic Information from Bone Weathering", Paleobiology 4 (2), pp. 150-62.
Behrensmeyer, Anna Kay y Andrew Hill
1980. Fossils in the Making: Vertebrate Taphonomy and Paleoecology. Chicago, University of Chicago Press.
Binford, Lewis
1981. Bones: Ancient Men and Modern Myths. Nueva York, Academic Press.
Bishop, Walter W.
1980. "Paleogeomorphology and Continental Taphonomy", en Fossils in the Making, eds. Anna K. Behrensmeyer y Andrew P. Hill, pp. 20-40. Chicago, Prehistoric Archaeology and Ecology Series, University of Chicago Press.
Blanton, Dennis
1996. "Accounting for Submerged Mid-Holocene Archaeological Sites in the Southeast. A Case Study from the Chesapeake Bay Estuary, Virginia", en Archaeology of the Mid-Holocene Southeast, eds. Kenneth E. Sassaman y David G. Anderson, pp. 200-217. Gainesville, University Press of Florida.
Blumenschine, Robert J.
1995. "Percussion Marks, Tooth Marks, and Experimental Determinations of the Timing of Hominid and Carnivore Access to Long Bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania", Journal of Human Evolution 29, pp. 21-51.
Bonnichsen, Robson
1982. "Bone Technology as a Taphonomic Factor: An Introductory Statement", Canadian Journal of Anthropology 2 (2), pp. 137-144.
Bonnichsen, Robson, Bradley Lepper, Dennis Stanford y Michael Waters
2005. Paleoamerican Origins: Beyond Clovis. Orono, Center for the Study of the First Americans, Texas University Press.
Borrero, Luis A.
2003. "Taphonomy of the Tres Arroyos Rockshelter", Quaternary International 109-110, pp. 87-93.
Borrero, Luis A. y Fabiana Martin
1996. "Tafonomía de carnívoros: un enfoque regional", en Arqueología Solo Patagonia. Segundas Jornadas de Arqueología de la Patagonia, ed. Julieta Gómez Otero, pp. 189-198. Puerto Madryn, Centro Nacional Patagónico.
Bouzas, Ana y Ana Laborde
2003. "La degradación del hueso", Monte Buciero 9, pp. 267-276.
Bradley, Bruce y Dennis Stanford
2004. "The north Atlantic Ice-edge Corridor: A Possible Paleolithic Route to the New World", World Archaeology 36 (4), pp. 459-478.
Bunn, Henry T.
1993. "Bone Assemblages at Base Camps: A Further Consideration of Carcass Transport and Bone Destruction by the Hadza", en From bones to behavior: Etnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains, ed. Jean Hudson, pp. 156-68. Illinois, Southern Illinois University at Carbondale.
Bunn, Henry. T., Laurence Bartram y Ellen Kroll
1988. "Variability in Bone Assemblage Formation from Hadza Hunting, Scavenging, and Carcass Processing", Journal of Anthropological Archaeology 7, pp. 412-457.
Carabias, Diego
2001. "Primeras aproximaciones a través de la arqueología subacuática al pasado prehistórico del Norte chico: Punta Ñagué", Boletín de la Sociedad Chilena de Arqueología 31, pp. 39-41.
2005. "Patrimonio cultural subacuático de Chile. El rescate de un legado sumergido en Chile", en País oceánico, pp. 124-131. Santiago, Ocho Libros Editores. [ Links ]
Carabias, Diego
2008. "Informe de evaluación arqueológica subacuática e implementación de área de exclusión preventiva en torno al sitio GNLQ1", en Proyecto Terminal de Gas Natural Licuadi (GNL) en Quintero, VRegión, Comuna de Quintero. Valparaíso.
Casamiquela, Rodolfo
1984. "Critical Catalogue of Some Chilean Fossil Vertebrates I. The Deers: Complementary Considerations on Antifer (Antifer niemeyeri n. sp.), the Pleistocene Giant Deer", Quaternary of South America & Antarctic Peninsula 2, pp. 41-50.
1999. "The Pleistocene Vertebrate Record of Chile", Quaternary of South America & Antarctic Peninsula 7, pp. 91-107. [ Links ]
Clausen, Carl J., Arthur Cohen, Cesare Emiliani, Alan Holman y Jerry Stipp
1979. "Little Salt Spring Florida: A Unique Underwater Site", Science 203 (4381), pp. 609-614.
De Ruiter, Darryl y Lee Berger
2000. "Leopards as Taphonomic Agents in Dolomitic Caves. Implications for Bone Accumulations in the Hominid-bearing Deposits of South Africa", Journal of Archaeological Science 27, pp. 665-684.
Dean, Martin, Ben Ferrari, Ian Oxley, Mark Redknap y Kittie Watson
2000. Archaeology Underwater. The NAS Guide to Principles and Practice. Londres. Editorial Archetype Publications Ltd.
Dillehay, Tom
2009. "Probing Deeper into First American Studies", Proceedings of the National Academy of Sciences 106 (4), pp. 971-978.
Domínguez-Rodrigo, Manuel
1999. "Flesh Availability and Bone Modifications in Carcasses Consumed by Lions", Palaeogeography, Palaeoclimatology, Palaeoecology 149, pp. 373-388.
Dunbar, James, David Webb y Dan Cring
1989. "Culturally and Naturally Modified Bones from a Paleoindian Site in the Aucilla River, North Florida", en Bone Modifications, eds. Robson Bonnichsen y Marcella Sorg, pp. 473-497. Orono, Center for the Study of the First Americans.
Dunbar, James, David Webb y Michael Faught
1992. "Inundated Prehistoric Sites in Apalachee Bay, Florida, and the Search for the Clovis Shoreline", en Paleoshorelines and Prehistory: An Investigation of Method, eds. Lucille Johnson y Melanie Stright, pp. 117-148. Ann Arbor, CRC Press.
Faught, Michael
1996. "Clovis Origins and Underwter Prehistoric Archaeology in Northwestern Florida" Ph.D. dissertation, University of Arizona, Tucson. University Microfilms, Ann Arbor.
2004a. "The Underwater Archaeology of Paleolandscapes, Apalachee Bay, Florida", American Antiquity 69 (2), pp. 275-289. [ Links ]
2004b. "Submerged Paleoindian and Archaic Sites of the Big Bend, Florida", Journal of Field Archaeology 29 (3-4), pp. 273-290. [ Links ]
Faught, Michael y Joseph Donoghue
1997. "Marine Inundated Archaeological Sites and Paleofluvial Systems: Examples from a Karst-controlled Continental Shelf Setting in Apalachee Bay, Northeastern Gulf of Mexico", Geoarchaeology 12 (5), pp. 417-458.
Flemming, Nic C.
2004. "Submarine Prehistoric Archaeology of the Indian Continental Shelf: A Potential Resource", Current Science 86 (9), pp. 1225-1230.
Frison, George
1982. "Bone Butchering Tools in Archaeological Sites", Canadian Journal of Anthropology 2 (2), pp. 159-167.
Galili, Ehud, Mina Weinstein-Evron, Israel Hershkovitz, Avi Gopher, Kislev Mordecai, Omri Lernau, Liora Kolska-Horwitz y Hanan Lernau
1993. "Atl it-Yam: A Prehistoric Site on the Sea Floor Off the Israeli Coast", Journal of Field Archaeology 20 (2), pp. 133-157.
Goebel, Ted, Michael Waters y Dennis O'Rourke
2008. "The Late Pleistocene Dispersal of Modern Humans in the Americas", Science 319, pp. 1497-1502. Gusick, Amy y Michael Faught
2011. "Prehistoric Archaeology Underwater: A Nascent Subdiscipline Critical to Understanding Early Coastal Occupations and Migration Routes", en Trekking the Shore, Changing coastlines and the antiquity of coastal settlements, eds. Nuno F. Bicho, Jonathan A. Haws y Loren G. Davis, pp. 27-50. Nueva York, Springer.
Hanson, C. Bruce
1980. "Fluvial Taphonomic Processes: Models and Experiments", en Fossils in the Making, eds. Anna K. Behrensmeyer y Andrew P. Hill, pp. 156-181. Prehistoric Archaeology and Ecology Series. Chicago, University of Chicago Press.
Haynes, Gary
1983a. "A Guide for Differentiating Mammalian Carnivores Taxa Responsable for Gnaw Damage to Herbivore Limb Bones", Paleobiology 9 (2), pp. 164-172. 1983b. "Frequencies of Spiral and Greenbone Fractures on Ungulate Limb Bones in Modern Surface Assemblages", American Antiquity 48, pp. 102-114. [ Links ]
Hoffecker, John, Roger Powers y Ted Goebel
1993. "The Colonization of Beringia and the Peopling of the New World", Science 259, pp. 46-53.
Hill, Andrew y Anna Kay Behrensmeyer
1985. "Natural Disarticulation and Bison Butchery", American Antiquity 50 (1), pp. 141-145.
Jackson, Donald, César Méndez, Patricio López, Douglas Jackson y Roxana Seguel
2005. "Evaluación de un asentamiento arqueológico en el semiárido de Chile: procesos de formación, fauna extinta y componentes culturales", Intersecciones en Antropología 6, pp. 139-152.
Jackson, Donald, César Méndez, Roxana Seguel, Antonio Maldonado y Gabriel Vargas
2007. "Initial Occupation of the Pacific Coast of Chile during Late Pleistocene Times", Current Anthropology 48 (5), pp. 725-731.
Josenhans, Heiner, Daryl Fedje, Reinhard Pienitz y John Southon
1997. "Early Humans and Rapidly Changing Holocene Sea Levels in the Queen Charlotte Islands-Hecate Strait British Columbia, Canada", Science 277, pp. 71-74.
Labarca, Rafael y M. Alejandra Alcaraz
2011. "Presencia de Antifer ultra Ameghino (=Antifer niemeyeri Casamiquela) (Artiodactyla, Cervidae) en el Pleistoceno tardío-Holoceno Temprano de Chile Central (30-35° S)", Andean Geology 38, pp. 156-170.
Lambeck, Kurt, Emma-Kate Potter y Tezer Esat
2002. "Links between Climate and Sea Levels for the Past Three Million Years", Nature 419, pp. 199-206.
López, Patricio
2007. "Tafonomía de los mamíferos extintos del Pleistoceno Tardío de la costa meridional del semiárido de Chile (IV Región-32° latitud): alcances culturales y paleoecológicos", Chungara 39 (1), pp. 69-86.
López, Patricio, Rafael Labarca y Lautaro Núñez
2004. "Nivel Quereo I: una discusión acerca del poblamiento temprano en la Provincia del Choapa", Revista Werkén 5, pp. 15-20.
Masters, Patricia M. y Nic C. Flemming
1983. Quaternary Coastlines and Marine Archaeology. Towards the Prehistory of Land Bridges and Continental Shelves. Nueva York, Academic Press.
Myers, Thomas, Michael Voorhies y Georde Corner
1980. "Spiral Fractures and Bone Pseudotools at Paleontological Sites", American Antiquity 45 (3), pp. 483-490.
Noakes, Scott, Ervan Garrison y Greg McFall
2009. "Underwater Paleontology: Recovery of a Prehistoric Whale Mandible Offshore Georgia", en Proceedings of the American Academy of Underwater Sciences 28th Symposium, ed. N. W. Pollock, pp. 245-251. Atlanta, American Academy of Underwater Sciences.
Núñez, Lautaro, Juan Varela, Rodolfo Casamiquela y Carolina Villagrán
1994. "Reconstrucción multidisciplinaria de la ocupación prehistórica de Quereo, Centro de Chile", Latin American Antiquity 5 (2), pp. 99-118.
O'Connell, James, Kristen Hawkes y Nicholas Blurton Jones
1990. "Reanalysis of Large Mammal Body Part Transport among the Hadza", Journal of Anthropological Research 17, pp. 31-316.
Petriaggi, Roberto y Barbara Davidde
2007. Archeologia sott'acqua. Teoria y practica. Pisa, Accademia Editoriale.
Rinderknecht, Andrés
2000. "La presencia de osteodermos en las extremidades posteriores de Glyptodon clavipes Owen, 1839 (Mammalia: Cingulata)", Ameghiniana 37 (3), pp. 369-373.
Stewart, David
1999. "Formation Processes Affecting Submerged Archaeological Sites: An Overview" en Geoarchaeology 14 (6), pp. 565-587.
Stright, Melanie
1995. "Archaic Period Sites on the Continental Shelf of North America: The Effect of Relative Sea-level Changes on Archaeological Site Locations and Preservation", en Archaeological Geology of the Archaic Period in North America, ed. Elmer A. Bettis, pp. 131-147. Geological Society of America Special Paper 297.
Vargas, Gabriel y Cristina Ortega
2008. "Contexto geomorfológico y análisis sedimentológico preliminar de muestras de la bahía Quintero, V Región", en Proyecto Terminal de Gas Natural Licuado (GNL) en Quintero, V Región, Comuna de Quintero. Valparaíso.
Waters, Michael
1992. Principles of Geoarchaeology: A North American Perspective. Tucson, The University of Arizona Press.

