










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCES Medicina Veterinaria y Zootecnia
On-line version ISSN 1900-9607
Ces. Med. Vet. Zootec. vol.9 no.1 Medellín Jan./June 2014
Evaluación morfométrica del cráneo de diversas razas bovinas actuales: análisis geométrico según perfiles
Morphometric evaluation of the skull in several bovine breeds: geometric analysis according to their profiles¤
Avaliação morfométrica do crâneo de diversas raças bovinas atuais: analise geométrico segundo seus perfis
Pere M. Parés-Casanova1*, MV, Dr.
1*Autor para correspondencia: Departament de Producció Animal. Universitat de Lleida. Av. Alcalde Rovira Roure, 191. E 25198 Lleida (Catalunya, España) .Tel +34 973 70 64 60. Fax + 34 973 70 28 74. peremiquelp@prodan.udl.cat
¤Para citar este artículo: Parés-Casanova PM. Evaluación morfométrica del cráneo de diversas razas bovinas actuales: análisis geométrico según perfiles. Rev CES Med Zootec. 2014; Vol 9(1): 58-67.
(Recibido: 13 de noviembre, 2013; aceptado: 25 de abril, 2014)
Resumen
La morfometría geométrica permite la captura y análisis de datos en forma de matrices de morfocoordenadas que representan la geometría de un espécimen y no se limita a la obtención de datos lineales de él como medidas de alto o ancho, elementos que carecen de la precisión y la riqueza de los datos geométricos. Se analizó la variación de perfil fronto-nasal de diversas razas bovinas actuales mediante métodos de este tipo. El análisis morfométrico geométrico constó de tres etapas fundamentales: obtención de los datos primarios, obtención de las variables de la forma, y análisis estadístico. La obtención de los datos primarios se realizó a partir de 15 hitos y dos semi-hitos homólogos, situados sobre la vista lateral del cráneo de 19 individuos adultos representativos de cada raza. Los resultados obtenidos, aplicando los test estadísticos habituales en los análisis de morfometría geométrica, revelaron que el perfil fronto-nasal no exhibe configuraciones craneales (de tamaño y forma) diferenciadas, por lo que forma y tamaño craneales no tienen relación alguna con la tipificación aloídica, presentando además ambas variables una falta de correlación mutua.
Palabras clave: Aloidismo, alometría, cráneo, morfología geométrica, perfil cefálico.
Abstract
Geometric morphometrics allows the acquisition and analysis of shape coordinates representing the geometry of a specimen and is not limited to obtaining linear data such as height or width which are not enough precise compared with geometric data. The fronto-nasal profile variation of various bovine breeds was analysed using this method. The analysis consisted of three steps: collection of primary data, obtaining shape variables, and statistical analysis. The primary data were obtained from 15 homologous landmarks and two homologous semi-landmarks. These points were taken on the lateral view of the skull of 19 adult individuals representative of each breed. The results obtained by applying standard statistical tests revealed that the cranial profile lacks a differentiating size and shape configuration (absence of fronto-nasal profile correlation with skull shape). Additionally, both variables were not cross-correlated.
Key words: Aloidism, allometry, geometric morphology, head profile, skull.
Resumo
A morfometria geométrica permite a captura e análise de dados em forma de matrizes de morfo-coordenadas que representam a geometria de um espécime e não se limitam a obtenção de dados lineares deles como medidas de alto ou largo, elementos que carecem da precisão e riqueza dos dados geométricos. Analisou-se a variação dos perfis fronto nasais de diversas raças bovinas atuais mediante métodos de este tipo. A análise morfo-geométrica constou de três etapas fundamentais: obtenção dos dados primários, obtenção de variáveis de forma e analise estatístico: A obtenção dos dados primários realizou-se a partir de 15 fossas e dois semi-fossas homólogas, situados sobre a região lateral do crânio de 19 indivíduos adultos representativos de cada raça. Os resultados obtidos aplicando os testes estatísticos habituais nos analises de morfometria geométrica, revelaram que os perfis fronto nasais não exibem configurações craniais (de tamanho e forma) diferenciadas, pelo qual a forma e o tamanho cranial não tem relação alguma com a tipificação aloidica, apresentando além as duas variáveis uma falta de correlação.
Palavras chave: Aloidismo, alometria, crânio, morfologia geométrica, perfil cefálico.
Introducción
En los últimos años ha habido un notable aumento en el uso de la morfometría geométrica versus la tradicional morfometría multivariada 22, conocida en biometría como “morfometría tradicional”, se aplica a grupos de variables cuantitativas, como largo, ancho, alto y ángulos 28. La morfometría geométrica, en lugar de enfocarse en el análisis de un conjunto de medidas lineales entre puntos morfométricos, como hace la tradicional, se basa en el estudio de los cambios de tamaño y forma a partir del desplazamiento de un conjunto de puntos morfométricos en el plano bi o tridimensional18, 28. La relación espacial en dimensiones de estos puntos se conserva a lo largo de todo el análisis morfométrico, lo que permite “reconstruir” con tanta precisión como se desee la forma y el tamaño de los objetos estudiados 18. Por todo ello, la morfometría geométrica se ha convertido en una herramienta fundamental y enormemente potente, para el estudio de cualquier estructura fenotípica. Otra ventaja de este método es que suele proporcionar una mejor visualización de los resultados22 y, por ende, facilita su interpretación. Algunos autores 6, 10, 26, 27 permiten profundizarse en lo que se refiere a los tratamientos generales del tema de la morfometría geométrica.
Variaciones fenotípicas y genéticas a través del espacio geográfico están presentes en muchas especies de mamíferos7, 9, 20. Estas son determinadas por procesos históricos o ecológicos, incluyendo estos últimos, tanto factores bióticos como físicos 10, 11. En los estudios sobre variabilidad morfológica de los mamíferos domésticos y salvajes, suele darse especial énfasis a los cambios fenotípicos centrados en las estructuras esqueléticas16, y concretamente cráneo y mandíbula. Juega pues, la craneometría, un papel preeminente en las investigaciones osteológicas 1, 2, 4, 17, 21, constituyendo el cráneo un elemento de gran importancia para la filiación taxonómica y de información sobre los cambios derivados de la selección 8, 13, 14, 1.
Los caracteres étnicos suministrados por la cabeza tienen además gran importancia etnológica, sobretodo porque su somación no está influenciada por el manejo. De los caracteres étnicos suministrados por el cráneo, son las diversas medidas de longitud y anchura y, sobretodo perfil, los más comúnmente utilizados con fines etnológicos 5.
El sistema descriptivo, clásico, de Baron considera los animales domésticos con tres tipos de perfil: el recto, que se considera el tipo medio, el cóncavo, que sería la desviación negativa, y en sentido positivo, el convexo. El perfil recto se denomina ortoide, el perfil cóncavo, celoide, y el convexo, cirtoide. Se usan asimismo las denominaciones ultra y sub, que no se han considerado en este estudio. Se consideran cráneos ortoides aquellos en los que no existe hundimiento alguno del fronto-nasal; celoides, cuando la depresión es netamente marcada; y cirtoides, cuando por el contrario existe saliente en dicha región frontal5. Pero toda la clasificación según el perfil se basa en una mera apreciación visual y de escaso valor como elemento de estudio geométrico, puesto que no hay ningún tipo de cuantificación.
La morfometría tradicional -y con mayor razón los estudios basados en medidas lineales y estadística uni y bivariada- ofrece, como se ha expuesto anteriormente, limitadas herramientas para el estudio de la cabeza, debido a que no puede considerar su geometría. El empleo de la morfometría tradicional presenta importantes limitaciones inherentes al manejo de datos primarios lineales. Entre estos problemas destacan la falta de una adecuada partición de los componentes de la forma en relación a los componentes de tamaño, la ausencia de una definición no ambigua de homología para los hitos anatómicos, la pérdida de poder estadístico debido a la sobreposición de las mediciones que cubren los ejes anatómicos mayores y la incapacidad de reproducir visualmente en dos o tres dimensiones el patrón de variación de la forma de los objetos biológicos 6.
Aunque es sumamente frecuente el uso del análisis lineal en estudios craneométricos en el ámbito de la etnología animal15, las variables obtenidas no llegan a reflejar siempre claramente las diferencias entre grupos o razas, y mucho menos cuando se intenta capturar información relativa al aspecto más importante de la geometría, la forma, que al fin y al cabo es lo que describe un perfil. Esto significa que, el aloidismo -el perfil- no puede describirse en términos de dimensiones craneales, sino a lo sumo de la relación espacial entre sus partes, que únicamente el estudio matemático de la forma puede permitir. A diferencia de la morfometría tradicional, la morfometría geométrica evita reducir la variación morfológica a medidas lineales o angulares, y permite ganar información referente a las relaciones geométricas del conjunto.
La literatura científica refleja la aceptación de los métodos de morfometría geométrica en aquellas disciplinas con una larga historia en el estudio de las formas, como son la Biología y la Arqueología, pero no en etnología veterinaria, donde es escasísimo lo publicado. Y ello, a pesar de que el análisis de la variación craneal en los animales domésticos es crucial, ya que su variación puede responder a diferentes roles funcionales o presiones selectivas en las poblaciones o razas.
Se aplicaron en la presente investigación, métodos de morfometría geométrica para el estudio de perfil fronto-nasal de diversas razas bovinas (Bos taurus y Bos indicus). Se pretende con ello, detectar si la forma y el tamaño -que únicamente pueden estudiarse mediante estos métodos geométricos- pueden tener alguna relación con la tipificación aloídica. Además, si se piensa en la utilidad de los resultados obtenidos mediante la aplicación de los análisis de morfometría geométrica, en comparación con aquellos obtenidos mediante una pura apreciación subjetiva, o incluso mediante morfometría tradicional, se podrá describir cómo se modifica la forma del cráneo según el perfil, dónde se localizan estos cambios dentro de esta área y cuáles son esos cambios, ya que podrán ser fácilmente visualizados.
El estudio resulta inédito, no existiendo, a conocimiento de los autores, investigaciones similares publicadas. Sus antecedentes son pues generales y por ende, los resultados obtenidos, no vinculantes.
Materiales y métodos
Material examinado
El presente estudio se realizó sobre cráneos de diversas razas bovinas actuales, procedentes de matadero. Los especímenes fueron obtenidos en fresco, en un matadero frigorífico industrial. El mismo día fueron trasladados y dejados a la intemperie, en un área controlada, hasta su “limpieza” (proceso que duró unos 4-5 años). Se contó finalmente con 19 cráneos bovinos sin patologías óseas pertenecientes a diversas razas bovinas españolas y francesas (Aubrac, “Bruna dels Pirineus”, Charolesa, Frisona, Rubia de Aquitania y Salers). Los cráneos fueron posteriormente desarticulados de la mandíbulas y clasificados individualmente de acuerdo a su perfil fronto-nasal en: celoides (n=7), ortoides (n=8) y cirtoides (n=4). Todos los especímenes examinados fueron individuos adultos (> 24 meses, juzgados por la erupción total de la serie molar, que debía ser completa) y se encuentran depositados en la colección de la Universidad de Lleida a la que pertenece el autor. Pueden considerarse representativos de cada raza, de acuerdo con el estándar racial de cada una. Puesto que los animales se sacrificaron con fines comerciales ajenos al objeto de este estudio y no fueron objeto de ninguna manipulación ligada a esta investigación, no se consideró pertinente la solicitud de autorización previa de experimentación animal.
Puntos anatómicos de referencia
Se tomaron fotografías individuales, en un plano sagital, por el lado izquierdo, con una cámara Nikon D70®. Los cráneos fueron colocados perpendiculares al plano focal, siempre a la misma distancia de la cámara y las imágenes fueron guardadas en formato .jpg hasta su posterior análisis. Sobre dichas imágenes se definieron una serie de puntos (hitos: puntos del espacio de coordenadas cartesianas (X, Y) y en forma bidimensional, que son las que describen su posición en el espacio).
En un principio se trató de analizar todo el perfil fronto-nasal, pero debido a que no existen referentes claros a lo largo de todo el borde dorsal, y a la pérdida del hueso nasal en algunos cráneos, se optó por añadir un par de semi-hitos -puntos localizados en una curva de acuerdo a la posición de otros hitos o estructuras, y que pueden desplazarse levemente en ella- en el hueso frontal, evitando establecerlos en el hueso nasal.
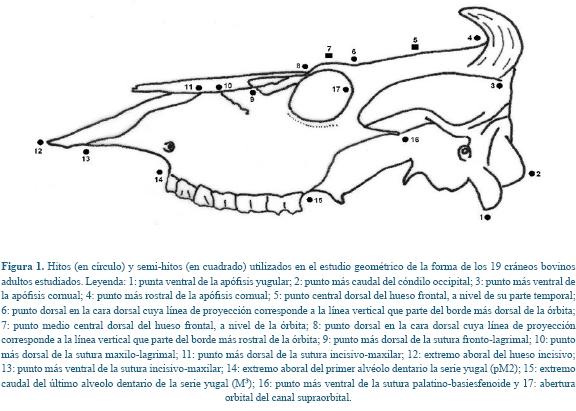
En cada vista lateral del cráneo se definieron finalmente quince hitos y dos semi-hitos que se consideran pueden describir adecuadamente el perfil fronto-nasal y representaron puntos discretos en el cráneo (Figura 1); éstos son descritos como: 1: punta ventral de la apófisis yugular; 2: punto más caudal del cóndilo occipital; 3: punto más ventral de la apófisis cornual; 4: punto más rostral de la apófisis cornual; 5: punto central dorsal del hueso frontal, a nivel de su parte temporal; 6: punto dorsal en la cara dorsal cuya línea de proyección corresponde a la línea vertical que parte del borde más dorsal de la órbita; 7: punto medio central dorsal del hueso frontal, a nivel de la órbita; 8: punto dorsal en la cara dorsal cuya línea de proyección corresponde a la línea vertical que parte del borde más rostral de la órbita; 9: punto más dorsal de la sutura fronto-lagrimal; 10: punto más dorsal de la sutura maxilo-lagrimal; 11: punto más dorsal de la sutura incisivo-maxilar; 12: extremo aboral del hueso incisivo; 13: punto más ventral de la sutura incisivo-maxilar; 14: extremo aboral del primer alvéolo dentario la serie yugal (pM2); 15: extremo caudal del último alveolo dentario de la serie yugal (M3); 16: punto más ventral de la sutura palatino-basiesfenoide y 17: abertura orbital del canal supraorbital. Los hitos y semi-hitos fueron digitalizados usando el programa tpsDig, versión 2.12 24. Se realizó una única réplica.
Estudio morfogeométrico
Como se ha expuesto, la morfometría geométrica se basa en el estudio de la variación en forma de puntos anatómicos de referencias que describen la forma de la estructura analizada, registrados en un sistema de coordenadas X e Y. La forma de una estructura está contenida en las propiedades geométricas que no varían con los cambios de escala, traslación y rotación de la estructura 25. Para todos los especímenes, una vez obtenida la matriz de 34 coordenadas (X, Y) y a fin de eliminar el efecto de la talla, escala y posición se realizó el análisis generalizado de Procrustes 28 mediante el paquete informático CoordGen (disponible en http://www3.canisius.edu/∼sheets).
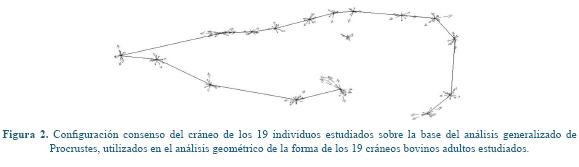
Este procedimiento consiste en la superposición, escalamiento y rotación de todas las conformaciones a un tamaño único. Para esto se siguió el criterio de los mínimos cuadrados, o sea que el ajuste entre las coordenadas de cada uno de los individuos resultase la suma mínima de las distancias al cuadrado entre cada coordenada y la configuración consenso3 (Figura 2). La talla del cráneo fue estimada por el tamaño del centroide, obtenido como la raíz cuadrada de la suma de las distancias al cuadrado de todos los hitos respecto a su centroide25, con el mismo paquete informático CoordGen.
Las diferencias se estudiaron mediante un test ANOVA de una vía, que fue multivariado para el estudio de la forma. Se recurrió también al análisis de componentes principales (ACP) a partir de la matriz de correlación de datos (procrustes) para ver si los clústeres se solapaban o no. Para explorar el posible efecto alométrico -cambios de dimensión relativa de las partes corporales correlacionados con los cambios en el tamaño total, y en este caso concreto, el efecto de la talla del cráneo (inferida por el tamaño del centroide) sobre la forma- se utilizó un análisis de regresión múltiple, utilizando la matriz con las variables de forma como variable dependiente y la tamaño de centroide transformado logarítmicamente como variable independiente 23. Se obtuvo finalmente un fenograma para comprobar las relaciones de similitud (o disimilitud) entre los especímenes estudiados basándose en la morfología craneal. Para ello se utilizó el algoritmo de aglomeración de grupos pareados usando la distancia euclideana (recomendable cuando las variables sean homogéneas y estén medidas en unidades similares). El coeficiente cofenético se usó para medir cuan bien la estructura jerárquica del dendrograma representa a las verdaderas distancias.
Análisis de los datos
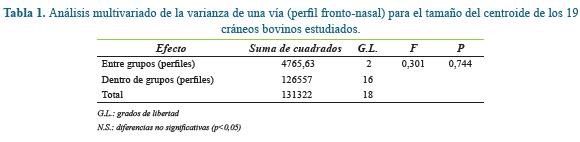
Los análisis estadísticos fueron realizados mediante el programa PAST 12, aplicando los tests que son habituales en los trabajos de morfometría geométrica. No se consideró ni la raza ni el sexo para el tratamiento estadístico, puesto que no era el objeto de este trabajo, centrado únicamente en la comparación de perfiles por métodos de morfometría geométrica. Al presentar el tamaño del centroide una distribución normal (Shapiro-Wilk, W=0,944, P=0,313), se aplicó un test ANOVA (Analysis of Variance, según terminología inglesa) de efecto fijo (Modelo 1) para la comparación de tamaños entre perfiles.
La comparación de formas se realizó mediante un test NPMANOVA (por su nombre en inglés Non Parametric Multivariate Analysis of Variance) utilizando la matriz de distancias, con la corrección p de Bonferroni. NPMANOVA es una extensión del ANOVA para cubrir los casos donde hay más de una variable dependiente que no pueden ser combinadas de manera simple; en el caso de la presente investigación, 17 coordenadas Procrustes X e Y. La corrección de Bonferroni lo que hace es ajustar los resultados estadísticos en comparaciones múltiples.
En nuestra investigación, al probar las hipótesis al nivel p = 0,05, tenemos un 5% de probabilidad de cometer un error de tipo I, es decir, de rechazar falsamente la hipótesis nula, permitiendo contrastar de forma sencilla y sin asunciones adicionales cada una de las hipótesis (nulas) individuales involucradas en el estudio. El nivel de confianza se estableció pues, en todos los casos, en un 5%. Mediante una regresión lineal múltiple se analizó si existía correlación entre tamaño (expresado como el tamaño de centroide transformado logarítmicamente) y la forma (todo el conjunto de coordenadas Procrustes X e Y). Seguidamente, mediante un análisis de componentes principales (en español ACP, en inglés, PCA) se redujo la dimensionalidad de un conjunto de datos.
El ACP se obtuvo a partir de la matriz de correlación de los datos, analizando forma por un lado, y forma y tamaño por el otro. Finalmente, la matriz de distancias euclideanas se utilizó para formar un fenograma mediante el algoritmo del Método de Análisis de Grupos Pareados no ponderados (en inglés Unweighted pair-group average, UPGMA). El coeficiente cofenético se usó para medir cuan bien la estructura jerárquica del dendrograma representaba a las verdaderas distancias.
Resultados y discusión
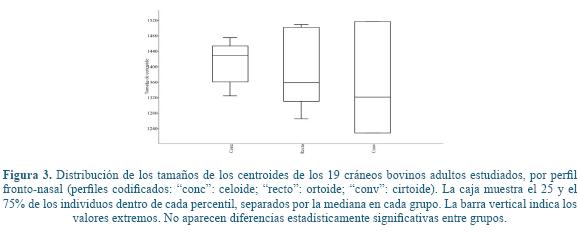
El ANOVA para el tamaño del centroide mostró la ausencia de diferencias significativas en el tamaño craneal (estimado por el centroide) para cada perfil (Tabla 1 y Figura 3). La comparación de los grupos de perfiles en cuanto a su forma mediante un NPMANOVA tampoco mostró diferencias significativas (F=-0,568, P<0,05).
Regresión
No apareció relación significativa alguna entre la forma y el tamaño del cráneo (éste expresado como tamaño del centroide) (R2=0.058); la regresión entre ambas variables explica pues tan solo un 5,8% de la variación, de lo que se deduce que la forma no es dependiente del tamaño, no existiendo pues ninguna relación alométrica -hecho esperable al tratarse, la muestra estudiada, de individuos adultos y, por ende, con el crecimiento finalizado-.
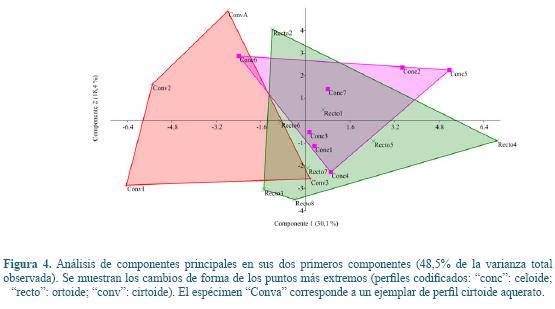
Descripción de los componentes principales (ACP)
En el análisis de la forma, el 48,5% de la varianza fue explicado por los dos primeros CP. El primer eje contenía únicamente el 30,1% de la variación y no separó los 19 individuos en grupos claramente definidos, mostrando una escasa separación por perfiles. El segundo eje, conteniendo sólo 18,4% de la variación total, siguió un notable solapamiento entre los grupos de perfil. La figura 4 muestra la distribución de los individuos en los dos primeros CPs. Aunque en el extremo positivo como en el negativo del CP1 tienden a aparecer los especímenes ortoides, y en el extremo negativo, los cirtoides, el bajo porcentaje de variación explicada por este CP, y la falta de diferencias estadísticamente significativas entre grupos, no permiten asignar con total certeza cada ejemplar a un grupo de perfil concreto, en función de su forma. Analizando forma y tamaño conjuntamente, el CP1 explicó el 29,2% de la variación.
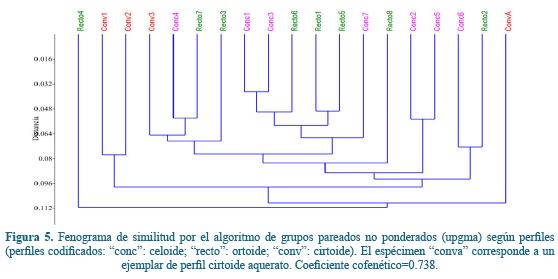
Fenograma
En lo que se refiere al fenograma (Figura 5), los especímenes no muestran ninguna agrupación por perfiles cefálicos. El coeficiente cofenético resultó de 0,738, un valor de bondad o del fenograma que puede considerarse elevado.
La teoría etnológica clásica establece que, a la línea característica del frontal corresponden configuraciones precisas de las arcadas supraciliares u orbitarias en el animal 5. En los perfiles ortoides, estas arcadas se encontrarían en la línea de prolongación del frontal, dando ojos grandes por regla general y a flor de cara, y en los celoides, las órbitas se mostrarían salientes, así como los ojos, correspondiendo al hundimiento de la frente5. En los cirtoides, dichas arcadas se presentarían como borradas o desdibujadas por el pronunciamiento de los huesos craneanos, dando, por consiguiente, ojos escondidos y oblicuos 5. Pero los especímenes estudiados no forman agrupaciones según su perfil fronto-nasal si se atienden los resultados morfogeométricos, de lo que podría deducir que los perfiles, en realidad, no marcan configuraciones generales diferenciadas, ni en forma ni en tamaño. Podría ser, de todos modos, que el no haber podido marcar hitos en el hueso nasal haya reducido la capacidad discriminante de las formas obtenidas. En lo que se refiere a la alometría, se observa que la conformación no cambia con el tamaño. Ahora bien: las diferencias detectadas en los especímenes son sobre animales adultos, de lo que no se puede decir si esta falta de alometría ya está presentes al nacimiento y persiste durante la adultez o bien surge durante la ontogenia posnatal y es el producto de la extensión o acortamiento de esta trayectoria compartida.
Más estudios en esta línea de morfometría geométrica sobre cráneo son necesarios, con más especímenes, con más razas –que podrían incluir razas cebuínas, de perfil fronto-nasal convexo, y razas primitivas- y con piezas óseas completas. Permitiría ello extraer al máximo toda información asociada a la morfología craneal, así como establecer, para posteriores estudios que pudiesen compararse, los hitos con mayor poder discriminante. Los hitos no dejan de ser puntos topográficos elegidos por el investigador a tenor de su hipótesis de trabajo. Será ello posible a medida que se dispongan de colecciones extensas, accesibles y con especímenes de calidad. El establecimiento de colecciones osteológicas de animales domésticos completas y accesibles, como primer paso para ampliar nuevas líneas de investigación utilizando métodos de morfometría geométrica, debería ser un punto de máximo interés en este sentido, y facilitaría sin duda el realizar más trabajos en esta línea.
Conclusiones
Los métodos de morfometría geométrica aplicadas al estudio de los cráneos de animales domésticos permiten la captura de mayor información morfométrica que los métodos lineales clásicos, puesto que disponemos de información tanto del tamaño como de la forma, y por separado.
Para los 19 especímenes bovinos adultos estudiados, no aparecen diferencias ni de forma ni de tamaño entre los grupos de acuerdo a su perfil fronto-nasal, aunque la mayor proporción de la variabilidad en los datos morfométricos se debe al tamaño de la variación entre los individuos.
La variación en forma no se asocia al tamaño de los cráneos.
La apreciación aloídica en los bovinos tendría un sentido más en el plano etnológico (apreciación racial) que en el anátomo funcional.
Referencias
1. Agüera E, Massa R. Introducción a la topografía craneoencefálica en el perro basada en métodos radiográficos. Arch. Zoot. 26(101):9-21. 1977. [ Links ]
2. Agüera S, Robina A, Vásquez JM, Rodríguez I. Anatomía del craneoencéfalo del ovino (raza Merina) y sus fundamentos topográficos para una estereotaxis aplicada. Arch. Zoot. 32 (124):255-277. 1983. [ Links ]
3. Agüera S, Castejón FJ, Díez A, Miró F, López-Rivero JL. Diferenciación racial de la oveja Manchega y Merina mediante valoración de parámetros radiográficos cefálicos. Arch. Zoot. 37(139):205-216. 1988. [ Links ]
4. Agüera S, Rodríguez V, Miró F, Vivo R. Parámetros cefálicos radiográficos de la oveja Merina Segureña y su aplicación racial. Arch. Zoot. 37(138):115-129. 1988. [ Links ]
5. Aparicio G. Zootecnia especial. Etnología compendiada. Imp. Moderna. Córdoba. 465 pp. 1960. [ Links ]
6. Bookstein FL. Morphometric tools for landmark data: Geometry and Biology. Cambridge University Press, New York. 435 pp. 1991. [ Links ]
7. Bornholdt R, Oliveira LR, Fabian ME. Size and shape variability in the skull of Myotis nigricans (Schinz, 1821) (Chiroptera: Vespertilionidae) from two geographic areas in Brazil. Braz. J. Biol. 68:623-631. 2008. [ Links ]
8. Bruenner H, Lugon-Moulin, N, Balloux F, Fumagalli L, Hausser JA. Taxonomical re-evaluation of the valais chromosome race of the common shrew Sorex araneus (Insectivora: Soricidae). Acta Theriogenol. 47:245-275. 2002. [ Links ]
9. D'anatro A, Lessa EP. Geometric morphometric analysis of geographic variation in the Río Negro tuco-tuco, Ctenomys rionegrensis (Rodentia: Ctenomyidae). Mam. Biol. 71:288-298. 2006. [ Links ]
10. Dryden IL, Mardia KV. Statistical shape analysis. John Wiley and Sons, New York. 347 pp. 1998. [ Links ]
11. Gould SJ. Allometry and size in ontogeny and phylogeny. Biol. Rev. 41:587-640. 1966. [ Links ]
12. Hammer Ø.; Harper DAT, Ryan PD. Past. Palaeontol. Stat. 4(1):1-9. On Line: http://nhm2.uio.no/norlex/past/download.html. 2010. Fecha de consulta: 23 de Abril de 2010. [ Links ]
13. Heberer G, Herre W, Rensch B, Roehrs M, Schwanitz F. Die Evolution der Organismen, Domestikation und Stammesgeshichte. Gustav Fischer Verlag. Stuttgart. Pp 29-174. 1971. [ Links ]
14. Herre W, Roehrs M. Haustiere-zoologisch gesehen, Gustav Fischer Verlag. Stuttgart. 412 pp. [ Links ] 1990.
15. Kobrynczu KF, Krasinka M. Skulls of European bison and cattle hybrids. Acta Theriogenol. 32:11-20. [ Links ] 1987.
16. Komosa M, Molinski K, Godynicki S. The Variability of Cranial Morphology in Modern Horses. Zool. Sci. 23:289-298. 2006.
17. Lissovsky AA. Geographical variation of skull characters in pikas (Ochotona, Lagomorpha) of the alpine-hyperborea group. Acta Theriogenol. 48:11-24. 2003. [ Links ]
18. O'higgins P, Jones N. Tools for statistical shape analysis. Hull York Medical School. On Line: http://sites.google.com/site/hymsfme/resources. 2006. Fecha de consulta: 23 de Abril de 2010. [ Links ]
19. Radinsky LB. Evolution of skull shape in carnivores. 1. Representative modern carnivores. Biol. J. Linn. Soc. Land 15:369-388. 1981. [ Links ]
20. Ritke ME, Kennedy ML. Geographic Variation of Sexual Dimorphism in the Raccoon Procyon lotor. Am. Midland Nat. 129:257-265. 1993. [ Links ]
21. Robina A. Topografía craneoencefálico del vacuno de raza Frisona, por métodos radiológicos. Arch. Zoot. 31(119):51-72. 1982. [ Links ]
22. Rohlf FJ. Bias and error in estimates of mean shape in geometric Morphometrics. J. Hum. Evol. 44:665-683. 2003. [ Links ]
23. Rohlf FJ. TpsRegr 1.34. Ecology and Evolution, State University of New York at Stony Brook. http://life.bio.sunysb.edu/morph/index.html. 2007. Fecha de consulta: 23 de Abril de 2010. [ Links ]
24. Rohlf FJ. TpsDig 2.12. Ecology and Evolution, State University of New York at Stony Brook. State University of New York at Stony Brook. http://life.bio.sunysb.edu/morph/index.html. 2008. Fecha de consulta: 23 de Abril de 2010.
25. Rohlf FJ, Bookstein FL. Proceedings of the Michigan Morphometrics Workshop. The University of Michigan Museum of Zoology. Michigan Special. Publication No. 2. 380 pp. 1990. [ Links ]
26. Rohlf FJ, Marcus LF. A revolution in morphometrics. Trends Ecol. Evol. 8:129-132. 1993. [ Links ]
27. Small CG. The statistical theory of shape. Springer, New York. 227 pp. 1996. [ Links ]
28. Zeldtich ML, Swiderski DL, Sheets HD, Fink WL. Geometric morphometrics for biologists: a primer. Elsevier Academic Press, San Diego, California. 443 pp. 2004. [ Links ]
