











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkLa provincia de Santa Fe, y principalmente la llanura aluvial del Paraná, contó con pioneras investigaciones arqueológicas desde fines del siglo XIX y durante el XX, debido a que la zona resultaba atrayente, por cuanto demostraba un gran potencial, dada la cantidad de registro que podía observarse en superficie. Tales investigaciones derivaron en el planteo de diversos modelos de ocupación para el área del Nordeste argentino (en adelante, NEA) (De Aparicio 1936; Frenguelli 1923; Outes 1918; Serrano 1931; Zeballos y Pico 1878). Así mismo, en los últimos años se produjo una mayor diversificación del conocimiento al surgir nuevos enfoques que derivaron en la generación de nuevos interrogantes y la aproximación a nuevas problemáticas.
Una de las temáticas que comenzaron a desarrollarse en la provincia de Santa Fe a comienzos del año 2000 fue la zooarqueología, realizándose análisis de conjuntos faunísticos, aunque provenientes generalmente de un solo depósito (Feuillet Terzaghi 2002; Pérez Jimeno 1996; Santiago 2003, entre otros). Recién a partir de los primeros resultados obtenidos a nivel de sitio comenzó a integrarse la información intersitios, lo que permitió realizar inferencias acerca de la subsistencia, los rangos de acción y uso de los recursos (Acosta et al. 2010; Cornaglia 2009; Cornero, Solomita y Curetti 2010; Mucciolo y Pérez Jimeno 2015; Pérez Jimeno 2007; Santiago 2003; Sartori 2008; 2013; Sartori y Colasurdo 2011; Sartori, Colasurdo y Santiago 2014; Sartori y Galligani 2014). En tal sentido, se considera que el presente trabajo constituye un aporte para el desarrollo de investigaciones regionales al permitir comparar el registro arqueofaunístico de doce sitios localizados en diferentes ambientes de la provincia de Santa Fe (ver la tabla 1y la figura 1).
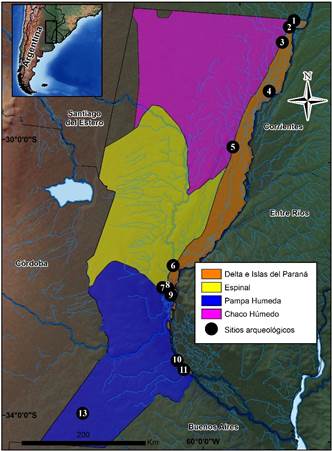
Fuente: los autores. Los números corresponden con la columna de la tabla 1.
Figura 1 Mapa de Santa Fe con los conjuntos zooarqueológicos analizados
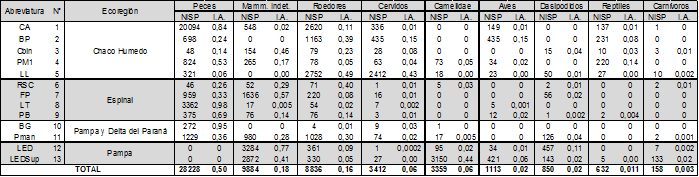
Tabla 1 Localización de los sitios, fechados radiocarbónicos y número de especímenes identificados por taxón (NISP) de cada uno
| Abreviatura | N° | Latitud | Longitud | Sitio | Fechado | NISP | Referencia |
| TOTAL | |||||||
| CA | 1 | 28°0'52.99" | 59°5'41.96" | Cerro Aguara | 530±70 | 23885 | Pérez Jimeno 2005 Santiago 2004 |
| BP | 2 | 28°6'45.26" | 59°9'32.61" | Barrancas del Paranacito | S/d. | 2962 | Pérez Jimeno 1996 |
| Cbin | 3 | 28°20'19.33" | 59°15'15.60" | Campo Binaghi | S/d. | 337 | Sartori y Pérez Jimeno 2012 |
| PM1 | 4 | 29°3'28.43" | 59°24'17.91" | Parana Mini 1 | S/d. | 1557 | Schmitz et al. 1972 |
| LL | 5 | 29°54'8.48" | 59°55'37.92" | La Lechuza | 1760±60 | 5613 | Cornero, Solomita y Curetti 2010 |
| RSC | 6 | 31°40'42.23" | 60°46'22.88" | Río Salado Coronda II | 1305±48 | 179 | Sartori 2008, 2010a |
| FP | 7 | 32°0'26.26" | 60°55'25.96" | Familia Primón | 370±30 | 2887 | Sartori 2008, 2010a |
| LT | 8 | 32°0'24.69" | 60°50'8.38" | Las Tejas | 1014±45 | 3445 | Sartori 2013 |
| PB | 9 | 32° 6' 37" | 60º 44' 35" | Pajas Blancas | 506±43 | 545 | Sartori 2010b |
| BG | 10 | 33°2'28.72" | 60°34'50.96" | Bajada Guereño | S/d. | 286 | Escudero y Feuillet 2002 |
| Pman | 11 | 33°9'56.73" | 60°27'5.63" | Playa Mansa | 2400±20 | 3456 | Sartori y Colasurdo 2011 |
| LED | 12 | 33°54'20'' | 62°08'43'' | Laguna El Doce | 8274±64 | 4239 | Cornaglia 2014 |
| LEDSup | 13 | Laguna El Doce superficie | 1555±85 | 7081 | Cornaglia 2013 | ||
| TOTAL NISP | 56472 | ||||||
Fuente: los autores.
Considerando que la mayor parte de los análisis zooarqueológicos en la provincia de Santa Fe son a nivel microrregional, este trabajo, al realizarse desde una escala regional, pretende integrar la información zooarqueológica para poder comprender cómo se estructuran los distintos conjuntos arqueofaunísticos en las distintas unidades del paisaje que componen la provincia. La aplicación de SIG para este análisis brinda modos gráficos, y más sencillos, de visualizar e interpretar los datos provenientes de los conjuntos faunísticos en términos de sus variaciones espaciales.
Ambiente
La provincia de Santa Fe, al situarse en la llanura chaco-pampeana, carece de accidentes orográficos importantes; sin embargo, su paisaje se desarrolla con suaves ondulaciones y con un fuerte modelado de los rí os de llanura que la surcan. Con base en las caracterí sticas de relieve y el gradiente de temperatura se pueden diferenciar seis ecorregiones. Los sitios que se tratan en este trabajo se ubican, mayormente, en sectores de transición entre el Delta e Islas del Paraná con la región Pampeana (PDP), el Espinal (ESP) y el Chaco Húmedo (CHH). La excepción la constituye el sitio localizado al suroeste de la provinc ia, el cual se emplaza exclusivamente en la región Pampeana (PAM). Cada una de las zonas posee particularidades en la composición florística y faunística que las caracteriza. A continuación se detallan las generalidades ambientales que poseen los cuatro sectores (PDP, ESP, CHH y PAM) en los cuales se localizan los sitios arqueológicos que permiten generar expectativas arqueológicas respecto a la composición de los conjuntos arquefaunísticos en cada una de las zonas por analizar. Los sitios que se emplazan más hacia el norte corresponden al Chaco Húmedo (CHH), y zoogeográficamente pertenecen a la Región Neotropical y al subdistrito chaqueño, el cual es el más rico por su gran biodiversidad (Cabrera y Yepes 1940). Se caracteriza por la presencia de gran cantidad de especies, como el mono carayá (Alouatta caraya), el ocelote (Felis pardalis), el aguará guazú (Chrysocyon brachyurus), la corzuela colorada (Mazama gouazoubira) y el tapir (Tapirus terrestris), entre otros. En cuanto a los peces, predominan los dos órdenes más representativos, correspondientes a los charácidos y silúridos. Los sitios del Espinal (ESP), en términos zoogeográficos, pertenecen al Dominio Pampásico (Subregión Guayano-brasileña, Región Neotrópica) (Ringuelet 2004) y abarcarían tanto al sector Pampásico como al de transición entre este último y el sector de Delta e Islas del Paraná (Peña 1997). Ambos ambientes se caracterizan por poseer especies pampeanas típicas, tales como Cavia aperea y Chaetophractus villosus; pero también especies características que se encuentran adaptadas a la vida acuática. Dentro de esta pueden mencionarse como los más destacados al carpincho (Hydrochoerus hydrochaeris) y el coipo (Myocastor coypus). Las aves más características de la zona son el leñatero (Anumbius annumbi), los espineros (Phacellodomus sp.) y los pijuíes (Synallaxis sp.), y también los lechuzones del campo (Asio flammeus). Cabe señalar que, como se mencionó anteriormente, la mayoría de los sitios arqueológicos que se emplazan en las dos ecorregiones descriptas (CHH y ESP) se hallan en realidad en la planicie de inundación del Paraná, es decir, en la intersección con la ecorregión del Delta e Islas del Paraná. Esto se debe a que en el sector este de la provincia de Santa Fe, el ESP y el CHH se ven atravesados por el tramo medio del río Paraná, que se extiende desde la confluencia del río Paraguay hasta las inmediaciones de la ciudad de Rosario. Con una longitud de cauce de unos 800 km, atraviesa todo el sector este de la provincia de Santa Fe (Ramonell et al. 2011). La planicie de inundación del tramo medio del Paraná es compleja, ya que se encuentra formada por un mosaico de unidades geomorfológicas que se disponen a cotas diferentes entre sí y se encuentran atravesadas por cauces de distinta conectividad con el principal del sistema (Bosisio y Ramonell 2014). Debido a esto es que poseen una alta productividad ambiental, ecológicamente regulada por los pulsos de inundación del río (Bó y Malvárez 1999; Neiff 1999). Este ejerce un efecto climático que se traduce en la presencia de una cuña del Dominio Amazónico, el cual permite la penetración de poblaciones animales típicamente amazónicas hacia el sur (Cabrera 1971; Ringuelet y Aramburu 1957). De esta forma, la fauna característica de toda la zona incluye especies subtropicales que se caracterizan por estar adaptadas al ámbito fluvial, como el ciervo de los pantanos (Blastocerus dichotomus), el coipo (Myocastor coypus), el carpincho (Hydrochoerus hydrochaeris), el lobito de río (Lontra longicaudis), yacaré (Caiman sp.) y tortuga de río (Phrynops sp. e Hydromedusa sp.) (SPANP 1997). Otro aspecto característico de todo el sector es la alta concentración de peces, representados por dos órdenes: Characiformes y Siluriformes. Entre los primeros se encuentran el dorado (Salminus maxillosus), sábalo (Prochilodus platensis), boga (Leporinus obtusidens), tararira (Hoplias malabaricus), pacú (Piaractus mesopotámicus), y las mojarras (Aphyocharax sp., Astyanax sp.); y entre los Siluriformes, el surubí (Pseudoplatystoma coruscans), patí (Luciopimelodus pati), bagre (Pimelodus clarias y Rhamdia sapo), armado (Pterodoras granulosus), moncholo (Pimelodus albicans) y vieja del agua (Plecostomus commersoni) (Ringuelet 2004). Por último, la fauna de región Pampeana (PAM) está caracterizada por especies pampeanas que poseen cierta plasticidad adaptativa, como el venado de las pampas (Ozotoceros bezoarticus), el guanaco (Lama guanicoe), el puma (Puma concolor), la comadreja (Didelphis albiventris) y cuis (Cavia aperea). En el orden de los roedores, también se encuentran la vizcacha (Lagostomus maximus) y el tuco-tuco (Ctenomys sp.) (Gollán 1958). Entre los carnívoros se encuentran el zorrino común (Conepatus humboldtii), el zorro pampeano (Lycalopex gymnocercus), el hurón (Galictis cuja) y el gato montés (Oncifelis geoffroyi) (Gollán 1958). También se hallan diversos edentados como el pichiciego (Chlamyphorus truncatus), el quirquincho (Tolypeutes matacus), el peludo (Chaetophractus villosus) y la mulita (Dasypus hybridus) (Gollán 1958; Manzi y Gallardo 1970). Las aves que predominan pertenecen al orden de los Ciconiiformes (Ciconiae y Ardeae), y también a las familias de los anímidos y anátidos, y a la familia Rheidae (Rhea americana) (Manzi y Gallardo 1970).
El registro arqueofaunístico de la provincia de Santa Fe
Cada uno de los ambientes de la provincia de Santa Fe posee datos variados respecto a los análisis arqueofaunísticos. Considerando que el objeto de este apartado es sintetizar la información generada para cada paisaje, aquí se exponen de manera resumida los trabajos que realizan análisis en profundidad y que corresponden a la zona del Chaco Húmedo (CHH) (Cornero, Solomita y Curetti 2010; Pérez Jimeno 2007; Mucciolo y Pérez Jimeno 2015; Santiago 2004; Sartori y Pérez Jimeno 2012), el Espinal (ESP) (Sartori 2008; 2010a; 2012; 2013; Sartori y Galligani 2014)1, la Pampa (PAM) (Cornaglia 2012) y la zona de la pampa en su intersección con el Delta e Islas del Paraná (PDP) (Escudero y Feuillet 2007; Feuillet 2002; Sartori y Colasurdo 2011) (ver la figura 2).
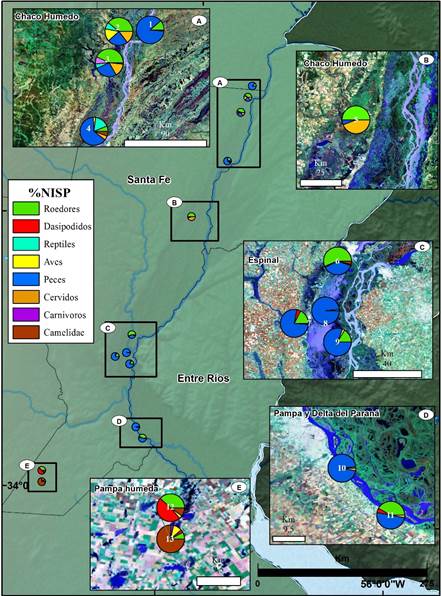
Fuente: los autores.
Figura 2 Localización de los sitios y representación de los conjuntos en términos de porcentaje de Número de especímenes identificados (%NISP) por categorías utilizadas en este trabajo
Cabe realizar una distinción respecto a los sitios que se emplazan en el ámbito ribereño de las zonas PDP, ESP y CHH vs. aquellos que pertenecen a la llanura pampeana santafesina. Si bien es abundante la cantidad de sitios pertenecientes a este último ambiente (Ceruti 1987), no hay una publicación de análisis zooarqueológicos acorde. En tal sentido, si bien Ceruti estudió sitios que formarían parte de lo que denomina "entidad cultural esperanza" (Ceruti 1984), no existen estudios que abarquen en profundidad al componente arqueofaunístico de los registros, por lo que no pueden ser incluidos aquí (Ceruti 1984; 1987; 2000; 2004). El único sitio en el cual sí se ha realizado un estudio en detalle corresponde a Laguna el 12 (LED12) (Cornaglia 2013; 2014). Este sitio posee diversos fechados, entre los cuales el más antiguo es el de 8274±68 años 14C AP (dientes humanos, 7452-7396 años cal AC, AA-89915; δ13C=-15,1‰). Por su parte, la ocupación más reciente corresponde al Holoceno tardío, y cuenta con dos fechados obtenidos sobre tiestos cerámicos, que arrojaron una antigüedad de 1555±85 14C AP (AA-89918; 420-587 cal AC) y 2350±180 14C AP (AA-89919; 753-685 cal AC) (Avila 2011). Tales dataciones sitúan a este sitio como el más antiguo de la provincia de Santa Fe (Avila 2011). En cuanto a las inferencias del uso del espacio, Cornaglia sostiene que, considerando que la fauna recuperada corresponde a especies de estirpe central y/o patagónica, los grupos cazadores que ocuparon el sitio habrían desarrollado estrategias adaptativas adecuadas para desenvolverse en un ambiente árido-semiárido (Cornaglia 2014). En los conjuntos recuperados de este sitio, tanto en superficie como en estratigrafía, se hallan presentes Lama guanicoe, Rhea americana y Ozotoceros bezoarticus, especies que constituyeron una parte importante de la dieta de los grupos humanos. La dominancia de Lama guanicoe con respecto a las demás es un indicador de que esta fue la principal fuente de proteínas animales para los distintos grupos humanos durante el Holoceno (Cornaglia 2009; 2013; 2014).
Por su parte, en la ecorregión pampeana, pero en su transición con el Delta e Islas del Paraná (sureste de la provincia de Santa Fe), hay análisis realizados en profundidad acerca de la fauna, pero no se llevaron a cabo planteos de modelos regionales. Esto se debe fundamentalmente a que en la zona sólo se han localizado dos yacimientos, y, por ende, se han realizado estudios a nivel de sitio (Escudero y Feuillet 2007; Feuillet 2002). Estos se centran en el sitio Playa Mansa, que hasta el momento constituye el depósito más antiguo (2400 ± 20 AP) de la planicie de inundación del Paraná en su tramo medio e inferior (Sartori y Colasurdo 2011). En este sitio, los análisis faunísticos habrían estado orientados a discutir las distintas trayectorias de ingreso al registro arqueológico, y también la importancia relativa de cada especie. Las autoras concluyen que en Playa Mansa se constató el consumo por parte humana de diferentes especies, con un aprovechamiento de recursos tanto de alta como de baja jerarquía. Entre los primeros se encuentran Blastocerus dichotomus, Ozotoceros bezoarticus y Mazama gouazoubira, y entre los segundos pueden ubicarse Myocastor coypus, Cavia aperea, dasipódidos y peces (Sartori y Colasurdo 2011).
En cuanto al sector más al norte de la provincia, en el Chaco Húmedo, el depósito más antiguo corresponde al sitio La Lechuza, que posee un fechado radiocarbónico de 1760 ± 60 años AP (Cornero, Solomita y Curetti 2010). El modelo de ocupación humana para esa zona ha sido planteado por Pérez Jimeno (2007) para grupos que ocuparon el área, aunque con posterioridad a 1500 años AP. La autora sostiene que habría estado ocupada por grupos humanos semisendentarios, vinculados estrechamente a los ambientes lénticos generados por la dinámica del río Paraná. Los grupos, a su vez, habrían contado con una tecnología especializada (cerámica, ósea y lítica en ciertos sectores), que les permitía basar su subsistencia en la pesca, la caza de mamíferos (fundamentalmente, roedores y ciervos) y la recolección de moluscos. Los ambientes lénticos serían explotados intensivamente durante los períodos de bajante del río, ya que a comienzos de los períodos de inundación se fraccionaban y desplazaban hacia el oeste, hacia áreas ecotonales, o las tierras altas de la margen izquierda del río Paraná (Pérez Jimeno 2007).
En los conjuntos de los sitios que fueron estudiados por Pérez Jimeno y colaboradores se refleja una explotación de cérvidos, tales como Blastocerus dichotomus y Ozotoceros bezoarticus, roedores (Myocastor coypus, Cavia aperea e Hydrochoerus hydrochaeris) y peces (Pérez Jimeno 2007; Mucciolo y Pérez Jimeno 2015; Santiago 2002). Sin embargo, hay algunas diferencias respecto a la composición de los conjuntos, que ha sido explicada en términos espaciales (Sartori y Pérez Jimeno 2012). Si bien se estima que la pesca constituyó una actividad importante para los habitantes de este sector, que habrían explotado este recurso en los cuerpos de agua más próximos, y no en el cauce principal del río Paraná (Pérez Jimeno 2007), es interesante que en los sitios ubicados en espacios fuera de la planicie de inundación decrece la dominancia de los peces en los conjuntos (Sartori 2013).
Para el Espinal hay planteadas líneas de investigación vinculadas a ciertas expectativas arqueológicas que están siendo desarrolladas por una de las autoras de este manuscrito. Estas sugieren que ca. 1000 años AP, y hasta el momento de contacto entre europeos y pueblos originarios, habría habido una recurrencia en la ocupación de espacios insulares y sectores de borde de la planicie de inundación del Paraná. Estos últimos se ubicarían en terrenos altos, con una buena visibilidad, situados próximos a cursos de agua, pero no inundables. Esto permitiría el abastecimiento de recursos críticos como agua, leña y gran cantidad de vegetales y animales potencialmente consumibles (Sartori 2008; 2010a).
Por su parte, los espacios insulares serían intensamente ocupados, debido al acceso a los recursos faunísticos que propiciarían, fundamentalmente la obtención de peces en ambientes de bajo riesgo y mayor concentración. Es decir que se daría una jerarquización en la ocupación de ciertos tipos de espacios en detrimento de otros, los cuales serían ocupados recurrentemente a lo largo del tiempo y presentarían una demarcación simbólica, establecida fundamentalmente por la presencia de entierros múltiples (Sartori 2013).
En cuanto a los recursos faunísticos para la zona, estos reflejan la diversidad de taxa aprovechados, entre los cuales se encuentran aquellos que son de alta jerarquía (i.e. B. dichotomus, O. bezoarticus y H. hydrochaeris) y de baja jerarquía (i.e. M. coypus, dasipódidos y peces). También pudo corroborarse en los diferentes conjuntos el aprovechamiento de siete especies, considerando los mamíferos y las aves, que habrían sido aprovechados con seguridad, debido a que presentan atributos vinculados a la actividad antrópica (huellas y/o alteraciones térmicas). Por otra parte, si dentro de este conteo se incluye al macrotaxón de los peces, el número se incrementa aún más, ya que se observaron diez especies en los diferentes registros, entre las cuales las más representadas corresponden a bagres (Pimelodus albicans y Pimelodus maculatus) y armado (Pterodoras granulosus). La composición de los conjuntos permite realizar inferencias basadas en la abundancia y presencia/ausencia de ciertos taxa. En tal sentido, la variabilidad de especies presentes indica un predominio de peces en los sitios insulares, mientras que los sitios del borde occidental de la llanura aluvial reflejan un mayor aprovechamiento de mamíferos. De todos modos, en ambos casos se estaría optando por estrategias que reducen o minimizan el riesgo al incluir presas de baja jerarquía (coipo, dasipódidos, peces) en la dieta (Sartori 2013).
Metodología
El análisis que se realiza en este trabajo es de escala regional, ya que abarca distintos sectores de la provincia de Santa Fe. Para este fueron relevados los datos provenientes de trece conjuntos provenientes de doce yacimientos arqueológicos localizados en diferentes zonas. La definición de cada conjunto fue establecida en función de la información que se encuentra publicada por los autores que realizaron los análisis para cada muestra, considerándose como requisito que estas se compongan de más de cien especímenes. Para el análisis se descartaron los especímenes no identificados (Número de especímenes indeterminados-NUSP) (Lyman 2008) o indeterminados y se agruparon los taxa en nueve categorías amplias. Para el estudio de las arqueofaunas se consideró sólo al macrotaxón vertebrados; siendo excluidos para los fines de este trabajo aquellos restos que pertenecen a la categoría de invertebrados. En la tabla 2 se enumera el Número de especímenes identificados por taxón (NISP) de cada conjunto, los índices de abundancia calculados (IA), las fuentes bibliográficas de referencia y el fechado radiocarbónico para cada uno.
Tabla 2 Datos zooarqueológicos de sitios de la Provincia de Santa Fe, considerados por ecorregión, NISP por taxones analizados, índices de abundancia calculados y referencias bibliográficas
Fuente: los autores.
Metodológicamente, para poder generar un análisis que contribuyera a evaluar tendencias en términos espaciales se realizó la agregación de categorías taxonómicas, las cuales quedaron englobadas bajo el nivel de Familia o incluso Clase (siguiendo la propuesta de Lyman 2003). Una vez realizado esto se procedió a georreferenciar los sitios de los cuales procedían los conjuntos faunísticos y conformar nuevos conjuntos por medio del agrupamiento de nueve taxa en nuevas categorías (Peces, Roedores, Mammalia Indeterminados, Cérvidos, Reptiles, Aves, Dasipodidae, Camelidae, Carnivoridae). De ellos se calcularon los índices de abundancia (IA) siguiendo lo propuesto por Lyman (2003), para lo cual se empleó la siguiente fórmula:
∑NISPpeces
(∑NISPpeces+∑NISPaves+∑NISPMann.Indet.+∑NISProedores+∑NISPdasipódidos+∑NISPreptiles+∑NISPCamelidae+∑NISPcérvidos+∑NISPCarnivoridae)
Entre los muchos dispositivos analíticos incorporados en un programa SIG, podemos encontrar la geoestadística, un conjunto de herramientas que pueden ser utilizadas para múltiples usos. Nosotros lo utilizamos para conocer mejor la variación espacial de datos zooarqueológicos, por medio de predicciones fiables de atributos muestreados en ubicaciones no muestreadas. Es decir, tomamos los datos zooarqueológicos de ubicaciones puntuales y obtenemos predicciones o probabilidades para sectores del espacio del cual no tenemos datos. Este procedimiento se denomina Interpolación, que es la estimación del valor de una variable en una ubicación no medida con los valores observados en los alrededores. Por medio de la interpolación construimos nuevos datos a partir de un conjunto discreto de puntos de datos conocidos (las arqueofaunas de los sitios analizados).
Mediante el programa ArcGIS 10.1 se generaron mapas en formato raster, denominados Bonescapes, según el procedimiento de Santiago y Vázquez (2013), derivado de Isoscapes: Isotopic Landscapes (sensu West et al. 2010). Estos mapas raster se realizaron por medio de interpolación de datos, con el procedimiento ordinario, llamado Ponderación de las distancias inversas (IDW, por su sigla en inglés). Finalmente se procedió a recortar los mapas raster (estos mapas son superficies continuas, donde cada pixel contiene un dato numérico; en el caso de los Bonescapes, los datos son los valores derivados de los índices de abundancia) con un mapa vectorial de la provincia de Santa Fe. La generación de Bonescapes con datos zooarqueológicos nos permite modelar el uso espacial de los recursos. Estos mapas pueden ayudarnos a interpretar patrones espaciales en el aprovechamiento humano de los animales en toda la provincia de Santa Fe, así como generar nuevos paisajes óseos, con la incorporación de datos de zonas vecinas.
Debe tenerse en cuenta que la información faunística para la planicie de inundación del Paraná en la provincia de Santa Fe es amplia, aunque el nivel de análisis con que se han tratado los diferentes conjuntos es heterogéneo, con lo cual resulta un desafío poder realizar comparaciones de grano fino o análisis integradores. Por lo expuesto, y acorde a los objetivos del presente trabajo, se decidió realizar el análisis sobre la base del NISP, lo cual permitirá incluir una mayor cantidad de datos. No obstante, se es consciente de las dificultades de derivar directamente de estos valores patrones de subsistencia, pero se considera que el NISP es la única medida de abundancia taxonómica disponible en la gran mayoría de los análisis, y cuyo cálculo es unívoco, lo cual resulta adecuado para fines comparativos (Grayson 1984; Lyman 1994; 2003; 2008). Además, el NISP tiene más ventajas a la hora de realizar comparaciones, tales como las que planteamos en el presente trabajo, que desventajas. Es por ello que consideramos al NISP como una medida robusta en sí misma y óptima para poder realizar este tipo de comparaciones intersitio a nivel regional, ya que otras medidas zooarqueológicas tradicionales (Número mínimo de elementos-MNE y Número mínimo de individuos-MNI) acarrean mayores diferencias a la hora de calcularlas (Grayson 1984; Lyman 1994; Grayson y Frey 2004).
Por otra parte, los aspectos culturales, tafonómicos y metodológicos que deben haber influido en la representación de los taxa de los conjuntos se desdibujan a la hora de hacer análisis de grano grueso. Lo que cabe mencionar es que, en mayor o en menor grado, en los distintos sitios incluidos en este trabajo se han realizado análisis tafonómicos por los equipos de trabajo de cada uno de ellos. En aquellos sitios en los que se encuentran realizados en mayor detalle los análisis tafonómicos, los resultados reflejan que no hay agentes que afecten de manera intensa el registro arqueofaunístico (Cornaglia 2009; Pérez Jimeno 2007; Santiago 2012; Sartori 2013; Sartori y Pérez Jimeno 2012; Sartori y Colasurdo 2011).
Por otro lado, en lo que refiere a los posibles sesgos generados por diferentes métodos de recuperación, cabe señalar que las técnicas empleadas son prácticamente uniformes, ya que, en todos los casos, los sitios se excavaron por unidades artificiales, tridimensionando los materiales y utilizando zaranda de mayas finas (0,5 cm a muy finas, 0,1 cm). Esto ha permitido la recuperación de elementos muy pequeños, correspondientes generalmente a peces o roedores. Por lo expuesto, no se considera que haya fuentes de variabilidad tan significativas como para que no se pueda realizar el presente análisis.
Resultados
De los trece conjuntos analizados, cinco (sitios CA, BP, CBin, PM1, LL, de la tabla 1 y figura 1) corresponden a sitios emplazados en el sector CHH; cuatro, al sector ESP (sitios RSC, FP, LT, PB); dos, a PDP (BG, PMan), y dos, a la PAM (sitio LED con dos conjuntos). Los taxa representados se distribuyen en un total de diecinueve especies -considerando a los peces como macrotaxón-, que fueron agrupadas en nueve categorías (ver la tabla 3). Estas han sido establecidas en función de que, en la mayoría de los casos, su presencia en el registro se debe a agentes antrópicos.
Tabla 3 Datos zooarqueológicos recalculados por ecorregión. NISP total, NISP por taxones analizados e índices de abundancia recalculados

Fuente: los autores.
Todos los conjuntos sumaron un total de 56.472 restos faunísticos, pertenecientes al macrotaxón vertebrados; encontrándose constituido el conjunto más pequeño por 179 restos, y el mayor, por 23.885. Considerando el total de los conjuntos desde categorías taxonómicas amplias, se observa que los peces constituyen, en promedio para toda las ecorregiones, el 49,5%, mientras que los mamíferos representan el 47,5%, y las aves y los reptiles sumados, el 3% restante. Cabe señalar que las cuatro primeras categorías de la tabla 3 (Peces, Roedores, Mammalia Indeterminados y Cérvidos) representan el 88% de los restos faunísticos de todas las ecorregiones de la provincia de Santa Fe.
Si se observa la composición de cada sector, tenemos que en la zona del CHH, los cinco sitios presentan un total de 34.354 especímenes, con una media de 6.870, mientras que la mediana es de 2.962, y es la que exhibe mayor robustez en sus muestras (tabla 3). Esta zona es la que refleja una mayor abundancia de Cérvidos, Roedores, Aves y Reptiles. El sector ESP presenta NISP más bajos en sus registros, para un total de 7.056, una media de 1.764 y una mediana de 1.716. La zona PDP presenta en sus dos conjuntos un total de 3.742, y por último, la zona PAM presenta un total de 11.320, concentrados en un solo sitio con un conjunto de estratigrafía y otro de superficie.
Describiendo cada uno de los recursos, los peces están presentes en once sitios, y en su conjunto constituyen el recurso mejor representado en términos de NISP (N=28228). Cabe señalar que en el sitio ubicado en el sector pampeano, estos se encuentran ausentes. Si nos detenemos en su abundancia a lo largo de la planicie de inundación del Paraná, pueden observarse algunas diferencias. En tal sentido, las representaciones más robustas se observan en sentido decreciente Sur (PDP)-Norte (CHH) (ver la figura 3). Este aspecto concuerda con las proporciones en la presencia de los otros dos recursos que habrían sido centrales para la subsistencia: los roedores y los cérvidos, en esta zona. Es decir que se observa una tendencia inversamente proporcional entre los peces y los demás recursos presentes. Si se atiende a cada ambiente específico, el caso del CHH representa el 77% (N=21985) de todos los restos asignados a peces; siendo destacable el sitio Cerro Aguará, con un NISP de 20.094. Además, en la zona de la planicie de inundación del Paraná (tanto en el CHH como en el ESP), los peces cobran gran importancia, superando en los sitios insulares el 50% de los NISP de los conjuntos.
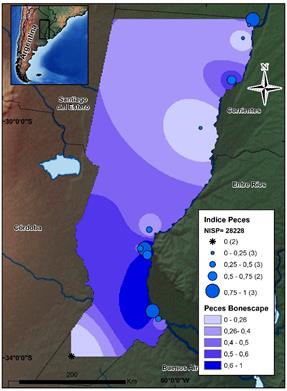
Fuente: los autores.
Figura 3 Mapa de distribución de los valores por sitio del índice de abundancia de peces. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
En el sector del ESP se produce un incremento de la abundancia de los peces (IA=0,67), y, además, hacia el interior de esta zona fue detectada una diferencia entre la composición de los conjuntos insulares vs. aquellos que corresponden a los sitios localizados en la zona continental. Tal variación refiere al predominio de los mamíferos en la zona continental, mientras que en el sector insular el predominio es de los peces. Este patrón se observa también en el sector CHH, en el que, sin embargo, la disminución de los peces (IA=0,64) va acompañada de un notable incremento en los cérvidos, roedores y aves. Más allá de esto, es precisamente en el CHH donde se encuentra el sitio Cerro Aguará, en el cual se ha registrado la mayor cantidad de especies de peces identificadas (N=23) (Musali, Feuillet y Sartori 2013), mientras que en otras zonas de la planicie de inundación se registran en todos los conjuntos entre cinco y once taxa pertenecientes al macrotaxón peces.
Los roedores (N=8836) constituyen, conjuntamente con los peces, el otro recurso ampliamente representado en toda la provincia (ver la figura 4). Sus restos aportan el 15,5% dentro del NISP general para todas las ecorregiones, y si se tiene en cuenta que los mamíferos identificados a nivel de orden-familia en su conjunto constituyen sólo el 29,2% del NISP, puede observarse la gran importancia de estos recursos en la dieta de los cazadores-recolectores. La distribución espacial de los roedores como categoría amplia abarca todas las ecorregiones, hallándose presentes en el 100% de los sitios, aunque un notable incremento en su abundancia se observa en los sectores PDP (IA=0,27) y CHH (IA=0,19). En el sector pampeano se registran los NISP más bajos para esta Familia, presentando los índices de abundancia un valor de 0,06, ya que el recurso preponderante es el guanaco.
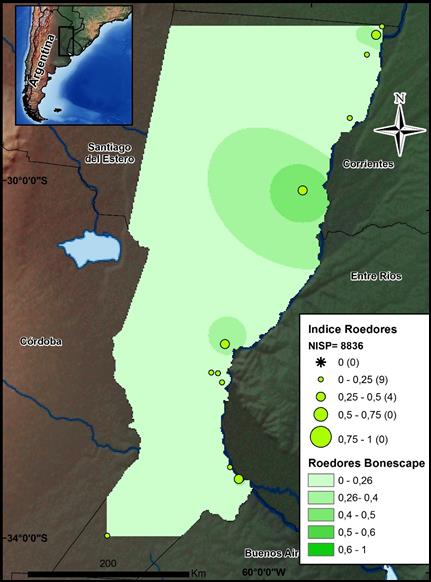
Fuente: los autores.
Figura 4 Mapa de distribución de los valores por sitio del índice de abundancia de Rodentia. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
Los cérvidos, como categoría amplia, se encuentran en todos los sitios de los cuatro sectores, pero, a diferencia de los otros dos recursos más abundantes, presentan una acentuada disminución en términos de NISP (N=3412). Dentro del total de especímenes, esta categoría representa sólo el 3%, lo cual refleja valores sustancialmente más bajos que los de peces y roedores. El incremento en el índice de abundancia de los cérvidos se produce en sentido Sur-Norte (ver la figura 5), presentando los registros de la zona CHH el 70% de los elementos identificados para alguna de las tres especies que se registran en dicha zona. La representación aumenta crecientemente en los conjuntos del CHH, mientras que disminuye hacia el sur en el ESP, PDP, y sobre todo en la PAM.
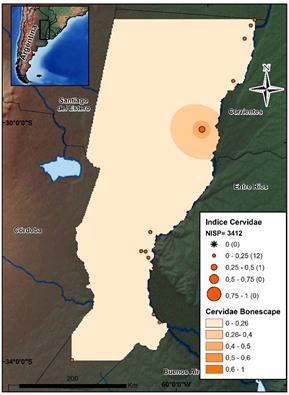
Fuente: los autores.
Figura 5 Mapa de distribución de los valores por sitio del índice de abundancia de Cervidae. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
En cuanto a Lama sp., este taxa se encuentra presente en sólo 33% de los sitios, y el NISP es sustancialmente diferente entre los sitios del CHH, ESP y PDP -que en total suman 78 restos-, mientras que en PAM se encuentra representado por 3.245 especímenes (ver la figura 5). Entre los demás mamíferos que presentan bajas frecuencias se encuentran los dasipódidos (N=850), los cuales se hallan representados en depósitos de los tres sectores, aunque su abundancia refleja un patrón espacial diferencial para los conjuntos de los sectores CHH, ESP, PDP y PAM. Los dasipódidos están presentes en un 33% de los conjuntos y se distribuyen en cuatro especies identificadas. Su presencia es mayor en los sitios ubicados en los sectores PAM (IA=0,05) y PDP (IA=0,03), mientras que en el CHH y ESP, los IA son menores a 0,009 (ver la figura 6). Para el caso de los carnívoros (N=158), si bien se encuentran presentes en seis sitios, el 88% de los restos pertenecen al sector PAM. Cabe señalar que, entre las diferentes especies que se ven representadas, no todas poseen indicios de haber sido aprovechadas, mientras que en el caso de los dasipódidos, su presencia suele estar asociada a su aprovechamiento humano.
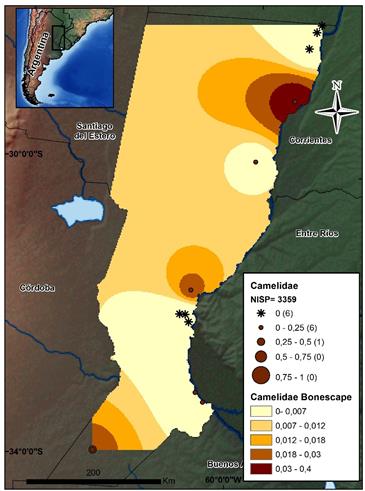
Fuente: los autores.
Figura 6 Mapa de distribución de los valores por sitio del índice de abundancia de Camelidae. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
Las aves (N=1113), al igual que los roedores y los cérvidos, presentan mayor abundancia en el sector CHH, decreciendo en sentido Norte-Sur, aunque se observa un incremento en el sector suroeste (ver la figura 7). Como macrotaxón, su presencia se registra en un 41% de los sitios, y no han sido identificadas a niveles específicos en la mayoría de los casos -a excepción de Rhea americana-, debido a la amplia variabilidad específica de toda la zona y la falta de colecciones de referencia.
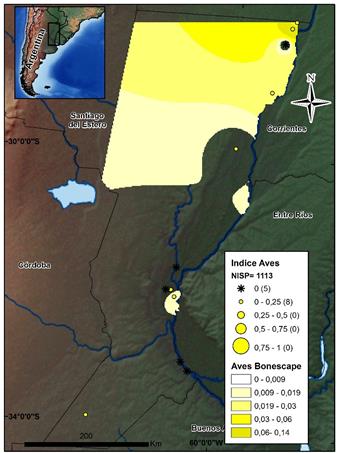
Fuente: los autores.
Figura 7 Mapa de distribución de los valores por sitio del índice de abundancia de Aves. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
Por último, los reptiles (N=632) presentan una mayor abundancia en el sector CHH (IA=0,018), decreciendo significativamente sus IA en sentido Norte-Sur (ver la figura 8). Este hecho se ve reflejado también en que la zona del sector CHH posee el 84% de los restos asignados a esta Clase.
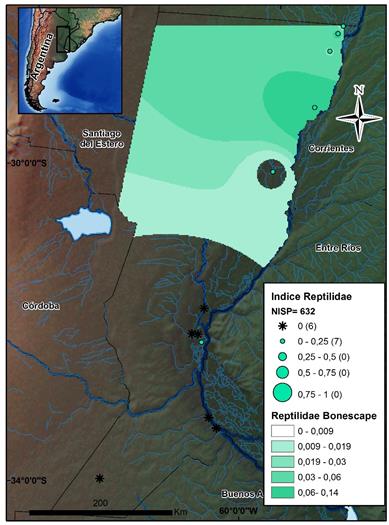
Fuente: los autores.
Figura 8 Mapa de distribución de los valores por sitio del índice de abundancia de Reptilidae. Se indican el NISP total de este conjunto y el Bonescape generado para este taxón
El sector que posee mayor cantidad de NTAXA, en promedio (sin considerar la variabilidad del macrotaxón peces), es el CHH de la planicie de inundación, con un 11,1; en tanto que el sector ESP (con 7,8) y el PAM (5)2 presentan valores más bajos. El cálculo de la moda -realizada considerando los valores por sitio y por zona-, que refleja la mayor recurrencia en los valores de riqueza específica para cada sector, es de 9 para CHH, de 8 para el ESP, mientras que el PAM sólo se compone de una muestra. Estos valores son coincidentes con la variabilidad faunística que es intrínseca a cada una de las ecorregiones de la provincia de Santa Fe, en donde el Chaco Húmedo se destaca por su mayor riqueza.
Discusión
Considerando que la provincia de Santa Fe posee una riqueza en la biodiversidad que se incrementa en sentido Sur-Norte y Oeste-Este, las características ambientales que posee cada una de las zonas en las cuales se emplazan los sitios tornarían esperable que haya diferencias en la preponderancia de los recursos explotados en cada sector. Es decir que, más allá de las variaciones en la composición de cada conjunto en particular, la expectativa es que se observen tendencias en cada sector, que irían de la mano de la oferta faunística. Se consideró que para realizar un análisis en términos espaciales, el GIS constituye una herramienta versátil que hace posible visualizar tendencias en cada una de las zonas analizadas, permitiendo compararlas entre sí. Los resultados expresados en el apartado anterior posibilitaron observar recurrencias en cuanto a las categorías taxonómicas amplias que dominan los conjuntos en cada ecorregión. Si bien esto es sumamente útil para la discusión de los recursos que habrían tenido un rol central en la subsistencia de los grupos cazadores-recolectores de la región, debe hacerse una consideración al respecto. Dado que las categorías de análisis engloban diferentes taxa, un análisis de grano más fino hacia el interior de cada ecorregión se torna necesario, ya que existen variaciones que reflejan la explotación puntual de ciertas especies, en detrimento de otras. Tal situación se encuentra estrechamente vinculada con la estructura del ambiente y con los requerimientos ecológicos que poseen las distintas especies que constituyeron la base principal de la subsistencia.
En lo que respecta a los peces, en sitios del CHH, ESP y PDP, los bagres (Pimelodella laticeps, Pimelodus maculatus y Pimelodus albicans) y otras de las especies identificadas en los diferentes conjuntos (como vieja del agua, cascarudo y torito) tienen mayor presencia dentro de los NISP% de las muestras (Musali, Feuillet y Sartori 2013; Sartori 2013). Si se tiene en cuenta el hábitat de tales especies, puede deducirse que estas habrían sido capturadas en aguas poco profundas y vegetadas. Además, algunas de ellas suelen habitar ambientes lénticos y lóticos, lo cual coincide con las características del emplazamiento de los distintos sitios insulares (Musali, Feuillet y Sartori 2013; Sartori 2013). Cabe destacar que en la PAM, los peces se encuentran totalmente ausentes, y al respecto es interesante que en las prospecciones realizadas por Ceruti en sistemas lagunares del sudoeste santafesino tampoco se registran restos de este macrotaxón, pese a que todos los sitios se encuentran todos cercanos a cursos de agua o lagunas (Ceruti 1987; Cornaglia 2013; 2014).
Los roedores son el segundo recurso más explotado en términos de NISP y se hallan en todos los sitios, aunque cabe señalar que entre las especies potencialmente consumibles, y con evidencia de haberlo sido, se registran diferencias. El coipo presenta la distribución más homogénea, encontrándose en casi todos los conjuntos, a excepción de PAM. En los sitios localizados en el CHH, ESP y PDP se halla representado por una amplia cantidad de restos (N=4186), mientras que para el carpincho, si bien se halla en un 75% de los conjuntos, los NISP son sustancialmente más bajos (N=496), lo cual refleja una baja explotación de este roedor, que supera ampliamente en porte al coipo. Su bajo NISP y su menor presencia en los sitios han llevado a considerar alguna restricción en su aprovechamiento, sobre todo para la zona del Paraná inferior y Delta (Acosta 2005; Feuillet 2002). No obstante, para la zona media, Sartori (2013) propone que las variaciones en su representación podrían estar dadas en términos espaciales, ya que este animal se encuentra mejor representado en los sitios insulares, decreciendo su abundancia en la zona continental. En tanto que en la zona más al norte, Pérez Jimeno (2007) y Santiago (2002) concluyen que este gran roedor constituiría parte de la dieta de los grupos cazadores-recolectores-pescadores. Este hecho se ve avalado también con los datos de crónicas y estudios etnográficos entre grupos indígenas del Gran Chaco que registran el consumo de esta especie. Cabe señalar que, a diferencia de lo que ocurre en los sectores CHH, ESP y PDP, en la PAM la oferta de roedores potencialmente consumibles es más baja, y esto se evidencia también en la baja explotación de estos taxa en sitios que están alejados de la influencia de la ecorregión del Delta e Islas del Paraná.
En cuanto a las tres especies de cérvidos identificadas, en todos los conjuntos se registra alguna de ellas, aunque existen diferencias entre las especies más abundantes. La especie de mayor porte -ciervo de los pantanos (Blastocerus dichotomus)- es la que predomina en todo el centro-este santafesino (hallándose presente en un 75% de los sitios), mientras que en los sitios que se encuentran hacia el interior y en PAM, esta se halla ausente. Le sigue en abundancia el venado de las pampas (Ozotoceros bezoarticus, N=788), que es la segunda especie de mayor tamaño, la cual posee una menor representación, hallándose presente en un 66% de los conjuntos. Por último, Mazama gouazoubira es el cérvido más pequeño, y sólo se halla representada en un 16% de los conjuntos analizados, siendo estos un sitio del noreste y otro del sureste de la provincia de Santa Fe. Probablemente la preponderancia del ciervo de los pantanos responda a cuestiones espaciales que se vinculan con los requerimientos ecológicos de esta especie y de las otras dos que habitan la región en estudio. Es de destacar que no se encuentren presentes restos de ciervo en sectores alejados de zonas anegadas, lo que refleja que en los sectores pampeanos, los grupos humanos estarían explotando recursos locales (como Lama sp. y dasipódidos). Esta tendencia (que debe seguir siendo evaluada a medida que se estudien nuevos contextos pampeanos en la provincia) contrasta con lo que se observa en los sitios del CHH, ESP y PDP emplazados en zonas continentales ecotonales, en donde se encuentran presentes especies adaptadas a diferentes ambientes (tanto acuáticos como secos-abiertos). En cambio, Ozotoceros bezoarticus es un ciervo típico de ambientes llanos, abiertos, sin vegetación arbórea, con lo cual su presencia en algunos conjuntos reflejaría vínculos con la región Pampeana, próxima a la planicie de inundación del Paraná. En cambio, la presencia del ciervo de los pantanos refleja rangos de acción más acotados y vinculados al sector insular.
Los camélidos, al igual que la presencia de O. bezoarticus, serían otros de los animales que reflejan contacto con zonas más secas y no anegadas. Al respecto es interesante señalar que las dos especies mencionadas se encuentran siempre asociadas: en todos los casos en que se presenta Lama sp. -que es la que más baja frecuencia presenta- se halla también el venado. Este recurso sería el preponderante en el sitio localizado en la región PAM. Por su parte, Rhea americana sólo se encuentra en un sitio al norte y en la PAM, en donde tiene una representación del NISP% que refleja cierta importancia económica.
La preponderancia de los recursos en términos espaciales ha servido para establecer tendencias que se observan en sentido Norte-Sur, pero también Este-Oeste. En este último caso (si se tiene como referencia el NISP% de los peces, que mantienen una relación inversamente proporcional con los mamíferos -que son la otra clase más representada-) se evidencia que los taxa más representados en espacios insulares son los peces, siguiéndoles en abundancia los roedores y luego los ciervos (y exclusivamente, el ciervo de los pantanos), en los sitios del CHH, ESP y PDP. En este sentido, en los sitos emplazados en los espacios continentales, en cambio, los peces no superan el 50% del NISP de los conjuntos, viéndose más representados los mamíferos, entre cuyas especies se hallan aquellas típicas de ambientes más abiertos y secos. Una mayor diferencia se observa en la PAM, en donde los peces se encuentran ausentes, y Lama sp. posee gran importancia en términos de NISP%.
En síntesis, puede considerarse que en el CHH, ESP y PDP, los peces, los roedores y el ciervo de los pantanos habrían tenido un rol preponderante para la dieta-subsistencia de los grupos humanos de la zona. Esto refleja circuitos de explotación vinculados al ambiente ribereño, en tanto que en ciertos sectores (los sitios emplazados en el sector continental) se registra el contacto con recursos propios de la región pampeana. No obstante esto, en la mayoría de los sitios, cuando los peces disminuyen, aumentan los roedores, los cérvidos, y en algunos casos -en la zona norte, fundamentalmente- se produce una mayor diversidad entre los taxa, registrándose, por ejemplo, un incremento en los reptiles y aves.
Conclusiones
Este trabajo realiza la síntesis de la información arqueofaunística disponible para las distintas ecorregiones de la provincia de Santa Fe. Los estudios en esta zona se han incrementado en los últimos quince años, aunque se observa una desigual producción de información, ya que esta se centra casi exclusivamente en ambientes cercanos al río Paraná. Esto refleja la necesidad de continuar investigando los diferentes espacios de la región para comprender el particular uso del espacio y los recursos que se habrían realizado en este sector de Argentina durante el Holoceno tardío. Sólo a medida que se avance en el conocimiento a nivel de sitio podrán realizarse trabajos integradores que se basen en la información de grano fino ya generada.
En toda la zona de la cuenca del Paraná hay una gran cantidad de especies disponibles, sin embargo, la mayor cantidad recae en los peces y en dos especies de roedores, ciervos y Lama sp. No obstante, hay variabilidad en la preponderancia de cada recurso, tanto en sentido Norte-Sur como Este-Oeste. Es decir que la explotación faunística va de la mano de la oferta en cada ecorregión, demostrando que es útil analizar la importancia de los recursos en función de su distribución espacial. Además se observa una especialización recurrente para el CHH, ESP y PDP, aunque si se mira con detenimiento cada uno de los sectores, se presentan casos en donde las aves, los reptiles o los dasipódidos habrían tenido cierta importancia económica.
La implementación de GIS a escala regional posibilitó evaluar la variabilidad en la explotación faunística en la provincia de Santa Fe, la cual habría estado basada -en términos generales- en las mismas especies en los sectores norte y sur de la provincia de Santa Fe, mientras que en sentido Este-Oeste se produce una variación en la mayor presencia y dominancia de especies pampeanas.

