










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCES Medicina Veterinaria y Zootecnia
On-line version ISSN 1900-9607
Ces. Med. Vet. Zootec. vol.11 no.1 Medellín Jan./June 2016
Artículos originales de investigación
Polymorphisms of the Dopamine beta-hidroxylase gene and its association with bovine temperament¤
Polimorfismos del gen Dopamina beta hidroxilasa y su asociación con temperamento bovino
Polimorfismos no gene dopamina beta hidroxilase e sua associação com temperamento bovino
Julia Victoria Arredondo Botero1*, Zoot, PhD; Germán Darío Saavedra Correa1, MSc, PhD (c).
¤ Para citar este artículo: Arredondo Botero JV; Saavedra Correa GD. Polimorfismos del gen Dopamina beta hidroxilasa y su asociación con temperamento bovino. Rev. CES Med. Zootec. 2016; Vol 11 (1): 51-61.
* Autor para correspondencia: Julia Victoria Arredondo Botero. Facultad de ciencias pecuarias. Corporación universitaria Santa Rosa de Cabal, Kilometro 4 Vía Santa Rosa de Cabal - Chinchiná. Santa Rosa de Cabal - Risaralda - Colombia. Email: jarredondob@unal.edu.co
1 Corporación universitaria Santa Rosa de Cabal, Facultad de Ciencias pecuarias, Carrera Zootecnia, Grupo de investigación en reproducción y mejoramiento genético. Kilometro 4 Vía Santa Rosa de Cabal - Chinchiná. Santa Rosa de Cabal - Risaralda - Colombia.
(Recibido: 8 de agosto, 2015; aceptado: 12 de enero, 2016)
Abstract
Temperament in bovines is defined as behavior in response to human handling and is influenced by the sympatheticadrenal system. The bovine Dopamine beta-hidroxylase (DBH) gene is located on chromosome 11 and codes for the Dopamine beta hidroxylase enzyme, essential por the synthesis of this neurotransmitter. Simple nucleotide polymorphisms (SNP) have been identified on exon 12, possibly associated with changes in bovine temperament. The aim of this study was to find polymorphisms in this region and associate them with temperament traits in BON (Blanco Orejinegro - Bos taurus taurus), fighting bulls (Bos taurus taurus) and Brahman (Bos taurus indicus) cattle breeds. A total of 80 individuals were used to evaluate chute behavior (CB) and flight speed (VS), and in 52 of them a 702bp fragment in exon 12 of the gene was sequenced, identifying 22 haplotypes. The mínimum number of haplotypes (4), the lowest nucleotide diversity (0,0012±0,0011) and the highest fixed loci number were found on fighting bulls. Moderate differentiation between the populations was observed with the lowest genetic distance (FST=0.03309) between Bos taurus taurus breeds. Differences in VS were observed (p<0.05) being lower in BON males than in Brahman (1.45±0.61 and 2.02±0.74 respectively) and in Brahman females than in Fighting bull females (1.05±0.58 y 2.23±0.80 respectively). Even though no association with the genetic plymorphisms was found, a posible influence of selection in exon 12 is suggested which may influence in other behavioral characteristics.
Key words: Chute behavior, flight speed, genetic association, haplotype..
Resumen
El temperamento en bovinos se define como el comportamiento en respuesta a la manipulación humana y está influenciado por el sistema simpático adrenal. El gen DBH (Dopamina beta hidroxilasa) bovino está ubicado en el cromosoma 11 y codifica para la Dopamina beta hidroxilasa (DBH), enzima indispensable para la síntesis de este neurotransmisor. Se han identificado polimorfismos de nucleótido simple en el exón 12, posiblemente asociados a cambios en el temperamento en bovinos. El objetivo de este estudio fue encontrar polimorfismos en dicha región y asociarlos con rasgos de temperamento en las razas Blanco orejinegro (BON - Bos taurus taurus), de lidia (Bos taurus taurus) y brahman (Bos taurus indicus). Se utilizaron 80 individuos para evaluar comportamiento en brete (CB) y velocidad de salida (VS), y en 52 de ellos se secuenció un fragmento de 702 pb en el exón 12 del gen identificando 22 haplotipos. El menor número de haplotipos (4), la menor diversidad nucleotídica (0,0012±0,0011) y el mayor número de loci fijos se presentaron en bovinos de Lidia. Hubo moderada diferenciación entre las poblaciones con la menor distancia genética (FST=0,03309) presente entre las razas Bos taurus. Se presentaron diferencias (p<0,05) en VS, siendo ésta menor en machos BON que en Brahman (1,45±0,61 y 2,02±0,74 respectivamente) y en hembras Brahman que en hembras de Lidia (1,05±0,58 y 2,23±0,80 respectivamente). Aunque no se presentó asociación con los polimorfismos genéticos encontrados, se sugiere una posible influencia de fuerzas de selección en el exón 12 que podría influir en otras características comportamentales.
Palabras clave:Asociación genética, comportamiento en brete, haplotipo, velocidad de salida.
Resumo
O temperamento em bovinos se define como o comportamento em resposta a manipulação humana e está influenciado pelo sistema simpático adrenal. O gene DBH bovino está localizado no cromossomo 11 e codifica para a dopamina beta hidroxilase (DBH), enzima indispensável para a síntese de este neurotransmissor. Tem se identificado polimorfismos de nucleotídeo simples no éxon 12, possivelmente associados a mudanças no temperamento em bovinos. O objetivo de este estudo foi buscar possíveis polimorfismos no éxon 12 do gene DBH e avaliar sua associação com características de temperamento nas raças Blanco Orejinegro (BON - Bos taurus taurus), de Lídia (Bos taurus taurus e zebu Brahman (Bos taurus indicus). Se utilizaram 80 indivíduos para avaliar o comportamento no curral (CB) e velocidade de saída (VS). Em 52 deles sequenciou-se um fragmento de 702 pb no éxon 12 do gene DBH identificando 22 haplotipos. O menor número de haplótipo (4), a menor diversidade nucleotídica (0,0012±0,0011) e o maior número de loci fixos apresentaram-se em bovinos de Lídia. Houve moderada diferenciação entre as populações com a menor distancia genética (FST=0,03309) presente entre as raças Bos Taurus. Apresentaram-se diferenças (p<0,05) em VS, sendo esta menor em machos BON que em Brahman (1,45±0,61 e 2,02±0,74 respectivamente) e em fêmeas Brahman que de Lídia (1,05±0,58 y 2,23±0,80 respectivamente). Ainda que não se apresentou associação com os polimorfismos genéticos encontrados, sugere-se uma possível influência de forças de seleção no éxon 12 que poderia influenciar em outras características comportamentais..
Palavras-chave:Associação genética, comportamento no curral, haplótipo, velocidade de saída.
Introducción
El temperamento en bovinos se define como la respuesta de comportamiento animal a la manipulación por humanos5 o en respuesta al estrés 24; es un carácter heredable, que impacta sobre el bienestar y la producción bovina y puede utilizarse como criterio de selección 30.
El comportamiento del animal afecta directamente su productividad, al modificar el balance general y gasto energético diario 16 e influye en el desarrollo, la salud 21, el hematocrito y el pH de la carne 9.
La docilidad podría ser potencialmente usada como un indicador de características de importancia económica que son difíciles de medir 21, con una heredabilidad moderada11, por lo que se puede incluir el temperamento como un objetivo global del mejoramiento13. La selección de ganado por temperamento maximiza la eficiencia productiva reflejada en la ganancia de peso en los corrales de engorde32. Sin embargo, el ganado de lidia ha sido empíricamente seleccionado por características comportamentales, especialmente aquellas relacionadas con la agresividad 27, fundamental para el desarrollo del festejo taurino y la percepción por parte del público del riesgo inherente a él 12.
Existen varias formas de medir el temperamento, dos de las más usadas son el puntaje en la manga de compresión o comportamiento en el brete (CB), que muestra el comportamiento del animal mientras es confinado en un brete, y la velocidad de salida (VS), definida como la velocidad (m/s) a la cual el animal atraviesa una distancia específica después de salir de la manga de compresión 21. Bovinos temperamentales, tienen VS mayor a 2,4 m/s, mientras que los calmados, tienen valores inferiores a 1,9 m/s 5.
Bovinos más excitables tienen mayores concentraciones basales de glucocorticoides y catecolaminas 4.
La noradrenalina es el principal precursor de la adrenalina, y se sintetiza a partir de la dopamina (DBH) 16. Es una catecolamina implicada en la respuesta al estrés, regulación del tono del músculo liso, presión arterial, metabolismo de los hidratos de carbono y lípidos, termorregulación, secreción de diversas hormonas y numerosos procesos centrales y periféricos 3.
Su síntesis es catalizada por la enzima Dopamina Beta Hidroxilasa, codificada en el gen DBH bovino, el cual se encuentra ubicado en el cromosoma 11, posee 12 exones y una longitud de 17,79 Kpb, (NCBI Gene ID: 280758) 20. En bovinos, se han encontrado 62 polimorfismos y se ha hallado asociación entre polimorfismos del exón 12 y rasgos de temperamento, como un comportamiento en la manga más calmado (p=0,0326) en bovinos brahman de genotipo AA (NCBI SNP ID rs109805094) y la tendencia a tener un comportamiento más calmado en la manga y un mejor comportamiento a la salida, los individuos homocigotos GG (NCBI SNP ID rs110864292 y rs110796564, respectivamente) 16.
El objetivo de este trabajo fue evaluar los polimorfismos presentes en el exón 12 del gen DBH bovino, en razas de las subespecies Bos taurus taurus (Blanco Orejinegro- BON y bovinos de Lidia) y Bos taurus indicus (Brahman), y establecer su potencial asociación con rasgos de temperamento, medidos en variables como VS y CB.
Materiales y métodos
Animales
Para el análisis de temperamento, se utilizó un total de 80 individuos, de las razas BON, de Lidia, Brahman élite, es decir, de alto valor genético (e) y Brahman de tipo comercial (c), de los cuales se seleccionaron 49 de los individuos con temperamento más contrastante para el análisis molecular (Tabla 1).
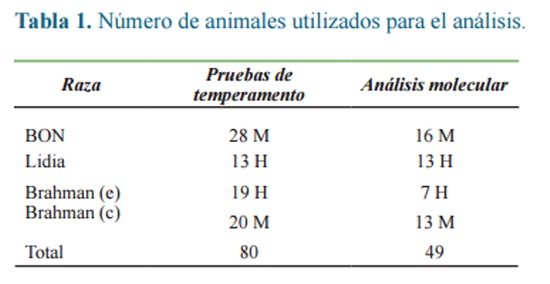
Pruebas de temperamento
Para evaluar la VS se registró en videocámara el comportamiento del animal durante los primeros 30 segundos después de ingresar a la báscula. Posteriormente se determinó la velocidad de salida, establecida como la tasa (m/s) a la cual el animal recorría 1,83 metros después de salir de la báscula, clasificándolos como temperamentales o nerviosos cuando recorren >2,4m/s y calmados cuando recorren 1≤,9m/s 5.
El CB fue evaluado independientemente por cinco personas con la siguiente escala: 1- Calmado, sin movimientos, 2- Movimientos incesantes, 3- Torsiones y sacudidas ocasionales y no vigorosas de la báscula, 4- Movimiento vigoroso y sacudimiento continuo del dispositivo, 5- Corcovos, giros o lucha violenta 32, posteriormente se asignó a cada animal un puntaje promedio producto de las cinco evaluaciones.
Muestras de sangre
Una vez realizada la prueba de temperamento, los animales fueron reingresados a la báscula, y se les extrajeron 5 mL de sangre de la región coccígea en tubos con EDTA. Se extrajo ADN mediante el protocolo Salting Out [19]. La calidad y concentración del ADN se evaluó en geles de agarosa al 0,8% en TAE al 1% (Tris, Acetato, EDTA, pH 8,0), utilizando bromuro de etidio como agente intercalante.
Condiciones de amplificación y secuenciación
Para la amplificación del exón 12 del gen DBH, se utilizaron los siguientes cebadores: Forward: 5' TTTGGTTTGGGAGAAGG 3' y Reverse:
5' TGCACCCGTCACACTCAA 3' 16. Las condiciones para la amplificación fueron 2mM MgCl2, 25 µM dNTPs, 0,2 µM de cada cebador, 20 ng de ADN genómico y 0,1 U Taq DNA polimerasa (Bioline). El perfil térmico de la PCR fue: incubación a 95 °C por 5 min (95 °C por 45 s, 62 °C por 45 s, disminución de 2°C cada ciclo, 72 °C por 45 s) con repetición 5 veces; 95 °C por 45 s, 55 °C por 45 s, 72 °C por 45s y repetición por 30 ciclos e incubación a 72 °C por 10 min. Los fragmentos amplificados fueron enviados al servicio de secuenciación de Macrogen Sequenciation Service, en Maryland EEUU.
Análisis estadístico
La evaluación del temperamento se realizó mediante el procedimiento de Modelos Lineales Generales (GLM) del paquete estadístico SAS ver 9.0. (SAS Institute Inc., Cary, NC, USA), según el modelo: Yij = µ + Si + Rj + εij, donde Yij es el valor observado, Si es el efecto del sexo i, Rj es el efecto de la raza j y εij es el error aleatorio y se usó el test de rangos múltiples de Duncan, con un nivel de significancia α=0,05. Para la asociación entre rasgos de temperamento y genotípicos, se utilizó el mismo paquete estadístico, utilizando la corrección de Bonferroni para comparaciones múltiples.
Para el análisis genético, las secuencias obtenidas fueron editadas mediante el software GeneStudio ⢠Professional Edition Ver 2.2.0.0. y alineadas con el programa Mega 6 28, tomando como referencia la secuencia con el número de accesión GCA_000003055.5 (GenBank), perteneciente a Bos taurus. El cálculo de las frecuencias haplotípicas, índices de diversidad molecular, distancia genética y AMOVA se llevó a cabo con el software Arlequín Ver. 3.5.1.2. 10 y las frecuencias genotípicas, alélicas y las desviaciones del equilibrio de Hardy-Weinberg (EHW) para los sitios polimórficos, con el software GENALEX ver 6.5 22
Resultados
Evaluación del temperamento
La comparación entre machos y hembras solo fue posible dentro de la raza Brahman, debido a que fue la única raza en la que se pudo realizar muestreo para ambos sexos, encontrando que los machos son más temperamentales (p<0,05) que las hembras para el rasgo comportamiento en el brete (Tabla 2).
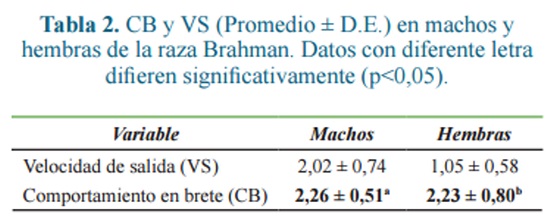
CB no mostró diferencias entre todas las razas evaluadas (Tabla 3), sin embargo, VS fue superior (p<0,05) en hembras de Lidia, frente a hembras Brahman élite y mayor en machos Brahman comercial que en machos de la raza criolla colombiana BON.
En la población general, no se encontraron diferencias en el CB, sin embargo, la VS fue superior (p<0,05) en bovinos de Lidia y Brahman (c), con valores de 2,23 ± 0,80 y 2,02 ± 0,74, respectivamente, frente a bovinos de las razas BON y Brahman (e), con valores de 1,45 ± 0,61 y 1,05 ± 0,58, respectivamente.
Análisis de polimorfismos
El alineamiento de las secuencias consenso con el SNP rs109805094, reportado previamente como posible indicador de temperamento en bovinos, mostró en todos los individuos el genotipo GG.
Según el análisis de secuencias, de los haplotipos encontrados, el haplotipo 2 fue el más frecuente en toda la población, con una frecuencia de 0,81 en bovinos de Lidia, 0,56 en BON y 0,20 en Brahman.
En BON se encontraron 8 haplotipos, con 10 sitios polimórficos (9 transiciones y 1 transversión), y 7 individuos presentaron haplotipos únicos con frecuencias del 6%. En Brahman se encontraron 13 haplotipos, con 17 sitios polimórficos (14 transiciones, 2 transversiones y 1 indel-inserciones/deleciones). Después del segundo haplotipo, los haplotipos 12 y 16, se encontraron en 3 individuos cada uno y los demás haplotipos presentaron frecuencias inferiores al 5%. En ganado de Lidia, se encontraron 4 haplotipos, con 5 sitios polimórficos todos correspondientes a transiciones; además del segundo haplotipo, se presentó el haplotipo 12 compartido con Brahman y dos haplotipos únicos, el 21 y 22, con frecuencias del 6% de la población.
En la tabla 4 se presentan los haplotipos encontrados y los sitios polimórficos que los caracterizan. Solo fueron considerados los haplotipos con frecuencias superiores al 5%, los demás son tomados como haplotipos raros.
El mayor valor de diversidad nucleotídica Ï (heterocigosis esperada a nivel nucleotídico o número promedio de diferencias por sitio entre secuencias tomadas a pares) se presentó en la raza brahman con 0,0120 ± 0,0070, seguido por la raza BON con 0,0081 ± 0,0050, y el ganado de lidia presentó el menor valor con 0,0012 ± 0,0011.
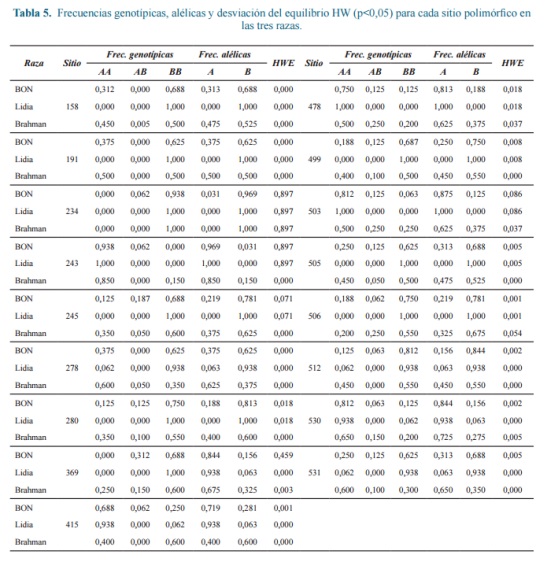
Con respecto a los sitios polimórficos, solo uno presentó equilibrio HW en las tres poblaciones (Tabla 5). En todos
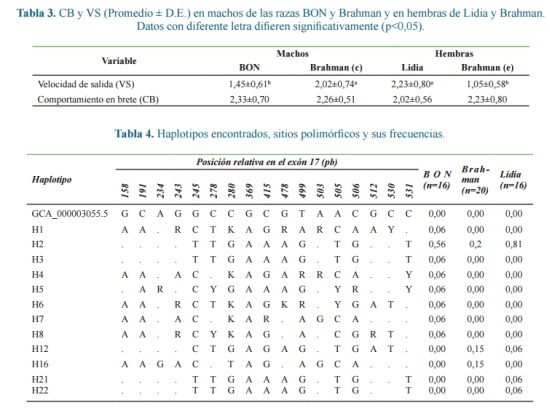
los individuos brahman 15 loci presentaron desviación significativa (p<0,05) a favor de los homocigotos, en bovinos de lidia se presentaron 13 y en BON 12 sitios, lo que podría sugerir, entre otras causas, que en todas las poblaciones se han llevado a cabo procesos de selección lo que indica que en todas las poblaciones se han llevado a cabo procesos de selección que involucran este exón. El mayor número de loci con genotipos fijos homocigotos se presentó en los bovinos de lidia, éste déficit de heterocigotos puede ser consecuencia de procesos de endogamia o de selección a favor de los homocigotos.
Estructura y diferenciación genética de las poblaciones
El AMOVA (Tabla 6), mostró mayor diferenciación entre individuos dentro de las razas, que entre razas. El estadístico FST para la población total fue moderado, con un valor de 0,1205 (p<0,05). Se encontraron valores de FST entre 0,03 y 0,21 (Tabla 7), con baja diferenciación entre BON y lidia, moderada entre BON y brahman, y la mayor distancia (p<0,05), se dio entre brahman y los bovinos de lidia, lo que indica que son las poblaciones más alejadas.
Asociación entre polimorfismos y temperamento
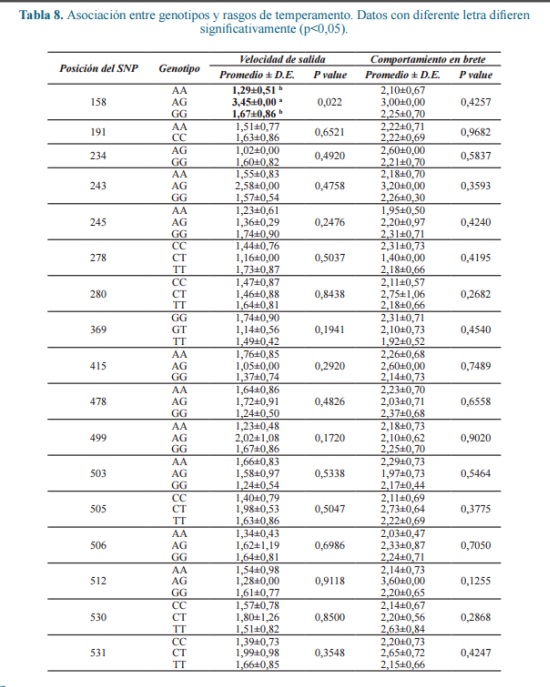
El análisis se realizó teniendo en cuenta que la frecuencia del alelo menor debe ser al menos del 5% para evitar resultados falsos en un estudio asociativo entre genotipo y fenotipo 2. De los 17 polimorfismos encontrados (Tabla 8), solo uno, ubicado en la posición 158 del exón, mostró efecto significativo, con una incremento en la VS de los heterocigotos, aunque después de la corrección de Bonferroni, este no fue significativo.
Al relacionar rasgos de temperamento con haplotipos, la VS no presentó diferencias significativas entre los diferentes haplotipos, sin embargo, el CB tuvo valor significativamente mayor (p>0,05) en el haplotipo 8, presente en bovinos de la raza BON, con un valor de 3,6 y el menor valor (p>0,05) se presentó en el haplotipo 9, presente en individuos brahman, con un valor de 1,2. Los demás haplotipos presentaron valores intermedios.

Discusión
El ganado con influencia Bos índicus tiene un temperamento más excitable comparado con el ganado Bos taurus 32. Esto se evidenció al comparar VS entre machos brahman, frente a BON. Sin embargo, las hembras de lidia presentaron mayor VS que las brahman, pues ésta ha sido una raza seleccionada típicamente por rasgos temperamentales.
En la población total, CB no mostró diferencias entre razas. Fue significativamente superior para machos que para hembras, con valores de 2,26 ± 0,51 y 2,23 ± 0,80 respectivamente. Esta medición sólo mostró diferencias significativas cuando se midió entre machos y hembras de la raza brahman, siendo más temperamentales los primeros. Sin embargo VS sí presentó diferencias significativas entre razas.
Se ha evaluado el temperamento en bovinos brahman y angus, encontrando que las hembras presentan 6 numéricamente mayor puntuación de temperamento que los machos, aunque sin efecto del sexo en ambas razas (pâ¥0,09). En bovinos mestizos de carne, la medición de temperamento no se ve afectada por factores como la edad al destete; sin embargo, el sexo influye en mediciones como el puntaje en el corral, presentando mayores valores las hembras frente a los machos adultos, con valores de 3,69 ± 0,13 y 3,19 ± 0,21 31.
Se ha reportado un menor valor de CB para la raza nativa brasilera Caracú, perteneciente a la especie Bos taurus (1,52 ± 0,83) frente a razas cebuínas, como Gyr, Guzerat y Nellore, con 2,48±1,22; 2,59±1,33 y 2,20±1,21, respectivamente, siendo Gyr y Guzerat las de mayor temperamento (p<0,05) y valores de VS superiores (p<0,05) para Caracú y Nellore (2,52±1,21 y 2,14±1,18), frente a las razas Gyr y Guzerat (1,51±0,97 y 1,64±1,18), respectivamente 23.
La VS en bovinos Brahman (2,02±0,74), fue inferior a lo reportado para bovinos Nellore (2,26±1,00) 26 y superior a lo reportado para bovinos Brahman élite (1,184 ± 0,39) 16.
A pesar de la marcada influencia de la raza encontrada en este estudio para la VS, se ha encontrado que durante la manipulación, bovinos con temperamento excitable presentan mayor nivel de cortisol plasmático vs bovinos de temperamento adecuado, independientemente del tipo de raza (B. indicus à B. taurus, p<0,01; B. taurus, p<0,01; B. indicus, p=0,04) o edad (vacas, p<0,01; novillas o novillos, p<0,01) 8.
En este estudio se presentó una baja correlación entre VS y CB (0,0493; p>0,05); otros autores han encontrado mayores valores de correlación, aunque también bajos (0,26561; p<0,0001) 14 e incluso valores negativos (-0,36; p< 0,01) 23.
Una respuesta de comportamiento negativa a la manipulación y procedimientos de manejo, puede afectar la productividad 4. La VS es el mejor predictor de los efectos del temperamento en las características de la canal y la tasa de crecimiento 33. Se han reportado además polimorfismos de nucleótido simple asociados a ganancia diaria de peso y consumo promedio diario de alimento, en el cromosoma bovino BT6, asociados a VS 15.
Con respecto al gen DBH bovino, en la actualidad se cuenta con pocos estudios. Se ha secuenciado el 97% del DNAc y el 13% de la secuencia no codificante, encontrando 62 cambios nuleotídicos, con una frecuencia de un SNP cada 85 pb 17. Específicamente en el exón 12 del gen, se ha encontrado una asociación significativa (p=0,0326) entre el polimorfismo A/G (17269 A>G) y un buen comportamiento en el brete para los individuos AA, así como la tendencia de los genotipos GG del polimorfismo A/G (17299 A>G), a ser más calmados para la misma medición y los genotipos GG del polimorfismo G/A (17300 G>A) a tener mejor temperamento a la salida 16.
Los valores de diversidad nucleotídica Ï son altos, si se tiene en cuenta que en razas criollas colombianas se han reportado valores de 0,0056 ± 0,003 1, evaluados en regiones mitocondriales, mientras que el valor encontrado en este estudio para la raza criolla BON, fue de 0,008144 ± 0,004767, en un gen nuclear, con menor tasa de mutación.
En los bovinos de lidia hubo menor diversidad nucleotídica, desviaciones significativas del EHW y el mayor número de loci con genotipos fijos homocigotos, lo que demuestra que en este exón pueden incidir fuerzas como selección contra heterocigotos, subdivisión de la población (efecto Wahlund) o consanguinidad 18, además son poblaciones que tradicionalmente utilizan un reducido número de machos reproductores, con aislamiento reproductivo entre encastes y un reducido tamaño efectivo consiguiente 7. Esto se evidencia también con el valor moderado de FST, que indica estructuración genética en las poblaciones analizadas, y una mayor diferenciación genética entre las razas Brahman y Lidia.
Las ganaderías de lidia son empresas que dependen de los espectáculos taurinos y su diferente estructura genética y tipo de selección, la hacen difícilmente intercambiable con cualquier otra raza 29.
A pesar de que en otro estudio en novillos Brahman élite se evaluó el SNP rs109805094, encontrando asociación significativa (p=0,0326) entre el genotipo AA y un comportamiento calmado en brete [16], para ese SNP, todos los individuos analizados en este estudio, presentaron un genotipo que podría ser clasificado como de temperamento bravo; es posible que dicho polimorfismo no se haya fijado en las poblaciones evaluadas.
Se ha concluido en otros estudios que el CB es una evaluación subjetiva, que puede variar entre individuos debido a sus diferencias en la observación del animal y su criterio de interpretación para la puntuación 14. Las pruebas de campo llevadas a cabo en este estudio pudieron presentar algún grado de subjetividad. Existe una variedad de parámetros de comportamiento, fisiológicos, bioquímicos, inmunológicos y patológicos que han sido propuestos para evaluar la capacidad de respuesta de los animales ante el estrés agudo 25 y podrían reforzar las mediciones en campo..
Conclusiones
Las pruebas en campo demostraron diferencias entre las dos mediciones realizadas, pudiendo ser más subjetivo el CB. Las VS mostraron resultados similares a los reportados en razas Bos indicus y Bos taurus, con excepción de los bovinos de lidia, que han sido seleccionados a favor de un temperamento agresivo.
Los resultados encontrados sugieren que en el exón estudiado podrían influir fuerzas de selección. Según el polimorfismo reportado previamente, todos los individuos tendrían un genotipo que determina un temperamento agresivo, sin embargo, las poblaciones evaluadas no presentan variaciones en este sitio.
Se detectaron 17 sitios polimórficos en el exón, pero no hubo una clara asociación entre un genotipo determinado y los rasgos de temperamento medidos en campo. No se descarta su influencia en otras características temperamentales, por lo que se sugiere la pertinencia de otro tipo de mediciones fenotípicas, incluyendo además la cuantificación plasmática de los niveles de noradrenalina.
Agradecimientos
A Fernando Morales Vallecilla Zoot, Esp, PhD (C) y a los productores Darío Sanint, Jaime Escobar y Juan Camilo Palomino. Al laboratorio de Biología molecular de la Corporación Universitaria Santa Rosa de Cabal. A Juliana Andrea Cuetia, MSc. y Darwin Yovanny Hernández, MSc. PhD. A los estudiantes Yency Neira, Guillermo Castro, Nathaly Valencia, Marcela Santamaría y Claudia Restrepo.
Referencias
1.álvarez L, Martínez A, Delgado J, Vinasco A, Muñoz J, Hernández D. Diversidad del ADN mitocondrial en ganado criollo colombiano. Actas Iberoamericanas de Conservación Animal, AICA 2014; 4(1): 19-21. [ Links ]
2. Balding D. A tutorial on statistical methods for population association studies.. Nat Reviews 2006; 7(10): 781-791. [ Links ]
3. Brandan N, Llanos I, Ruiz D, Rodríguez A. Hormonas Catecolamínicas Adrenales. Cátedra de Bioquímica Facultad de Medicina. [acceso: 4 de abril de 2015]. URL:http://med.unne.edu.ar/sitio/multimedia/imagenes/ckfinder/files/files/Carrera-Medicina/BIOQUIMICA/catecolaminas.pdf. 2010. Argentina: Universidad Nacional del Nordeste. [ Links ]
4. Burdick N, Randel R, Carroll J, Welsh T. (2011). Interactions between Temperament, Stress, and Immune Function in Cattle. International Journal of Zoology. 2011. Article ID 373197, 9. [ Links ]
5. Burrow H. Effect of intensive handling of zebu crossbred weaner calves on temperament,"Proceedings of the Australian Association. Animal Breeding and Genetics, 1991 vol. 9, 208-211. [ Links ]
6. Cafe L, Robinson D, Ferguson D, McIntyre B, Geesink G, Greenwood P. Cattle temperament: Persistence of assessments and associations with productivity, efficiency, carcass and meat quality traits. J. Anim. Sci. 2011; 89(5): 1452-1465. [ Links ]
7. Cañon J, Garcia-Atance M, Cortés O, García D, Fernández J. Genetic variation within the Lidia bovine breed. Anim. Genet. 2008; 39(4): 439-445. [ Links ]
8. Cooke R. Temperament and acclimation to human handling influence growth, health, and reproductive responses in Bos taurus and Bos indicus cattle. Kunkle interdisciplinary beef symposium. [acceso: 26 de Noviembre de 2014]. Oregon, USA: www. journalofanimalscience.org at State University Lib. [ Links ]
9. Davies P, Pighin D, Ceconi I, Pazos A, Méndez D, Buffarini M, et al. Efecto del temperamento y manejo prefaena sobre parámetros bioquímicos y calidad de carne en bovinos. Primer encuentro de investigadores en bienestar animal. Valdivia. 2009. [acceso: 1 de abril de 2015]. http://inta.gob.ar/sites/default/files/script-tmpmt2008_davies_efecto_del_temperamento.pdf [ Links ]
10. Excoffier L. Arlequin ver 3.5.1.3. CMPG. University of Berne. 2009. [ Links ]
11. Ferguson D. Less fearful livestock will improve welfare and Production. Livestock Horizons. 2006; 2 (1): 21-22. [ Links ]
12. Gil C. Variables neuroendocrinas y su relación con el comportamiento durante la lidia del toro bravo ("Bos taurus", l.). Tesis doctoral. Universidad complutense de Madrid. 2013. 197 p. [ Links ]
13. Hoppe S, Brandt H, König S, Erhardt G, Gauly M. Temperament traits of beef calves measured under field conditions and their relationships to performance. J Anim Sci. 2010; 88(6): 1982-1989. [ Links ]
14. Jones T. Measurement of Temperament in Beef Cattle and its Relationships to Animal Production Characteristics. Tesis de maestría. Population Medicine. Guelph, Ontario, Canadá: University of Guelph. 2013. 87 p. [ Links ]
15. Lindholm-Perry A, Kuehn L, Freetly H, Snelling W. Genetic markers that influence feed efficiency phenotypes also affect cattle temperament as measured by flight speed. Animal Genetics. 2015; 46 (1):60-64. [ Links ]
16. Lourenco D. Análisis funcional del gen dopamina beta hidroxilasa (DBH) y su asociación a temperamento en bovinos. Tesis doctoral. México: Instituto politécnico nacional. 2013. 84 p. [ Links ]
17. Lourenco D, Sifuentes A, Parra G, De la Rosa X, Segura A, Arellano W. Genetic diversity of tyrosine hydroxylase (TH) and dopamine β-hydroxylase (DBH) genes in cattle breeds. Genet Mol Biol. 2012; 35(2): 435-440. [ Links ]
18. Maudet C, Luikart G, Taberlet P. Genetic diversity and assignment tests among seven French cattle breeds based on microstellite DNA analysis. J. Anim. Sci. 2002; 80(4): 942- 950. [ Links ]
19. Miller S, Dykes D, Poletsky H. A simple Salting out procedure for extracting DNA from human nucleated cells. Nucl Acids, 1988. Res 16, 1215. [ Links ]
20. NCBI. National Center for Biotechnology Information, U.S.. National Library of Medicine. Gene ID: 280758.. Rockville Pike, Bethesda MD, 20894, USA: [acceso: 5 de julio de 2015]. URL: http://www.ncbi.nlm.nih.gov/gene/280758. [ Links ]
21. Norris D, Ngambi J, Mabelebele M, Alabi O, Benyi K. Genetic selection for docility. Anim. Plant Sci. 2014; 24(1): 13-18. [ Links ]
22. Peakall R, Smouse P. Genalex 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics, 2012; 28(19): 2537-2539. [ Links ]
23. Piovezan U, Dos Santos J, Rodrigues M. Breed and selection line differences in the temperament of beef cattle. Acta Sci., Anim. Sci. 2013; 35 (2): 207-212. [ Links ]
24. Pugh K, Stookey J, Buchanan F. Evaluation of corticotropin-releasing hormone and leptin SNPs relative to cattle behaviour. Can. J. Anim. Sci. 2011; 91(4): 567-572. [ Links ]
25. Romero M, Uribe l, Sánchez J. Biomarcadores de estrés como indicadores de bienestar animal en ganado de carne. Biosalud, 2011; 10 (1): 71-87. [ Links ]
26. Sant'Anna A, Paranhos da Costa M, Baldi F, Rueda P, Albuquerque L. Genetic associations between flight speed and growth traits in Nellore cattle. J. Anim. Sci. 2012; 90(10): 3427-3432. [ Links ]
27. Silva B, Gonzalo A, Cañón J. Genetic parameters of aggressiveness, ferocity and mobility in the fighting bull breed. Anim. Res. 2006; 55(1): 65-70. [ Links ].
28. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA 6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evolution. 2013; 30(12): 2725-2729. [ Links ]
29. UCTL. (2010). Union de criadores de toros de Lidia. Madrid - España : ISSN: 1888-2889. [ Links ]
30. Vaca R. Temperamento y bienestar, Efecto sobre la producción de bovinos de carne. Rev. Analecta Veterinaria, 2010; 30(2): 74-81. [ Links ]
31. Vann R, Riley D, Burdick N, Carroll J, Welsh T, Randel R. (2013). Evaluation of Methods of Temperament Scoring for Beef Cattle R.C.. Animal and Dairy Sciences Department Report. Division of Agriculture, Forestry, and Veterinary Medicine. Mississippi Stat. J. Anim. Sci. 91(E-Suppl. 2). [ Links ]
32. Voisinet D, Grandin T, Tatum J, O'Connor S, Struthers J. En engorde a corral, el ganado calmo gana más peso por día que el ganado arisco. Journal of Animal Science. 1997; 75 (4): 892-896. [ Links ]
33. Williams A, Boles J, Kohlbeck K, Meyers M, Perz K, Thomson J, et al. Effect of temperament and growth rate on carcass characteristics of Simmental cross steers. Ponencia. 60th International Congress of Meat Science and Technology, Maldonado, Uruguay, 17- 22 August 2014. [acceso: 11 de abril de 2015]. http://www.cabdirect.org/abstracts/20153118948.html;jsessionid=2E1F47A46397ED51E8858F65137E6305. [ Links ]

