











 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Los recientes avances en biología molecular y bioestadística12 han facilitado que se estudie la variación genómica y su posible asociación con parámetros productivos y reproductivos de interés en ganadería y reproducción humana.
Una de las posibilidades es el estudio de los polimorfismos, que se han definido como una variante en la secuencia de ADN que tiene una frecuencia mayor al 1% de la población 9. El estudio de los polimorfismos de los genes que regulan la función reproductiva permitirían clarificar los mecanismos de la función gonadal y la fertilidad en las especies de interés 3, en el humano se propuesto que si bien es cierto exista la posibilidad que el polimorfismo per se no afecte la proteína si se ha asociado a ciertos fenotipos, para el caso infertilidad 1.
Se propone que, más allá de la identificación del polimorfismo y sus asociaciones, el estudio combinado del genoma de los individuos junto con los fenotipos pudiera generar información sobre la naturaleza de los eventos que se estudian y desde la biología reproductiva comprender los fenómenos observados para proponer estrategias que mejoren los resultados cuando se aplican los conceptos desarrollados en las diferentes técnicas en los sistemas de producción.
En el caso eventos reproductivos, estudios previos en el laboratorio de Biotecnología Animal de la Universidad Nacional de Sede Medellín, han demostrado asociación entre los parámetros reproductivos y productivos de los genes receptor de la hormona folículo estimulante (FSHr), factor de crecimiento insulinoide (IGF1); inhibina A (InhA)4. En humanos se ha informado que existe asociación entre un polimorfismo de la hormona antimulleriana (AMH) y su receptor tipo II (AMHRII) 5. Un polimorfismo génico es una variante en la secuencia de ADN que tiene una frecuencia mayor al 1% de la población 9. El estudio de los polimorfismos de los genes que regulan la función reproductiva permitirían clarificar los mecanismos de la función gonadal y la fertilidad en las especies de interés 3, en el humano se propuesto que si bien es cierto exista la posibilidad que el polimorfismo per se no afecte la proteína si se ha asociado a ciertos fenotipos, para el caso infertilidad 1.
El potencial reproductivo de una hembra es el resultado del delicado balance entre la cantidad de folículos primordiales producidos en la vida fetal que son almacenados hasta la pubertad y su utilización después de la pubertad. Los folículos son reclutados durante cada onda y finalmente son ovulados, este balance es lo que le permite a cada hembra cumplir su función reproductiva 6 La mayoría de los folículos formados se atresian antes de la pubertad 7 y cada folículo inicia su crecimiento gracias a que adquiere receptores para la FSH y así poder continuar hacia la ovulación 8. Todo el conocimiento asociado a estos fenómenos ha sido obtenido de modelos animales, principalmente individuos genéticamente modificados, monos Rhesus y especies de granja 8,9,10.
Mantener ese balance entre la cantidad de folículos que se atresian y los que son utilizados para una ovulación es el resultado de la interacción de diversos factores controladores de la reserva ovárica y los efectores del desarrollo folicular como: el factor transformante b, la hormona antimulleriana (AMH) y las gonadotropinas (hormona folículo estimulante y su receptor, FSH,FSHr, hormona-LH y la inhibina). El control de la reserva ovárica depende principalmente de la interacción entre la FSH y su receptor y moléculas autocrinas pertenecientes a la familia del factor transformante beta 11. Para la ovulación, el control del paso de folículo primordial a folículo primario depende de la relación estrecha entre AMH, FSH y algunos factores de crecimiento, cuya función dado que son proteínas extracelulares depende de la expresión de receptores en las células de granulosa 10.
La AMH ejerce su función biológica mediante su unión al receptor tipo II de la hormona (AMHRII), que a su vez activa el receptor tipo I (AMHRI) desencadenando la via de señalización de las proteínas SMAD, en el humano, en el humano el AMH está localizado en el cromosoma 19 y el gen AMHRII está en el 12, posee 11 exones y se expresa exclusivamente en las células de granulosa y de la teca 12, en los vacunos están localizados en el cromosoma 7 y 5, respectivamente, sin que hayan sido informados polimorfismos en esta especie. En humanos se han informado dos polimorfismos en el gen de la AMH y dos en el AMHRII 13 que se han podido correlacionar con el estado reproductivo de mujeres adicionalmente también se ha asociado con los niveles de estradiol, al principio de la fase folicular durante el ciclo menstrual, mostrando el papel de la AMH en la regulación de sensibilidad a la FSH5.
La FSH es una glicoproteína que no puede pasar la membrana celular, lo que hace que toda su función se realice a través de su receptor (FSHr). FSH r está localizado en el cromosoma 11 de los vacunos, (FSHr), pertenece a la familia de las proteínas G, expresada por las células de la granulosa de la que se ha informado que mutaciones del mismo han sido asociadas con alteraciones en la reproducción. 14 Yang y colaboradores han informado la asociación de un polimorfismo en el promotor del gen del receptor con la respuesta a la superovulación en vacas Holstein 15.
La inhibina es una proteína dimérica compuesta por una subunidad A y dos subunidades B, ambas producidas en las células de la granulosa de los folículos, el gen está localizado en el cromosoma 2 de los bovinos. Esta proteína juega un importante rol en la supresión de la secreción de hormona folículo estimulante, y la concentración de ambas proteínas en sangre tiene una relación directamente inversa, por lo que afectan directamente el crecimiento folicular16. La subunidad alfa de la inhibina ha sido asociada principalmente con características reproductivas, encontrándose asociación entre polimorfismo del gen y la taza de superovulación en vacas Holstein, por lo que se ha postulado como un gen candidato para mejorar el desempeño reproductiva 17.
Dada la gran homología existente entre las secuencias de cada uno de los genes mencionados entre especies de bovinos, que los genes tengan la misma función, la poca información existente en la especie bufalina, y la posibilidad que una forma nueva de explicar los fenómenos observados sea la comparación entre especies, el objetivo del presente trabajo fue evaluar si en la especie bufalina y en vacunos indicus se encuentran los mismos polimorfismos asociados a parámetros reproductivos descritos en vacunos Holstein y humanos en genes importantes para el desarrollo folicular que han sido reportados por otros autores.
Materiales y métodos
Localización
Las muestras para este trabajo fueron obtenidas en dos explotaciones ganaderas del Magdalena Medio Colombiano, 6°18´48´´N 73°57´00´´O, en una zona clasificada como bosque húmedo tropical, a 120 MSNM, con una temperatura promedio de 24oC, humedad de 79% y una precipitación anual de 2766 mm3. Durante el año 2018, se tomaron muestras de sangre con anticoagulante EDTA (EDTA, Vacutainer, Beckton Dikinson). de 50 búfalos (Bubalus bubalis) mestizos y 50 vacas (Bos indicus) cebú comercial, sin alteraciones evidentes en su anatomía externa y en sus genitales internos, Cada especie con un peso y edad promedio similar 538 kg. vs 427 kg y 7,34 años vs 7,16 años para búfalos y vacas, respectivamente. Se registró la información sobre de paridad, días abiertos, intervalo parto primer servicio y edad al primer parto. Los animales fueron mantenidos en las mismas condiciones de alimentación y manejo, pastoreaban (Brachiaria humidicola) y consumían una mezcla mineral a voluntad, además de tener un manejo extensivo.
Al momento de la selección de los animales, la muestra tomada fue dividida en dos partes, una destinada para la determinación de los niveles de AMH, y la otra para la obtención de plasma que fue obtenido después de la centrifugación y congelado a -20oC hasta el momento de la determinación de AMH mediante el uso de un kit comercial AMH ELISA. (Cat No AL-114 Lot No 010616-B Ansh Labs, Webster, TX, USA.).
Extracción de ADN e Identificación de los polimorfismos
Para la obtención de ADN genómico se utilizó la técnica de Salting out modificada y descrita previamente 18.
Identificación de los polimorfismos
En la tabla 1, se muestra la información sobre los primers, la especie y la metodología usada para la identificación de los segmentos a evaluar, las condiciones de cada amplificación se encuentran en las referencias citadas para cada caso. Para la identificación de los polimorfismos de los genes FSHr e InhA se utilizó la técnica de reacción en cadena de la polimerasa más los fragmentos de restricción de longitud polimórfica (PCFR-RFLP), para los genes AMH yAMHr se utilizó la técnica de alta resolución de fusión (HRM, del inglés high melting resolution).

Dada la escasa información existente para los búfalos y los vacunos indicus se decidió utilizar como controles positivos el mismo tipo de muestras usadas en los artículos de referencia, en el caso de los genes FSHr e Inhibina, vacunos de la raza Holstein, a los que previamente se les había realizado la genotipificación en el laboratorio, y para los genes AMH y AMHrII muestras de humano.
Se realizó adicionalmente una búsqueda en los bases de datos de National Institute of Health en las bases de datos Gene, Genome and Protein, y se utilizó BLAST para la comparación de las secuencias de ADN, RNA y proteína de los genes estudiados. Fueron usados como genomas de referencia para vacas Bos taurus (assembly ARS-UCD1.2), para humano : Homo sapiens (assembly GRCh38.p13), vacas indicus: Bos indicus (assembly Bos_indicus_1.0) y para búfalos Bubalus bubalis (assembly UOA_WB_1).
La información sobre los parámetros reproductivos es presentada como la media ± la desviación estándar, fueron comparados por especie. Se estimaron las diferencias estadísticas fueron usando la prueba Mann Whitney utilizando el software GraphPad Prism 7.0 (GraphPad Software, San Diego, CA, USA), se consideraron diferencias significativas valores de p < 0,05.
Aval del comité de ética
El Comité Institucional para el Uso y Cuidado de los Animales en Investigación de la Sede Medellín, constituido mediante el Acuerdo 010 de 2016 del Consejo de Sede (CS), tomó en consideración el componente ético involucrado en la propuesta de investigación “Identificación de polimorfismos en genes asociados a la reserva ovárica y proteómica de oocitos en vacunos y bufalinos”. Al respecto, el Comité ha decidido otorgar el aval ético para desarrollar el proyecto, mediante el oficio CICUA-013 de octubre 2017.
Resultados
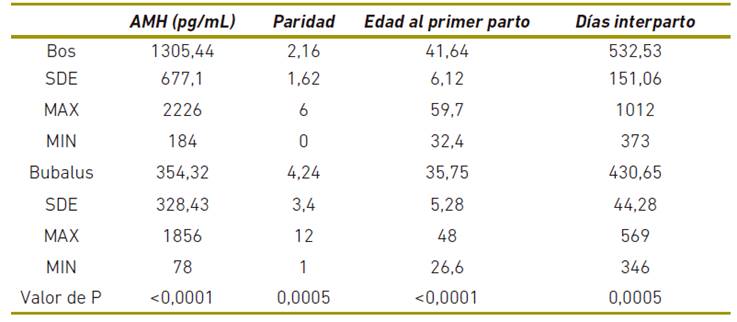
Los parámetros reproductivos de las dos especies evaluadas en este este estudio se muestran en la tabla 2. Los búfalos tienen menores niveles de AMH, edad al primer parto, intervalo ente partos que las vacas (p<0,001), mayor paridad (p=0,005). A pesar de seleccionar los animales al azar, se observan diferencias significativas en los días postparto al momento de la toma de muestra y la paridad (p=0,005).
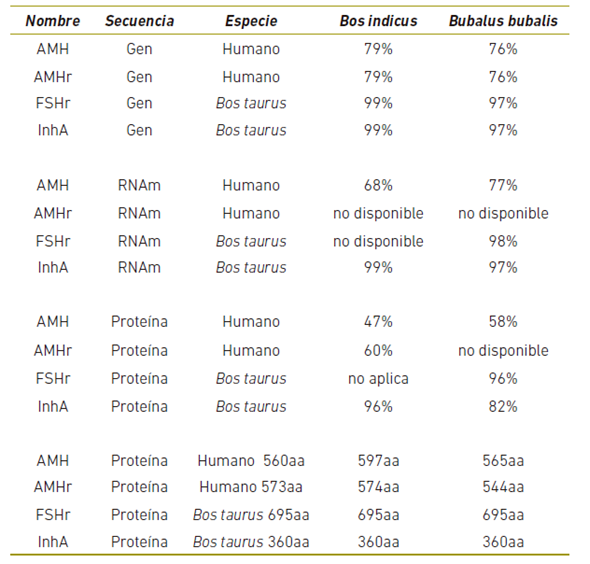
En la tabla 3 se muestran los resultados de la búsqueda en la literatura y las bases de datos, se pudo confirmar, la alta homología existente entre la secuencia de los genes evaluados, los RNAs y las proteínas.
En ninguna de las muestras de búfalos y de vacunos del estudio se pudo demostrar el polimorfismo descrito, debido a que no se obtuvieron amplificados con los primers descritos para los genes FSHr, InhA, AMH y AMHr., es importante mencionar que si se pudieron confirmar los polimorfismos en las muestras control.
Para los genes de AMH y AMHr, con la técnica de HRM, se pudo amplificar un fragmento de 143 pb y de 150 pb, respectivamente en todas las muestras humanas evaluadas, no se lograron amplificaciones en los vacunos ni en los búfalos. Cuando se evaluó la amplificación del gen AMHr utilizando primers vacunos, se lograron amplificaciones solamente en los vacunos no hubo amplificación en los búfalos, ni en los humanos, en los vacunos se encontró un fragmento entre 271 y 278.
Discusión
Los parámetros reproductivos de los animales utilizados en este experimento son comparables con lo informado en la literatura para búfalos 19 y para vacunos 20, y la búsqueda de explicaciones a estas diferencias son la base de la presente investigación. No confirmar los polimorfismos hallados en el ganado Holstein por otros autores, aun confirmando que tienen la misma función entre las especies, 21,22 lleva a replantear la forma de estudiar los fenómenos que se observan o a analizarlos de manera distinta. Existen algunos argumentos que favorecen las posibilidades de no encontrar correlaciones entre la estructura de los genes y la función, Cory et al. evaluaron 10 polimorfismos en los exones del Gen FSHr en bovinos de la raza Holstein y encontraron que siete producían mutaciones silenciosas14, otros autores han informado que ciertos SNP en los genes de interés no se asocian con parámetros reproductivos 23. Estas evidencias han obligado a los investigadores a incluir algunos aspectos técnicos de la investigación como parte de la discusión de los resultados.
Si bien es cierto que se ha informado en mujeres la existencia de un SNP en el gen FSHr, asociado a la cantidad de dosis requerida de FSH para superovulación 24, en bovinos y sus diferentes razas no se ha confirmado y existe poca evidencia en la literatura sobre el tema. Cory, et al., informaron en diferentes razas de bovinos polimorfismos en el receptor de la FSH 14. Hirayama estudio en Japón algunos polimorfismos del gen FSHr, en vacunos de la raza Japonesa Kedaka asociada con la cantidad de embriones transferibles 26, sin embargo hay que notar que si bien es cierto son polimorfismos no son los mismos. Milazotto et al., en el 2008 informó la existencia de polimorfismos en el ganado Nellore25 asociados con la pubertad. En referencia con los búfalos se debe mencionar que el receptor fue clonado desde el 2008 27 sin que hasta la fecha se hayan informado polimorfismos en el gen.
A diferencia de lo que ha pasado en los humanos 3, en los bovinos no se han informado polimorfismos en el gen de la AMH, solamente Nawaz et al., utilizando un chip de marcadores de alta densidad informó de la existencia de un grupo de 11 marcadores SNPs localizados en el cromosoma 11 bovino 2, para el caso del AMHr no se encontraron informes sobre polimorfismos en las especies de bovinos estudiadas.
Inhibina A, Sang et al., han informado asociación de polimorfismos con parámetros reproductivos en animales de la raza Holstein 28, no se han informado estudios similares en vacunos cebuinos o bufalinos.
La comparación de las secuencias de los genes de las dos especies estudiadas mostró una gran homología, aun comparando con el patrón Bos taurus y como era de esperarse la mayor divergencia se dio con los humanos. Cuando se comparan las secuencias de AMH y AMHr2 de los bovinos entre si la homología es bastante grande. Debe hacerse notar que para algunos genes como el FSHr y el AMHr2, no se ha reportado la secuencia de RNA , ni la secuencia de la proteína para los bovinos estudiados. Desde el punto de vista de la función, la estructura y la secuencia de las proteínas es bastante similar entre todas las especies y genes evaluados.
Con el conocimiento moderno, pudiera pensarse que para la obtención de amplificados, debiera haberse analizado primero una secuencia de primers o diseñado algunos específicos para amplificar las mismas regiones estudiadas, pero el equipo de investigación decidió probar las secuencias descritas en los controles escogidos, basado fundamentalmente en la homología de secuencias de RNA y la función de los genes, puede que metodológicamente sea incorrecto pero fisiológicamente tiene muchísimo sentido.
Después de evaluar los resultados , se analizó si los primers reconocían las mismas secuencias que habían sido informadas en la literatura, con la gran sorpresa que a pesar de todas las gran homología, ninguno de los primers usados reconocía secuencias de los genes propuestos en las especies propuestas, evidenciando las diferencias en secuencia, pero lo más importante es que la información no debe ser extrapoladas tan fácilmente entre especies y en este caso tan cercanas filogenéticamente, no olvidar que la función de la proteína no se altera, aun con las diferencias en secuencia planteadas.
La interrelación de las proteínas en este estudio es bastante grande, AMH y la expresión de su receptor (AMHr2) controlan la población de folículos primordiales que tiene la hembra bovina, estos folículos deben desarrollar receptores para la FSH y convertirse en folículos preantrales para que se dé el proceso ovulatorio. Para que se de el tránsito de folículo antral a ovulatorio aparecen las otras hormonas estudiadas en este trabajo, InhA, IGF1, adicionalmente InhA controla la secreción de FSH a nivel hipofisiario 29 30 . Finalmente, AMH e InhA pertenecen a la familia del factor transformante-β (TGFβ), cuya actividad es crítica para el control del crecimiento, diferenciación y destino celular.
La forma cómo actúan los genes estudiados y sus receptores, corresponden a moléculas extracelulares que están involucradas en el envío de señales dentro de la célula, bien sea a través de proteínas G o mediante la activación del SMAD, esta evidencia fortalece la idea de ver la función de las gonadotropinas con una relación de espacio-temporalidad así pues la misma molécula pude tener diferente función dependiendo el momento y lugar, en consecuencia los análisis tienen obligatoriamente dos enfoques, el que tiene que ver con el momento reproductivo del individuo, pudiera decirse momento del ciclo estral o de la onda folicular y el segundo con los aspectos básicos de biología celular que cada tejido y órgano tiene que cumplir y que han evolucionado a la par con las especies.
Este estudio demuestra como la homología en la función no quiere significa que haya necesariamente una homología en estructura de los genes y proteínas, especialmente cuando se quieran extrapolar los resultados en otras especies donde la información es escasa o inexistente como en este caso los búfalos.
La identificación de SNP o en general de polimorfismos, no necesariamente tienen que tener relación de causalidad, estos polimorfismos de los que no se duda su existencia pueden servir de marcadores de la función reproductiva. Teniendo claro que para lograr este objetivo hay que tener en cuenta el tamaño de la muestra, los aspectos genéticos del mismo y la naturaleza del fenotipo observado lo que nos vuelve a la gran discusión entre la estructura del gen y la función, adicionalmente plantea interrogantes sobre el control génico que pueden estar relacionados con la función, pero no necesariamente que sean los mismos entre las especies.
Conclusiones
De los resultados obtenidos en este trabajo no se pudieron confirmar en los vacunos indicus y los búfalos los polimorfismos descritos en otras especies, aun demostrando que existen diferencias en parámetros reproductivos No se puede confirmar que el estudio comparativo entre especies permita obtener información la biología de la reproducción al menos para el caso el desarrollo folicular y los parámetros reproductivos, que cada genoma requiere su propio estudio y análisis antes para poder proponer las posibles correlaciones con fenotipos escogidos , Se requiere entonces hacer un análisis de las regiones propuestas para comparación en búfalos y vacunos indicus, para poder proponer estrategias de análisis o diseñar experimentos que permitan explicar los fenómenos observados.

