










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCES Medicina Veterinaria y Zootecnia
On-line version ISSN 1900-9607
Ces. Med. Vet. Zootec. vol.8 no.2 Medellín July/Dec. 2013
Producción de metano in vitro de dos gramíneas tropicales solas y mezcladas con Leucaena leucocephala o Gliricidia sepium
In vitro methane production from two tropical grasses alone or in combination with Leucaena leucocephala or Gliricidia sepium*
Produção in vitro de metano a partir duas gramíneas tropicais avaliadas individualmente e associadas com Leucaena leucocephala ou Gliciridia sepium
Isabel Cristina Molina Botero1*, Zoot, (c) MSc; Juan Manuel Cantet2, MV, (c) PhD; Sebastián Montoya3, (c) Zoot; Guillermo Antonio Correa Londoño4, IF, MSc, PhD; Rolando Barahona Rosales5, BSc, MSc, PhD.
*Autor para la correspondencia: Isabel Cristina Molina, Bloque 50-321, Universidad Nacional de Colombia, Medellín, Antioquia. E-mail: icmolina@unal.edu.co.
1 Estudiante de Maestría en Ciencias Agrarias, Línea de Nutrición Animal, Departamento de Producción Animal, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Sede Medellín.
2 Estudiante de Doctorado en Ciencias Agropecuarias, Departamento de Producción Animal, Facultad de Agronomía, Universidad de Buenos Aires, Argentina.
3 Zootecnista, Joven investigador, Universidad Nacional de Colombia, Sede Medellín.
4 Profesor Asociado, Departamento de Ingeniería Agronómica, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Sede Medellín.
5 Profesor Titular, Departamento de Producción Animal, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Sede Medellín.
(Recibido: 10 de agosto, 2013; aceptado: 2 de noviembre, 2013)
*Para citar este artículo: Molina Botero IC, Cantet JM, Montoya Sebastián, Correa Londoño GA, Barahona Rosales R. Producción de metano in vitro de dos gramíneas tropicales solas y mezcladas con Leucaena leucocephala o Gliricidia sepium. Rev CES Med Zootec. 2013; Vol 8 (2): 15-31.
Resumen
La percepción del cambio climático como uno de los grandes problemas ambientales del siglo XXI viene creciendo en las últimas décadas. La emisión de metano por fermentación ruminal es una pérdida de energía potencialmente utilizable. El objetivo fue estimar las emisiones de metano de forrajes usados en dietas para alimentar bovinos. Las dietas evaluadas provenían de un sistema silvopastoril intensivo (SSPi) con leucaena (Leucaena leucocephala; Leu) y pasto guinea (Megathyrsus maximus; Gui) y de confinamiento con matarratón (Gliricidia sepium; Mat) y pasto ángleton (Dichantium aristatum; Ang). Muestras representativas de los forrajes fueron analizadas por su contenido de nutrientes; las emisiones de metano de los forrajes solos y sus mezclas (90:10, 80:20 y 70:30, teniendo mayor participación las gramíneas) fueron medidas in vitro, mediante la técnica de producción de gas. La degradación de la materia seca se midió a las 24, 48, 72 y 96 h. Las leguminosas tuvieron mayores contenidos de proteína y grasa bruta; mientras que las gramíneas, mayores contenidos de fibra insoluble en detergente neutro y ácido y cenizas. La mayor producción de gas a las 96 h fue para Gui70-Leu30 (156 ml) y la menor fue Leu100 (P≤0,05; 121 ml). Para el sistema en confinamiento, la acumulación de gas de Ang70-Mat30 y Ang80-Mat20 a partir de a las 48 horas fueron superiores a los demás tratamientos (P<0,05). Las leguminosas mostraron mayores tasas de degradación que las gramíneas (P≤0,05). En ambos sistemas de producción no se encontraron diferencias significativas en la pérdida de energía digerida en forma de metano.
Palabras clave: confinamiento, ganadería, gases con efecto invernadero, silvopastoreo.
Abstract
Climate change perception in recent decades has been growing as one of the major environmental issues of the XXI century. Methane emissions by rumen fermentation represent a loss of potentially usable energy. The objective of this study was to estimate methane emissions from various forages used in beef cattle diets. The diets evaluated corresponded to an intensive silvopastoral system (SSPi) with Leucaena (Leucaena leucocephala, Leu) and Guinea grass (Megathyrsus maximus; Gui) and a confinement system with Matarratón (Gliricidia sepium; Mat) and Angleton grass (Dichanthium aristatum; Ang). Representative forage samples were analyzed for nutrient contents. Methane emissions of individual forages and forage mixtures (90:10, 80:20, and 70:30, for grass:forage, respectively) were measured in vitro using the gas production technique. Dry matter degradation was measured at 24, 48, 72, and 96 h. Legumes had higher protein and crude fat content than grasses, while grasses had higher neutral detergent fiber, acid detergent fiber, and ash. The mixture composed by Gui70 - Leu30 had the highest gas production at 96 h (156 ml), while the lowest corresponded to Leu100 (P≤0.05; 121 ml). Regarding the confinement system results, gas accumulation after 48 hours by Ang70 - Mat30, and Ang80 - Mat20 were higher than the other treatments (P<0.05). Legumes had higher degradation rates than grasses (P≤0.05). No significant differences were found for methane losses between both production systems.
Key words confinement, greenhouse gases, livestock, silvopasture.
Resumo
A percepção da mudança climática como um dos grandes problemas ambientais do seculo XXI está aumentando nas últimas décadas. A emisão de metano por fermentação ruminal é uma perda de energia potencialmente utilizavel. O objetivo foi estimar as emisões de metano de forragens utilizados em dietas para alimentar bovinos. As dietas avaliadas vieram de um sistema silvopastoril intensivo (SSPi) com leucaena (Leucaena leucocephala; Leu) e capim-mombaça (Megathyrsus maximus; Gui); e de confinamento com gliricídia (Gliricidia sepium; Mat) e a pastagem ángleton (Dichantium aristatum; Ang). Amostras representativas das forragens foram analisadas por seu conteúdo de nutrientes; as emissões de metano das forragens semeadas em monocultivo e misturadas (90:10, 80:20 e 70:30, tendo maior participação às gramíneas) foram medidas in vitro, mediante a técnica de produção de gás. A degradação da matéria seca estimou-se ás 24, 48, 72 e 96 h. As leguminosas tiveram maiores conteúdos de proteína e gordura bruta; enquanto as gramíneas tiveram maior fibra insolúvel em detergente neutro e azedo e cinzas. A maior produção de gás ás 96 h foi para Gui70-Leu30 (156 ml) e a menor foi Leu100 (P≤0,05; 121 ml). Para o sistema em confinamento, a acumulação de gás de Ang70-Mat30 e Ang80-Mat20 a partir das 48 horas foram superiores aos demais tratamentos (P<0,05). As leguminosas demonstraram maiores taxas de degradação que as gramíneas (P≤0,05). Nos dois sistemas de produção não se encontraram diferenças significativas na perda de energia digerida em forma de metano.
Palavras chave: confinamento, gases com efeito estufa, produção de gado, silvopastoreio.
Introducción
La ganadería es una actividad de gran importancia dentro del sector agropecuario colombiano y casi 35% de la superficie del país es dedicada a esta actividad(13). Sin embargo, los índices productivos de este sector, especialmente los de la cadena cárnica bovina, son bajos. En respuesta a la globalización de mercados y la creciente presión sobre el uso del suelo, la ganadería bovina colombiana debe emprender una serie de cambios tecnológicos y científicos que le permitan continuar contribuyendo con el bienestar social, sin detrimento del medio ambiente.
Esto cobra gran importancia en lo que se refiere al cambio climático, sin duda uno de los problemas ambientales predominantes en el siglo XXI, que ha venido creciendo en las últimas décadas(52). Un informe de la FAO(14) sobre la ganadería señala que esta es la principal actividad antropogénica del uso de la tierra y que genera el 18% de las emisiones de gases de efecto invernadero, siendo responsable del 9% de las emisiones de dioxido de carbono (CO2), principalmente por deforestación, el 37% de las emisiones de metano (CH4), en gran parte por la digestión de los rumiantes y el 65% del óxido nitroso, generado en gran medida por el estiércol. En el 2008, utilizando la metodología Tier II del IPCC, se estimó que la ganadería vacuna colombiana, que en el 2004 contaba con cerca de 25 millones de cabezas de ganado, emitió un total de 2578 Gg de CH4 al año, el 56,9% de las cuales correspondía a emisiones de ganado bovino no lechero(19).
A partir del desarrollo del retículo y el rumen un par de semanas despues de su nacimiento, el ganado bovino inicia su producción de metano(23). En este proceso digestivo, que ocurre bajo condiciones anaeróbicas, participan microorganismos del dominio Archaea, pertenecientes al reino Eucariota(22, 27), los cuales pueden estar en concentraciones entre 107 y 109 células por ml del líquido ruminal(25). Los dos géneros más populares son Methanobrevibacter y Methanosarcina(27, 33). Los microorganismos metanogénicos utilizan diferentes sustratos tales como el acetato, grupos metilos, dihidrógeno y CO2(26, 33), productos finales de la fermentación de proteínas, almidones y compuestos de la pared celular por parte de microorganismos como protozoos, hongos y bacterias(29, 33).
La tasa de emisión de metano por fermentación ruminal, está relacionada con las características físico-químicas de la dieta (e.g. la digestibilidad de la materia seca, DMS y la concentración de fibra insoluble en detergente neutro, FDN), las cuales afectan directamente el nivel de consumo y la frecuencia de alimentación(49). Los rumiantes que producen más metano son habitualmente los que consumen praderas de trópico bajo, ya que son de regular a baja calidad (alto contenido de lignocelulosa y bajos tenores de otros nutrientes, principalmente proteína y carbohidratos solubles2), lo contrario sucede cuando se incluye leguminosas en la dieta(8,43), por tener contenidos altos de proteína, bajos de FDN y presencia de compuestos secundarios como los taninos(7, 47, 45).
Se ha estimado que la energía consumida que se elimina en forma de metano varía entre el 2 y 15%(23). Según Calsamiglia et al(9), esto reduce la eficiencia energética del animal, ya que estos gases se pierden a través del eructo y no van hacia destinos productivos como la formación de ácidos grasos volátiles (AGV), esto lo corroboran Murray et al(37), quienes afirmaron que alrededor del 89% del metano entérico producido es eliminado por la boca y la nariz.
En la búsqueda de mayor eficiencia y productividad, en Colombia se han promovido nuevos sistemas de producción, como los sistemas silvopastoriles intensivos (SSPi)(36) y la producción en confinamiento, que hacen un uso más intensivo de los recursos y aunque con pocas evaluaciones concretas de su impacto ambiental(11), representan una alternativa para disminuir las emisiones de metano, ya que es posible incluir en sus dietas leguminosas. El presente estudio tiene como propósito determinar el efecto sobre las emisiones de metano de una dieta típica de un SSPi y de una dieta basada en gramíneas y leguminosas que podría ser de gran utilidad para un sistema de ceba en confinamiento.
Materiales y Métodos
Forrajes
Los forrajes fueron recolectados en la Estación Agraria Cotové, propiedad de la Universidad Nacional de Colombia, sede Medellín, ubicado en el municipio de Santafé de Antioquia, vereda "El Espinal", distante 74 kilómetros de Medellín. Su clasificación ecológica según Holdridge es bs-T, siendo sus suelos provenientes de calizas y arcillas calcáreas. El centro está a 540 metros sobre el nivel del mar y los promedios de temperatura y precipitación son 27ºC y 1.100 mm, respectivamente. Las muestras representativas del SSPi: la leguminosa leucaena (Leucaena leucocephala) y la gramínea guinea (Megathyrsus maximus), fueron obtenidas de un SSPi de cinco ha en el que se encontraban pastoreando veinte novillos brahman de 360 kg de peso vivo. La leguminosa matarratón (Gliricidia sepium) y la gramínea ángleton (Dichantium aristatum) fueron obtenidas de la dieta ofrecida a otros veinte novillos de 330 kg de peso vivo que estaban en confinamiento. Cada uno de los forrajes tenía una edad promedio de rebrote de 50 a 60 días y su cosecha fue realizada durante la época seca.
Análisis bromatológicos
Después de su recolección, los forrajes fueron llevados al Laboratorio de Bromatología de la Universidad Nacional de Colombia sede Medellín, donde se secaron en estufa de ventilación forzada a una temperatura de 65ºC por 72 h y se molieron a través de una criba de un 1 mm, utilizando un molino Romer R.A.S (Romer Labs, México).
Cada uno de los materiales fue analizado por su contenido de materia seca (MS, en estufa de aire forzado a 105°C hasta alcanzar peso constante, basado en ISO 649621), proteína cruda (PC, por el método de Kjeldahl, según NTC 465716), fibra insoluble en detergente neutro y ácido (FDN y FDA, respectivamente, según técnica secuencial descrita por Van Soest et al50), lignina (Lig, según Van Soest et al51) y extracto etéreo (EE, por extracción Soxhelet por inmersión; NTC 66815). El contenido de cenizas (CEN) se determinó por incineración directa en una mufla a 500°C, según AOAC 942.05(4). El contenido de los minerales: Calcio (Ca) y Fosforo (P) se determinó por Espectrofotometría AA y U.V- VIS (basado en NTC 5151 y 498118, 17), respectivamente. Finalmente, se determinó el contenido de energía bruta por calorimetría, basado en ISO 9831(20).
Cinética de producción de gases
La técnica de gases se realizó de acuerdo a lo sugerido por Theodorou et al(48). Se pesaron 300 mg de cada uno de los cuatros forrajes, tanto solos como en mezclas representativas de cada sistema: SSPi (guinea y leucaena) y confinamiento (ángleton y matarratón) en las siguientes proporciones: 90:10, 80:20 y 70:30, donde las mayores proporciones correspondieron a las gramíneas y el resto a las leguminosas. Así, se generaron un total de 10 tratamientos, cinco por cada sistema de producción.
El líquido ruminal fue obtenido de tres vacas raza brahman, las cuales estaban recibiendo una dieta a base de pasto estrella (Cynodon plectostachys) de aproximadamente 45 días de crecimiento más sal mineralizada. La colecta del líquido ruminal se realizó en la Central Ganadera de Medellín, ubicada en la Autopista Norte en la ciudad de Medellín (altitud de 1536 msnm, temperatura entre 16 y 28°C y 1656 mm de precipitación promedio anual). La digesta fue retirada del rumen inmediatamente después de que este fuera separado de los animales durante el proceso de faenado. Rápidamente se exprimió la digesta a fin de extraer el líquido ruminal y este fue almacenado en termos previamente calentados con agua a 39°C para su transporte al laboratorio GRICA, propiedad de la Universidad de Antioquia, que se encuentra ubicado en la ciudad de Medellín a 15 minutos de la Central Ganadera. Una vez en el laboratorio, el líquido ruminal se filtró nuevamente a través de paños de algodón y fue transferido a un erlenmeyer sumergido en un baño maría a 39°C, con gaseo constante de CO2. Los tres inóculos ruminales utilizados fueron constituidos por 2 animales diferentes cada uno (i.e. inóculo= repetición estadística).
La incubación se realizó en botellas de vidrio de 100 ml. A cada botella se le adicionó el sustrato correspondiente y con una jeringa graduada, se le agregaron 45 ml del medio de cultivo y 5 ml de líquido ruminal (i.e. relación licor: medio, 1: 9). Por cada 10 botellas con sustrato, medio e inóculo se contaba con dos botellas que no contenía sustratos. Posteriormente, las botellas fueron selladas en forma hermética con tapas de caucho, agitados manualmente y transferidos a una estufa de ventilación forzada a 39ºC.
Las lecturas de presión (mmHg) se realizaron en 11 horarios (i.e. 2, 4, 6, 9, 10, 12, 15, 24, 48, 72 y 96 h pos-inoculación), con la ayuda de un transductor de presión (Ashcroft Inc, USA) conectado a un lector digital y a una válvula de tres salidas. La primera salida fue conectada a una aguja hipodérmica de 10 mm que se insertaba en el interior de los frascos incubados, la segunda al transductor de presión y la tercera a una jeringa plástica que sirvió para la medición del volumen. El volumen de gas colectado fue acumulado en botellas de 60 ml. Una vez las lecturas fueron llevadas a cabo, los frascos fueron agitados manualmente y llevados de nuevo a la estufa.
Los datos de producción de gas fueron ajustados al modelo no lineal de Gompertz modificado, que tiene la siguiente ecuación:
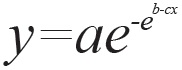
Donde, y es igual a la producción acumulada de gas a un tiempo x, a > 0 es la producción máxima de gas, el parámetro b > 0 es la diferencia entre el gas inicial y el gas final a un tiempo x y el parámetro c > 0 describe la tasa específica de acumulación de gas.
La aplicación práctica de este modelo requiere la conversión de los parámetros a, b, c en parámetros con significado biológico. Para los fines de este estudio, ellos son: hora al punto de inflexión (HPI, horas), gas al punto de inflexión (GPI, ml), tasa máxima de producción de gas (TMPG, ml/h) y fase Lag (FL o establecimiento microbiano, h). Para su estimación se usan las siguientes fórmulas: HPI = b/c; GPI = a/e; TMPG = (a*c)/e; FL= ((b/c)-(1/c)); donde "e" es el número de Euler, equivalente a ≈ 2,718281828459.
Degradación de la materia seca
Para medir la degradación de la materia seca y la materia orgánica, el contenido de las botellas retiradas de la fermentación a diferentes tiempos (i.e. 24, 48, 72 y 96 h) fue filtrado con la ayuda de un embudo de plástico, una bomba de vacío y papel filtro con un tamaño de poro de 0,5 mm. Los filtros fueron luego secados en estufa de aire forzado a 65ºC durante 48 horas y se pesaron, utilizando una balanza de precisión.
Determinación de la concentración de metano
La concentración de metano se determinó usando cromatografía de gases en muestras del gas acumulado durante las primeras 48 h de fermentación, las cuales fueron recolectadas en botellas de 60 ml. La cuantificación de metano fue realizada en el Laboratorio de Análisis Instrumental de la Escuela de Ciencias de la Universidad Nacional de Colombia, sede Medellín en un cromatógrafo de gases 6890-N (Agilent Technologies, USA). La curva para gas metano fue obtenida mediante inyección de diferentes volúmenes de un estándar realizando una regresión de las áreas debajo del pico y las concentraciones (volúmenes) de gas metano inyectado.
Análisis estadístico
Los resultados de los sistemas de producción (SSPi y confinamiento) se analizaron como dos experimentos independientes, utilizando en cada caso un diseño en bloques completos al azar. Cada sistema de producción contaba con cinco tratamientos (mezclas de gramíneas: leguminosas en proporciones de 90:10, 80:20 y 70:30, mas gramínea y leguminosa solas), con dos repeticiones por cada horario de estudio (cuatro tiempos de sacrificio: 24, 48, 72 y 96 h) y tres inóculos diferentes, siendo esta última la variable de bloqueo y la repetición estadística.
Las variables dependientes fueron: Producción acumulada de gas (ml), desaparición de MS (g), parámetros obtenidos mediante el modelo de Gompertz, producción neta de metano (mg) y por gramo de materia seca incubada (MSI) y porcentaje de energía bruta (EB) y energía digestible (ED) transformada en metano. Las primeras dos variables están en función del tiempo. La separación de las medias se hizo mediante la prueba de Tukey. La normalidad (P> 0,05) se evaluó con la prueba de Shapiro-Wilk sobre los residuales originales. Para los análisis estadísticos se usó el software SAS®, versión 9.2 (SAS Institute Inc., Cary, NC, USA, 2003).
Resultados y discusión
Calidad nutricional
El contenido de proteína de ambas leguminosas fue casi tres veces el del pasto guinea, y más de cinco veces el encontrado en el pasto ángleton (Tabla 1). Un alto contenido de proteína ha sido frecuentemente reportado en leucaena, siendo 21,6, 22,3 y 30,0% en los estudios de Cardona et al(10), Promkot y Wanapat(41) y Yousuf et al(55), respectivamente. Por su parte, Tedonkeng et al(44) reportaron niveles de proteína de 24,88 y 28,02% en muestras de leucaena cosechada en época seca o lluviosa, respectivamente.
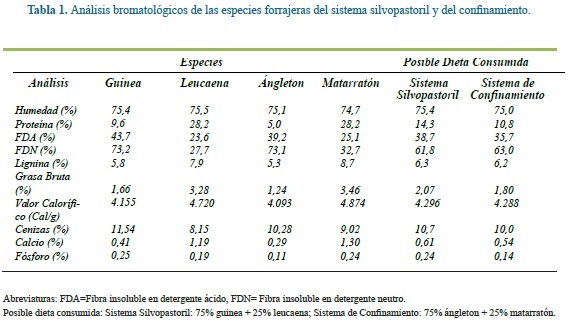
Ambas gramíneas se caracterizaron por tener altos contenidos (i.e. más de 73%) de pared celular (FDN). Por el contrario, el contenido de este componente fue muy bajo (menor al 33%), en las leguminosas. Barahona y Sánchez(6) reportaron que el contenido de pared celular de los recursos forrajeros oscila entre el 30 y el 80% de la materia orgánica (MO), y que, a mayor contenido, mayor es la limitante para la productividad de los animales que los pastorean. Esto está asociado con el consumo de MS del animal, dado que a mayor contenido de fibra, mayor es su tiempo de retención en el rumen, disminuyendo así el consumo voluntario de alimentos.
Los contenidos de hemicelulosa (FDN - FDA) fueron de 29,5 y 33,9% en las gramíneas (guinea y ángleton, respectivamente), siendo muy bajos en las leguminosas (i.e. leucaena: 4,1%, matarratón: 7,6%).
En estudios previos, Barahona y Sánchez(6) reportaron contenidos de proteína, FDN, FDA y Ca similares a los encontrados en el presente trabajo. Sin embargo, el contenido de proteína y cenizas encontrado en nuestro trabajo es superior en 1,5% y 1,2% respectivamente, y 4% menor en FDN que los reportados por Arreaza et al3, quienes trabajaron un pasto de mayor edad.
Al comparar el contenido de grasa bruta de los forrajes, se observó que el de las gramíneas es dos veces inferior al de las leguminosas (3,28 y 3,46 % para leucaena y matarratón, respectivamente). Sin embargo, Yousuf et al(55) reportaron el doble del contenido de grasa en leucaena y matarratón que lo encontrado en nuestro estudio.
Aunque el contenido de cenizas fue mayor en las gramíneas (10,9%) que en las leguminosas (8,6%), las leguminosas tenían 3,5 veces más contenido de calcio que las gramíneas (1,25 vs 0,35%), siendo ángleton la especie con menores aportes de Ca, seguida de guinea. Por el otro lado, en el caso de fósforo no se observaron diferencias entre gramíneas y leguminosas. Sin embargo, la relación entre el calcio y el fósforo fue 5,4 veces en el caso de las leguminosas y 2,6 veces para las gramíneas. Este mismo patrón fue reportado por Yousuf et al(55) al comparar el valor nutricional de una dieta suministrada a cabras y compuesta por guinea, leucaena y matarratón.
Al estimar el contenido de nutrientes en las dietas de ambos sistemas, asumiendo un consumo de 25% de leguminosa y 75% de gramínea, se observa un balance más favorable para el SSPi dado que hay mayor contenido de nutrientes como calcio, fósforo, grasa bruta y proteína, lo que se reflejaría en mayor productividad animal. Esto explica porque la inclusión de estas leguminosas a las praderas con gramíneas nativas ayuda al reciclaje de nutrientes, la recuperación de la biota, la fertilidad del suelo y la diversidad biológica, entre otras(1, 34, 35, 28, 39).
Producción de Gas
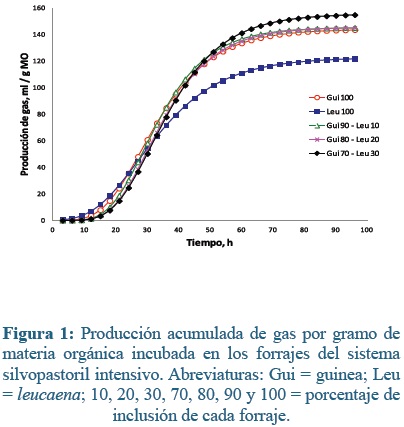
Los forrajes incubados presentaron incrementos en la producción de gas por gramo de materia orgánica (MO) a través del tiempo. En la acumulación de gas para las muestras de forrajes del SSPi (Figura 1) a las 24 h hubo mayor acumulación de gas para Gui100 y Leu100 que en la de Gui70- Leu30 (34,4, 35,4 y 24,5 ml g-1 MO, respectivamente, P= 0,045). A las 48 h no se observaron diferencias en acumulación de gas entre las muestras (P= 0,86); todos los forrajes y sus mezclas alcanzaron producciones de alrededor de 115 ml de gas g-1 MO en este horario, lo que corresponde aproximadamente entre el 77 y 83% del gas total producido durante el experimento. Esto es similar a lo encontrado por Barahona et al5, quienes reportaron acumulaciones a las 48 h de 110 y 104 ml según el estado de madurez de leucaena. A las 72 h pos-inoculación, la máxima tasa de producción fue de 149,9 ml g-1 MO para Gui70-Leu30, 32,5 ml mayor que observada con Leu100 (P=0,0046). A las 96 h de incubación, esta diferencia se aumentó a 28 ml, con el resto de los tratamientos mostraron una acumulación promedio de gas de 142 ml g-1 MO (P=0,006).
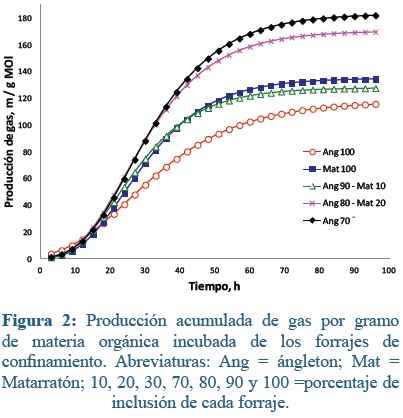
En los forrajes del sistema de confinamiento (Figura 2) a las 24 h pos-inoculación, la máxima acumulación de gas fue para Ang80-Mat20 (61 ml g-1 MO) y la mínima fue para Ang100 con una diferencia del 33% (P=0,006). Entre las 48 y 72 h las producciones de gas (ml g-1 MO) oscilaron entre 89,6 (Ang100) y 176,8 (Ang70-Mat30) (P≤ 0,05). A las 96 h los valores máximos se obtuvieron en las mezclas Ang70-Mat30 y Ang80-Mat20, seguidos de Mat100, Ang90-Mat10 y Ang100 (134,5; 127,7; 115,9 ml; respectivamente), esta ultima estaba 66 ml de gas g-1 MO por debajo de la mayor tasa de producción en este horario (P=0,006).
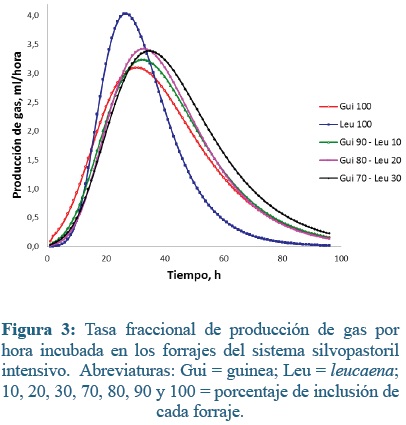
La técnica de producción de gases, a diferencia de otras técnicas in vitro, ofrece la posibilidad de establecer el curso y la importancia de la fracción soluble del sustrato dentro del proceso fermentativo(40). En las primeras horas de fermentación, una parte del sustrato, principalmente los azúcares solubles son fermentados inmediatamente. Sin embargo, los azúcares solubles generalmente sólo constituyen una pequeña parte del material potencialmente digestible(44). A medida que el proceso fermentativo avanza, el material es hidratado y colonizado por los microorganismos ruminales lo que origina diferentes tasas de degradación dependiendo de la concentración de carbohidratos estructurales. Esto se evidencia en las figuras 3 y 4, donde se muestran las producciones de gas (ml) por hora para ambos sistemas de producción.
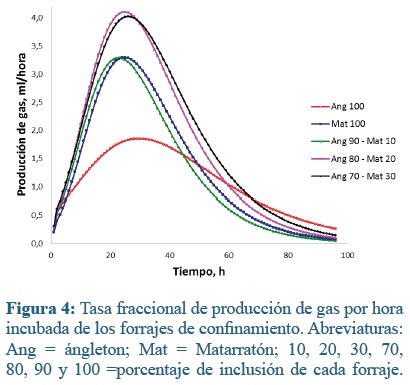
En el sistema de confinamiento, Ang80-Mat20 y Ang70-Mat30 obtubieron las máximas tasas fraccionales de producción de gas (4,11 y 4,03 ml h-1, respectivamente), seguidas de Mat100 y Ang90-Mat10 con aproximadamente 0,77 ml por h menos (3,30 y 3,29 ml h-1) esto ocurrió entre las 22 y 26 h, la menor tasa la produjo Ang100, 6 más tarde (1,85 ml h-1).
En el sistema silvopastoril, las máximas tasas fraccionales de gas (ml * h-1) de las materias primas incubadas, se encuentran entre las 27 y 35 h, siendo Leu100 la que produce mayor gas en menor tiempo (4,03 ml/h a las 27h), el resto de forrajes 5 horas después, produjeron aproximadamnte 3,2 ml /h, sin embargo Leu100 tiene una caída más abruta que las otras, es por esto que tiene la menor producción total acumulada.
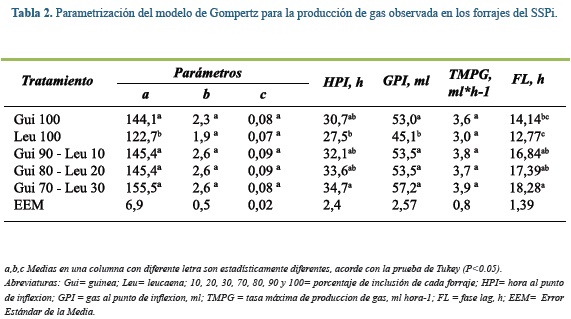
Al analizar las curvas de acumulación de gas de las muestras del SSPi con el modelo de Gompertz, se encontró que el parámetro a, que corresponde a la producción máxima de gas, fue menor en la leucaena que guinea o las mezclas evaluadas (P<0,05, Tabla 2). Lo mismo sucedió con el parámetro GPI, donde leucaena al 100% mostró menores valores que el resto de los forrajes (P<0,05). La máxima hora al punto de inflexión (HPI) fue para Gui70- Leu30 (34,7 h), aproximadamente 7 h menos que Leu100 (27,55 h), igual tendencia mostro el tiempo de colonización microbiana (FL; 18,28 vs 12,77 h, Gui70 - Leu30 y Leu100, respectivamente. En el resto de parámetros (b, c y TMPG), no se observaron diferencias significativas entre los tratamientos.
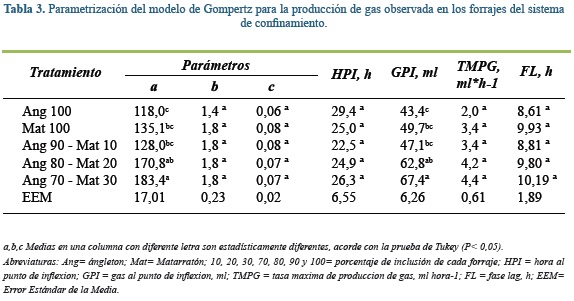
En los forrajes del confinamiento el modelo de Gompertz mostró que el parámetro a (producción máxima de gas) fue menor en ángleton 100% que en las mezclas con 20 y 30% de matarratón (Tabla 3; P< 0,05). A su vez, la mezcla con 30% de matarratón tuvo mayor producción de gas que la muestra con 10% de matarratón o el matarratón al 100%. Esta misma situación se dio para el parámetro GPI. Para los parámetros b, c, HPI y TMPG, no se observaron diferencias significativas entre los tratamientos. El tiempo promedio de colonización microbiana fue 9,4 h en promedio.
Degradabilidad de la materia seca
En las tabla 4 y 5 se muestra la degradabilidad de la materia seca (DMS) a las 24, 48, 72 y 96 h de incubación. Durante estos cuatro horarios de mediciones y en ambos sistemas de producción las leguminosas incubadas al 100% tuvieron mayores tasas de degradación que las gramíneas solas (P≤0,05), siendo esta diferencia de alrededor del 17%. En las muestras del SSPi, leucaena tuvo mayor DMS que las mezclas gramínea - leguminosa con excepción de Gui70-Leu30 a las 48, 72 y 96 h.
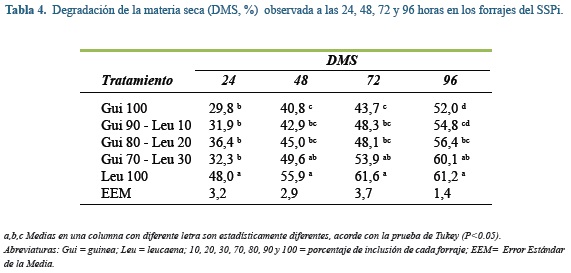
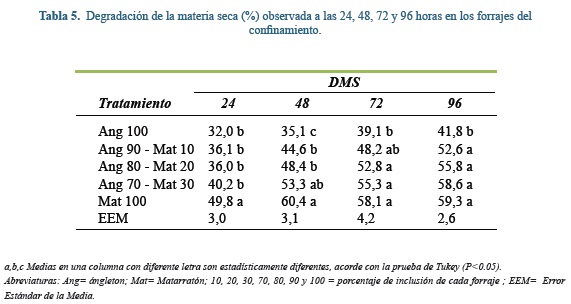
Al comparar la DMS en ambos sistemas de producción, se observa que a mayor cantidad de inclusión de gramínea en la incubación, menor fue la degradación de la MS. Esto es explicado por el alto contenido de fibra de las gramíneas, ya que la digestibilidad de los forrajes está inversamente relacionada con su contenido de FDN y este a su vez está en función de la composición interna y su estructura(6).
En ambos sitemas, la mayor DMS ocurrió durante las primeras 24 h de fermentación, para ese tiempo, al menos el 50% de la degradación final de la MS (i.e. 96 h) ya había ocurrido en todos los forrajes o sus mezclas. Es importante aclarar que dicha degradación no necesariamente es sinónimo de utilización, sino que puede estar más relacionada con la solubilización de los componentes de molienda más pequeños(38) ó rápidamente fermentables en el medio de incubación. Este concepto se demuestra al observar que en las primeras 24 h solo se acumuló en promedio 28,6% del total de gas en todos los tratamientos (rango: 16,1 a 41,4%).
Al comparar la DMS de leucaena con la reportada por Barahona et al(5) después de 144 h de incubación de muestras maduras de leucaena, se encuentra que dicho valor (61,5%) es el mismo que el encontrado en este trabajo hacia las 72 y 96 h de incubación. Algo diferente ocurre con la DMS del ángleton reportada por Sánchez et al(42), quienes reportaron desapariciones a las 48 h, 11% más altas (DMS: 46%) que las encontradas en este trabajo, a pesar de que los forrajes utilizados en ambos estudios presentan contenidos similares de nutrientes.
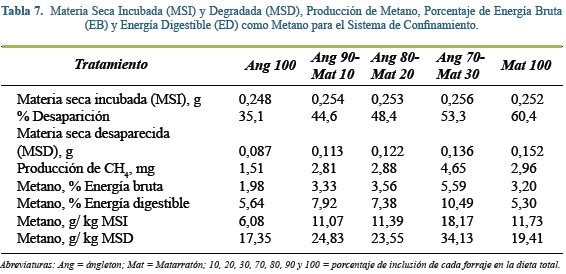
Producción de metano
Los análisis de la producción neta de metano (PCH4) fueron realizados para el gas generado a las 48 h de incubación de aproximadamente 300 mg de forrajes típicos suministrados en dietas de un SSPi (Tabla 6) y confinamiento (Tabla 7), su DMS osciló entre 35 y 60%. Se debe de recordar que a las 48 h pos inicio de la incubación se obtuvieron las máximas tasas fraccionales de producción de gas y aproximadamente el 82% del gas total producido durante el experimento.
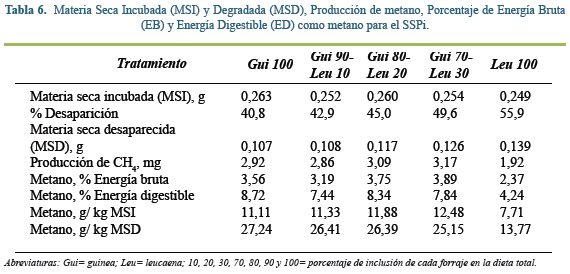
En todos los tratamientos del SSPi, la PCH4 no difirió significativamente (P=0,67), a pesar que el tratamiento Gui 100 y las diferentes mezclas (Gui90- Leu10, Gui80- Leu20; Gui70- Leu30) presentaron emisiones de alrededor de 3,0 mg CH4, un 36% más que lo observado con Leu100 (1,9 mg CH4). Similares valores fueron reportados por Singh et al(43) en un experimento in vitro para guinea (11,5 g CH4 kg-1 MS y 28,3 g CH4 kg-1 MSD), pero los valores reportados por este autor para leucaena son muy superiores a los observados en la presente investigación. La diferencia ente leguminosas y gramíneas podría deberse en parte a la presencia de taninos condensados (TC), ya que se ha reportado que la inclusión TC reduce las emisiones de metano(24, 53, 45), y el forraje de Leucaena leucocephala contiene hasta un 10% de TC(5).
Por su parte, en los forrajes del confinamiento, la acumulación de metano varió entre 1,5 mg en las muestras de Ang100 a 3,36 mg en la mezcla con 70% de ángleton. Resultados similares fueron reportados por Kurihara et al(26), al comparar la producción de metano por animales Bos indicus que pastoreaban una pradera con solo ángleton (113 g d-1) y una dieta a base de grano y alfalfa (Medicago sativa; 160 g d-1). En ambos estudios se puede observar una relación entre la calidad nutricional y la producción de gas. En el presente experimento, la inclusión de la leguminosa en el sistema de confinamiento estuvo asociada con mayores emisiones de metano, dada la pobre calidad nutricional de la gramínea incluida, ya que el contenido de proteína del ángleton fue de sólo 5%, siendo a este nivel limitante para la fermentación(30). Se ha sugerido que el disminuir la proteína en 1% la producción de metano, puede aumentar en 6,22 kg-1 la MS consumida54. Sin embargo, Meale et al31 no encantaron relación entre la emisión de metano y la calidad nutricional del sustrato fermentado. Al incluirse el matarratón, el contenido de proteína de la mezcla pasó a 7,32; 9,64 y 11,96%, con inclusiones de 10, 20, y 30% de matarratón, respectivamente.
Al calcular la proporción entre la producción de metano (g) y la cantidad de MS incubada (Kg) para el SSPi, este valor fue 35% inferior con Leu 100 que con los otros tratamientos (en promedio 11,7 g CH4 Kg-1 MS). En ambos sistemas, no se encontraron diferencias estadísticamente significativas. Cuando se relaciona el CH4 con la MS degradada, se observa una tendencia a disminuir las emisiones a medida que aumenta la proporción de leguminosa en la dieta en los forrajes del SSPi (y = -0.1407x + 28.295, R² = 0.9724). Johnson y Johnson(23) y McAllister et al(29), afirmaron que el tiempo de degradación influye directamente sobre la producción de metano, así los carbohidratos estructurales son más lentos en su tasa de fermentación, produciendo más gas por unidad de sustrato digerido. Sin embargo, para el sistema de confinamiento, la adición de cantidades crecientes de matarratón (entre 0 y 30%) estuvo asociada con aumentos en la emisión de metano por kilogramo de MSD.
En cuanto al porcentaje de energía digestible que se pierde en forma de metano, en ambos sistemas de producción la leguminosa presentó menores pérdidas (Leu100= 4,30; Mat100= 5,30) que las gramíneas y sus respectivas mezclas, siendo más marcada esta diferencia para el SSPi, donde la pérdida se reduce 50% para la Leu100 (Tabla 6). Esto puede ser explicado en parte por los componentes de FDN y FDA, ya que estos dos nutrientes se correlacionan con las emisiones de metano de forma negativa. Varios autores aseguran que los carbohidratos de la pared emiten más gas que los solubles(2, 32, 49, 54). Las gramíneas de trópico se caracterizan por tener paredes vegetales muy lignificadas, en especial las plantas C4 que pueden llegar a producir hasta un 30% más de metano que las leguminosas que están bajo las mismas condiciones medioambientales(2). Estos valores se relacionan con los reportados por De Ramus et al(12) y Carmona et al(11), quienes afirmaron que las emisiones anuales de metano por novillas de carne en pastoreo estuvieron entre 32 y 83 kg dia-1 y entre 60 y 95 kg dia-1 para vacas adultas que pastoreaban diferentes tipos de praderas, donde la emisión más alta correspondió a gramíneas de baja calidad nutricional, con sistemas de pastoreo continuo y baja disponibilidad forrajera, mientras que las emisiones más bajas correspondieron a praderas mejoradas, a sistemas de pastoreo rotacional y con alta disponibilidad de forraje.
Conclusiones y Recomendaciones
En ambos sistemas de producción, las leguminosas tuvieron mayores contenidos de proteína cruda, grasa, energía bruta y calcio y menores contenidos de FDN, FDA y cenizas. En general, estos cambios en contenidos de nutrientes se consideran como ventajas de la inclusión de leguminosas en los SSPi u otras asociaciones. Así, a medida que se aumentó la inclusión de leguminosas en las mezclas gramínea - leguminosa, se observó un aumento en la digestibilidad in vitro de la materia seca, el cual fue más pronunciado en el caso del confinamiento, probablemente debido al bajo contenido de proteína en el pasto ángleton evaluado en este estudio.
En todos los forrajes, la mayor producción de metano ocurrió después de las primeras 24 h de incubación. Hubo gran variabilidad en las emisiones entre los diferentes forrajes y mezclas evaluadas, lo cual probablemente obedeció a las diferencia en contenido de nutrientes en cada uno de ellos. La inclusión de leucaena mostró una tendencia a reducir las emisiones de metano a medida que se incrementó el contenido de leucaena en la mezcla fermentada. Dicha respuesta no se observó en el caso del confinamiento en respuesta a la inclusión de matarratón. Se recomienda realizar más experimentos, tanto in vitro como in vivo, para caracterizar mejor esta respuesta y determinar a qué niveles de inclusión se mejora la producción animal sin aumentar las emisiones de metano. En este trabajo, las emisiones de metano mostraron estar relacionados con la calidad nutricional del alimento, por lo que se recomienda realizar experimentos in vivo en donde la emisión de metano esté asociada con la productividad animal, a fin de determinar las combinaciones en las cuales se obtengan adecuadas producciones con las menores emisiones de metano posibles.
Agradecimientos
Los autores agradecen al Ministerio de Agricultura y Desarrollo Rural por la financiación del proyecto "Análisis comparativo de la Producción de Carne de Novillos Cebú en Sistema Silvopastoril Intensivo o en Confinamiento" del programa "Análisis Multifactorial de la Calidad Funcional de la Carne en Búsqueda de un Producto con Valor Agregado" que permitió la realización de este estudio y al Profesor Ricardo Rosero del Laboratorio GRICA de la Universidad de Antioquia, sitio donde se realizaron los experimentos de producción de gas.
Referencias
1. Alonso J. Los sistemas silvopastoriles y su contribución al medio ambiente. Rev Cubana Cienc Agríc 2011; 45(2): 107-115. [ Links ]
2. Archimède H, Eugène M, Marie Magdeleine C, Boval M, Martin C, et al. Comparison of methane production between C3 and C4 grasses and legumes. Anim Feed Sci Technol 2011; 166: 59-64. [ Links ]
3. Arreaza L, Sánchez D, Abadia B. Degradación de fracciones de carbohidratos en forrajes tropicales determinada por métodos in vitro e in situ. Corpoica Cienc Tecnol Agropecu 2005; 6 (1): 52-57. [ Links ]
4. Association Of Official Analytical Chemists (AOAC). Official Method 942.05. Determination of Ash in Animal Feed. In Official Methods of Analysis of AOAC International. 18th edition (Chapter 4). Gaithersburg, MD, USA: AOAC International. 2005. p. 8. [ Links ]
5. Barahona R, Lascano C, Narváez N, Owen E, Morris P, et al. In vitro degradability of mature and immature leaves of tropical forage legumes differing in condensed tannin and non-starch polysaccharide content and composition. J Sci Food Agric 2003; 83:1256-1266. [ Links ]
6. Barahona R y Sánchez M. Limitaciones físicas y químicas de la digestibilidad de pastos tropicales y estrategias para aumentarla. CORPOICA Cienc Tecnol Agropecu 2005; 6 (1): 69-82. [ Links ]
7. Boadi, D. A., Wittenberg, K. M. y McCaughey, W. P., 2002. Effects of grain supplementation on methane production of grazing steers using the sulphur (SF6) tracer gas technique. Can. J. Anim. Sci. 82, 151-157. [ Links ]
8. Boadi, D., Benchaar, C., Chiquette, J., Masse, D., 2004. Mitigation strategies to reduce enteric methane emissions from dairy cows: update review. Can. J. Anim. Sci. 84, 319-335. [ Links ]
9. Calsamiglia S, Castillejos L, Busquet M. Estrategias nutricionales para modificar la fermentación ruminal en vacuno lechero. En: XXI Curso De Especialización, FEDNA; España. 2005: 161- 185. [ Links ]
10. Cardona M, Sorza J, Posada S, Carmona J, Ayala S, et al. Establecimiento de una base de datos para la elaboración de tablas de contenido nutricional de alimentos para animales. Rev Col Cienc Pec 2002; 15 (2): 240-246. [ Links ]
11. Carmona JC, Bolívar DM, Giraldo LA. El gas metano en la producción ganadera y alternativas para medir sus emisiones y aminorar su impacto a nivel ambiental y productivo. Rev Col Cienc Pec 2005; 18 (1): 49-63. [ Links ]
12. de Ramus HA, Clement TC, Giampola DD, Dickison PC. Methane emissions of beef cattle on forages: efficiency of grazing management systems. J Environ Qual 2003; 32: 269-277. [ Links ]
13. Federación Colombiana de Ganaderos (FEDEGAN). Plan Estratégico de la Ganadería Colombiana 2019. Bogotá, Colombia 2009; 294 p. [ Links ]
14. Food And Agriculture Organization (FAO). La ganadería amenaza el medio ambiente. Roma 2006; [acceso: 09 de enero de 2013]: URL: http://www.fao.org/newsroom/es/news/2006/1000448/index.html. [ Links ]
15. Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC). NTC 668. Alimentos y materias primas. Determinación de los contenidos de grasa y fibra cruda. Bogotá D.C 1973. p. 8. [ Links ]
16. Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC). NTC 4657. Alimento para animales. Determinación del contenido de nitrógeno y cálculo del contenido de proteína cruda. Método Kjeldahl. Bogotá D.C 1999. [ Links ]
17. Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC). NTC 4981. Alimentos para animales. Determinación del contenido de fósforo. Método espectrofotométrico. Bogotá D.C 2001. [ Links ]
18. Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC). NTC 5151. Alimento para animales. Determinación de los contenidos de calcio, cobre, hierro, magnesio, manganeso, potasio, sodio y zinc. Método usando espectrometría de absorción atómica. Bogotá D.C 2003. [ Links ]
19. Instituto De Hidrología Meteorología Y Estudios Ambientales (IDEAM). Capítulo dos Inventario Nacional de Gases de Efectos invernadero. Bogotá, Colombia 2008; 119-152. [ Links ]
20. International Organization for Standardization. Animal feeding stuffs, animal products, and faeces or urine - Determination of gross calorific value - Bomb calorimeter method. ISO 9831. Geneva, Switzerland 1998. [ Links ]
21. International Organization for Standardization. Animal feeding stuffs. Determination of moisture and other volatile matter content. ISO 6496. Geneva, Switzerland 1999. p. [ Links ]
22. Jarvis GN, Strömpl C, Burgess DM, Skillman LC, Moore ER, et al. Isolation and identification of ruminal methanogens from grazing cattle. Curr microbial 2000; 40 (5): 327-332. [ Links ]
23. Johnson K A y Johnson DE. Methane emissions from cattle. J Anim Sci 1995; 73 (8): 2483-2492. [ Links ]
24. Kamalak A, Canbolat O, Gurbuz Y, Ozay O, Ozkan CO, et al. Chemical composition and in vitro gas production characteristics of several tannin containing tree leaves. LRRD 2004; 16 (6). [ Links ]
25. Kamra DN. Rumen microbial ecosystem. Curr Sci 2005; 89 (1): 124-135. [ Links ]
26. Kurihara M, Magner T, Hunter RA y McCrabb G J. Methane production and energy partition of cattle in the tropics. Br J Nutr 1999; 81: 227-234. [ Links ]
27. Liu Y y Whitman WB. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann N Y Acad Sci 2008; 1125: 171-189. [ Links ]
28. Mahecha L. El silvopastoreo: una alternativa de producción que disminuye el impacto ambiental de la ganadería bovina. Rev Col Cienc Pec 2002; 15: 49-63. [ Links ]
29. McAllister TA, Cheng KJ, Okine EK y Mathison GW. Dietary, environmental and microbiological aspects of methane production in ruminants. Can J Anim Sci 1996; 76 (2): 231-243. [ Links ]
30. McIvor JG y Chen CP. Tropical grasses: their domestication and roles in animal feeding systems. En: G. J. Blair, D. A. Ivory, and T. R. Evans (eds.). Forages in Southeast Asia and South Pacific Agriculture. Proceedings of an international workshop held at Cisarua, Indonesia, ACIAR; 1986 p 55. [ Links ]
31. Meale SJ, Chaves AV, Baah J y McAllister TA. Methane Production of Different Forages in vitro Ruminal Fermentation. Asian-Aust J Anim Sci2012; 25(1):86-91. [ Links ]
32. Moe PW y Tyrell HF. Methane production in dairy cows. J Dairy Sci 1979; 62 (10):1583-1586. [ Links ]
33. Moss AR, Jouany JP y Newbold J. Methane production by ruminants: its contribution to global warming. Ann Zootech 2000; 49 (3): 231-253. [ Links ]
34. Murgueitio E. Impacto ambiental de la ganadería de leche en Colombia y alternativas de solución; LRRD 2003; 15 (10). [acceso: Mayo 10, 2012]. URL: http://www.lrrd.org/lrrd15/10/murg1510.htm. [ Links ]
35. Murgueitio E y Calle Z. Diversidad biológica en sistemas de ganadería bovina en Colombia. En Conferencia electrónica de la FAO sobre Agroforestería para la producción animal en Latinoamérica. Roma: FAO. 1998; 27-46. [ Links ]
36. Murgueitio E. Calle Z, Uribe F, Calle A, Solorio B. Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest Ecol Manag 2011; 261 (10):1654-1663 [ Links ]
37. Murray RM, Bryant AM y Leng RA. Rates of production of methane in the rumen and large intestine of sheep. Br J Nutr 1976; 36 (1):1-14. [ Links ]
38. Ørskov ER. The In situ Technique for the Estimation of Forage Degradability in Ruminants. Forage Evaluation in Ruminant Nutrition (eds D.I. Givens, E. Owen, R.F.E. Axford and H.M. Omed), 2000:175-188. [ Links ]
39. Pagiola S, Ramírez E, Gobbi J, Haan C, Ibrahim M, et al. Paying for the environmental services of silvopastoral practices in Nicaragua. Ecol Econom 2007; 64(2):374-385. [ Links ]
40. Posada SL y Noguera RR. Técnica in vitro de producción de gases: Una herramienta para la evaluación de alimentos para rumiantes. LRRD 2005; 17. [acceso: Mayo 10, 2012]. URL: http://www.lrrd.org/lrrd17/4/posa17036.htm. [ Links ]
41. Promkot C y Wanapat M. Ruminal degradation and intestinal digestion of crude protein of tropical protein resources using nylon bag technique and three-step in vitro procedure in dairy cattle. LRRD 2003; 15 (11). [acceso: Mayo 10, 2012]. URL: http://www.lrrd.org/lrrd15/11/prom1511.htm. [ Links ]
42. Sánchez D, Arreaza L, Abadía B. Estudio de la cinética de degradación in vitro de cuatro forrajes tropicales y una leguminosa de clima templado. Corpoica Cienc Tecnol Agropecu 2005; 6 (1). [ Links ]
43. Singh S, Kushwaha BP, Nag SK, Mishra AK, Singh A, et al. In vitro ruminal fermentation, protein and carbohydrate fractionation, methane production and prediction of twelve commonly used Indian green forages. Anim Feed Sci Technol 2012; 178 (1):2-11. [ Links ]
44. Stefanon B, Pell AN y Schofield P. Effect of maturity on digestion kinetics of water-soluble and water-insoluble fractions of alfalfa and brome hay. J Anim Sci 1996; 74 (5):1104-1115. [ Links ]
45. Tan HY, Sieo CC, Abdullah N, Liang JB, Huang XD, et al. Effects of condensed tannins from Leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro. Anim Feed Sci Technol 2011; 169 (3):185-193. [ Links ]
46. Tedonkeng E, Fonteh F, Tendonkeng F, Kana J, Boukila B, et al. Influence of supplementary feeding with multipurpose leguminous tree leaves on kid growth and milk production in the West African dwarf goat. Small Ruminant Res 2006; 63: 142-149. [ Links ]
47. Tavendale, M. H., Meagher, L. P., Pacheco, D. Walker, N., Attwood, G. T. and Sivakumaran, S., 2005. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Feed Sci. Technol. 123:403-419. [ Links ]
48. Theodorou MK, Williams BA, Dhanoa MS, McAllan AB and France J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim Feed Sci Technol 1994; 48:(3-4):185-197. [ Links ]
49. Ulyatt MJ y Lassey K R. Methane emissions from pastoral systems: the situation in New Zealand. Arch Latinoam Prod Anim 2001; 9(1):118-126. [ Links ]
50. Van Soest PJ, Robertson JB y Lewis BA. Methods for dietary fiber neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 1991; 74 (10):3583-3597. [ Links ]
51. Van Soest PJ. Use of detergents in the analysis of fibrous feed. II. A rapid method for the determination of fiber and lignin. J Assoc Off Anal Chem 1963; 46 (5):829. [ Links ]
52. Walsh C, O'Regan B y Moles R. Incorporating methane into ecological footprint analysis: A case study of Ireland. Ecol Econom 2009; 68(7): 1952-1962. [ Links ]
53. Woodward SL, Waghorn GC, Laboyrie P. Condensed tannins in birdsfoot trefoil (Lotus corniculatus) reduced methane emissions from dairy cows. Proc N Z Soc Anim Prod 2004; 64:160-164. [ Links ]
54. Yan T, Mayne CS y Porter MG. Effects of dietary and animal factors on methane production in dairy cows offered grass silage based diets. En International Congress Series 2006; 1293:123-126. [ Links ]
55. Yousuf MB, Belewu MA, Daramola JO y Ogundun NI. Protein supplementary values of cassava, leucaena and gliricidia leaf meals in goats fed low quality Panicum maximum hay. LRRD 2007; 19. [acceso: Mayo 10, 2012]. URL: http://www.lrrd.org/lrrd19/2/yous19023.htm. [ Links ]
