











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Las mejores características genéticas permiten incrementar los rendimientos productivos de los bovinos para alcanzar superioridad en cuanto a su mayor desarrollo, resistencia a enfermedades, incremento de fertilidad y mejor calidad de leche y carne 31,25,69,78. Existe cierta complejidad en la identificación de animales que tienen la capacidad de producir descendencia superior con rasgos deseables o alto mérito genético, puesto que la mayoría de estas características son cuantitativas y controladas por grupos de genes que interactúan entre sí y con el medio ambiente 3.
El estudio del genoma bovino permitió el descubrimiento de polimorfismos de nucleótidos simples (del inglés SNPs) y su relación con el potencial productivo de los animales. Se ha logrado identificar a los locus de caracteres cuantitativos (quantitative trait locus) que influyen en los caracteres productivos de los bovinos 8,51. En estos estudios se identifican y analizan SNPs de genes que tienen influencia en las características productivas; permitiendo obtener parámetros genéticos más precisos 8,51. Además, se han identificado SNPs puntuales en los genes de la calpaína (CAPN) y calpastatina (CAST), que facilitan la identificación del valor genético del bovino. Estos SNPs permiten mejorar ciertas características de interés como la terneza, marmoleo, jugosidad, entre otras propiedades de calidad de la carne. Por ejemplo, el gen de la CAPN ubicado en el cromosoma 29 del bovino 69, el cual consta de 22 exones y 21 intrones 57,36 y el gen de la CAST ubicado en el cromosoma 7 37 comprendiendo en 35 exones 65 presentan SNPs que se asocian con ciertas características de calidad de la carne (Figura 1).
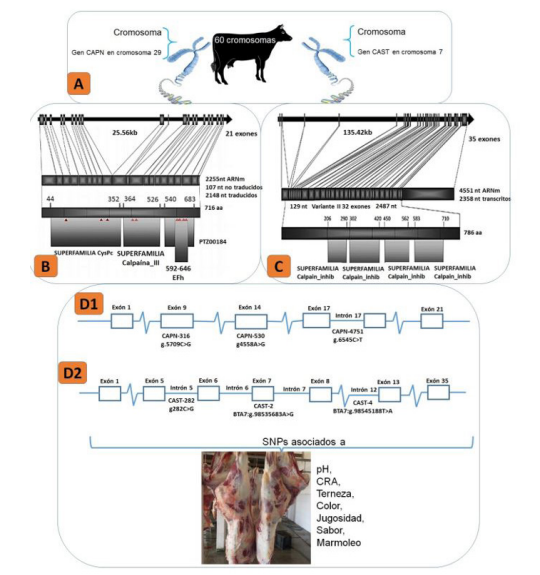
Figura 1 La variación en un solo par de bases por transición o transversión conlleva a formar un polimorfismo de nucleótido simple. A: Polimorfismos de nucleótidos simples de genes CAPN1 y CAST que se asocian a propiedades fisicoquímicas y organolépticas de la carne bovina se encuentran en el cromosoma 7 y 29. B: Estructura del gen CAPN1, se presentan los dominios predichos en la proteína lineal y los ΔΔΔ de la izquierda es el sitio catalítico, ΔΔ central es la formación de un loop ácido involucrado en la activación de la molécula durante la unión al calcio y ΔΔΔΔ de la derecha representa el sitio de unión al calcio de aminoácidos que constituyen los sitios activos de la molécula. C: Organización del gen CAST y los dominios predichos en la proteína lineal como resultado de la variante II de CAST son presentados 41. D1: SNPs del gen CAPN1 como CAPN-530 está ubicado en el exón 14, presenta una transición de Adenina/Guanina 57, CAPN-4751 está ubicado en el intrón 17, presenta una transición de Citosina/Timina 80, CAPN-316 está ubicado en el exón 9, presenta una transversión de Citosina/Guanina 57. D2: SNPs del gen CAST como CAST_2 está ubicado en el exón 7, presenta una transición de Adenina/Guanina 7. CAST-282 está ubicado en el intrón 5, presenta una transversión de Citosina/Guanina 70. CAST_4 está ubicado en el intrón 12, presenta una transversión de Timina/Adenina 7
El sistema proteolítico de las calpaínas es el responsable del ablandamiento de la carne, así la conversión de músculo a carne está regulada por la degradación proteolítica post mortem regulada por la enzima calpaína y la enzima inhibidora calpastatina. En el tejido muscular existen las enzimas proteolíticas como la µ-calpaína, m-calpaína y p94 77. La calpastatina es una proteína que inhibe la actividad de la µ-calpaína y m-calpaína 34. Las isoformas de calpaína influyen en la proteasa micromolar neutra que es activada por la µ-calpaína, cuya función es codificar una cisteína proteasa que degrada a proteínas miofibrilares post mortem (14,22.
La calidad de la carne depende de su proceso productivo, procesamiento y manejo peri mortem, donde existen efectos no genéticos que pueden afectar la calidad de la carne como el tipo de alimentación, sistemas de crianza, la raza, el sexo, castración en machos, composición de tejido conectivo y estrés pre faenado 18,53,64, y en efectos genéticos donde ocurren procesos moleculares que se dan en las etapas ante mortem y post mortem 13,12.
En este estudio analizamos los avances en el conocimiento sobre las asociaciones de polimorfismos de CAPN1 y CAST con las propiedades fisicoquímicas de la carne bovina, información que contribuirá a establecer líneas de investigación en especies de interés para mejorar la calidad de la carne.
CAPN1, CAST y la terneza de la carne
La terneza de la carne es una variable organoléptica, que impacta los aspectos más importantes en el ganado bovino, ya que está asociado con la aceptación y satisfacción del consumidor. Sin embargo, la terneza de la carne depende de la edad, la genética, el sexo y el manejo integral del animal 24,26. Asimismo, depende del tipo de músculo, donde intervienen las fibras musculares y el colágeno 67,18. En la etapa post mortem, el tejido muscular se convierte en carne y su terneza depende la actividad proteolítica. Algunos marcadores en genes de CAPN1 y CAST se vienen evaluando debido a que existe evidencia validada de que están altamente asociados con la terneza de la carne 6,61,73.
Dentro de los genotipos de CAST-2959 (AF159246:g.2959G>A) del gen CAST, el genotipo TT está asociado con una mayor terneza debido a que presenta menor actividad de la calpastatina y que interactúa menos con la µ-calpaína 10. Este efecto se ve reflejado en el hallazgo de Smith et al. (2009) 73, en el cual el genotipo TT se asoció a una fuerza de corte menor luego de 14 días de maduración de la carne (Tabla 1). En músculo Longissimus de poblaciones Bos taurus y cruces de Bos taurus con Bos indicus, los bovinos con herencia de genotipo CC y CT producen carne con mayor terneza respecto a bovinos con herencia de genotipos TT 10. En poblaciones de bovinos de la raza Angus, Limousin, Charolais y Simmental, el genotipo CC produce carme con mayor terneza respecto a GG y GC en el SNP CAST-282 (AY008267:g. 282C>G) 70 y en bovinos Nelore con genotipo CC también tienen carne con mayor terneza respecto a bovinos con genotipo CG y GG (SNP CAST-282) y el CC con mayor terneza en comparación con CT y TT del SNP CAPN-4751 (AF248054:g.6545C>T) 9. En el SNP CAPN-4751, el genotipo CT presentan carne con mayor terneza respecto a TT en bovinos Nelore y cruces de Angus con Nelore, Canchim, Brangus y Braunvieh 21) (Figura 2a). Dentro de los genotipos del SNP CAPN-316 (AF252504:g.5709C>G), el GG produce carne con mejor terneza respecto a CG y CC en bovinos Aberdeen Angus 29 (Figura 2b).
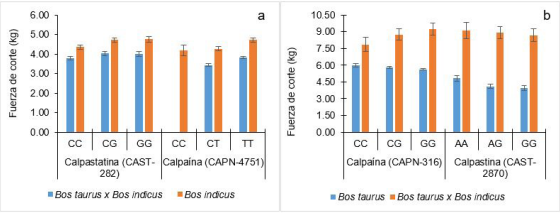
Figura 2 Media de mínimos cuadrados de terneza de carne, evaluado por fuerza de corte Warner Bratzler. Efecto del SNP CAST-282 en la terneza de la carne de bovinos ≥ 5/8 de sangre para cada raza (Angus, Limousin, Charolais, Simmental) y bovinos Nelore a 21 días post mortem, y efecto del SNP CAPN-4751 en la terneza de la carne de bovinos Nelore y cruces de Angus con Nelore, Canchim, Brangus y Braunvieh (a). Efecto del SNP CAPN-316 sobre la terneza de la carne de bovinos Aberdeen Angus (entre 21 a 30 días post mortem), cruces de Angus con Hereford y Limousin con Hereford-Angus y efecto del SNP CAST-2870 en la terneza de la carne de bovinos cruzados Angus con Hereford, Limousin con Hereford-Angus, Limousin con Luxi, Charolais con Fuzhou, Simmental con Mongolian y razas puras de Luxi, Jinnan, Angus y Hereford (b). Líneas de color negro en la terminación de cada barra indicar error estándar de la media. Adaptado de Carvalho et al. (2017) 9, Schenkel et al. (2006) 70, Gill et al. (2009) 29, Li et al. (2010) (46
En bovinos, a medida que la carne es madurada por más tiempo, la fuerza de corte es menor; por ejemplo, en bovinos Nelore la fuerza de corte baja de 5.02 kg/cm2 al séptimo día hasta 3,57 kg/cm2 al día 21. Los SNPs CAPN-4751, CAST-282 y CAST263 de los genes CAPN1 y CAST se asocian con la fuerza de corte en el músculo Longissimus dorsi en carne madurada en tres tiempos 61. Valores similares también se da en carne de bovinos Brahman al analizarse SNPs CAPN-316 y CAPN-4751 73. El atributo que se le confiere al CAPN-4751 es por la transición de citosina por timina y CAPN-316 por la sustitución de la base citosina por guanina, logrando codificar al aminoácido de alamina y sustituirla por glicina 57 (Tabla 1).
CAPN1, CAST y color de la carne
La característica organoléptica que tiene gran influencia sobre las decisiones en la compra de la carne es su color 49. El estado fisicoquímico de la mioglobina y la estructura del músculo están relacionado al color final de la carne. La estructura muscular está influenciada por el pH final, por ejemplo, pH final alto da como resultado una carne con fibras musculares más juntas y aumenta su capacidad de retener agua de la proteína muscular 68,1. La µ-calpaína (P07384) se correlaciona con todas las coordenadas de color del sistema CIEL*a*b* (L* representa luminosidad desde blanco a negro, a* es tonalidad de rojo a verde y b* es tonalidad de amarillo a azul) en carne de bovinos de raza French Blond d’Aquitaine 28. La variabilidad positiva (47%) por µ-calpaína es explicada por la abundancia de color amarillo en el músculo y la actividad de la µ-calpaína con la proteína de choque térmico de 70 KDa o Hsp70 que influyen en las coordenadas del color de la carne de manera interactiva 28,27. Este mecanismo está dado porque el daño de especies reactivas de oxígeno post mortem del retículo endoplasmático de las células, permiten liberar Ca2+ en el citosol, activando de esta manera la µ-calpaína 17,30. Por consiguiente, las proteínas son el sustrato principal de la µ-calpaína y sus características influyen en el color de la carne. Al aumentar los niveles de Hsp70 carbonilados a las proteínas estructurales se inhibe la actividad de la µ-calpaína, logrando aplazar los cambios en la estructura del pigmento y las proteínas miofibrilares. Este mecanismo influye en la reflectancia y otros aspectos del color de la carne 28.
El SNP CAPN-4751 del gen CAPN1 presenta efectos aditivos y dominantes en la coordenada de +a* (color rojo) y +b* (color amarillo) en el músculo Longissimus de bovinos Nelore a 7, 14 y 21 días de maduración 62 (Figura 3a). Esto indica que el alelo T en bovinos de raza Nelore favorece el aumento del color rojo y amarillo en la carne, pero este mismo alelo (T) no ayuda a mejorar la terneza 61. El aumento del color rojo en la carne se explica por la pérdida de actividad respiratoria de las mitocondrias en el proceso de maduración, lo que conlleva a mayor disponibilidad de oxígeno en la superficie del músculo y este se pueda usar en la formación de oximioglobina de color rojo 55. El ángulo de tonalidad del color es mayor en carne de bovinos heterocigotos (CG) respecto a homocigotos (GG), pero que en heterocigotos no difiriere de homocigotos (CC: genotipo favorable) en el día 6 de maduración de la carne. Sin embargo, en el día cero no se asocia los SNPs CAPN-947 (AF252504:c.947C>G) del gen CAPN1 y SNP CAST-155 (NM_174003:c.155C>T) del gen CAST con las coordenadas de color 47. Este fenómeno se explica porque las medidas físicas del color aumentan conforme también se incrementan los días de maduración de la carne, debido a la desintegración del disco Z del músculo 15,56.
El SNP CAST-282 del gen CAST está asociado con los parámetros de color en Longissimus thoracis et lumborum y semimembranosus de bovinos cruzados irlandeses 66 (Figura 3b). Los procesos biológicos que dan sustento a la conexión que existe entre la calpastatina y los rasgos de color podrían involucrar un vínculo entre la concentración de calcio y la contracción muscular 20. Por otro lado, la calpastatina influye en la glucólisis y la disminución del pH, y por consiguiente podría influir en el color de la carne (66.
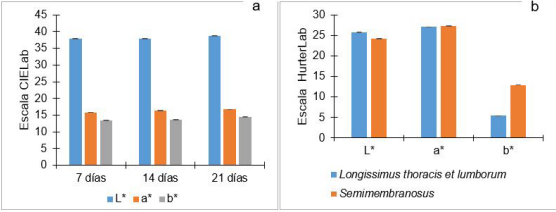
Figura 3 Media de color de carne (L: luminosidad, a* tonalidad de rojo a verde, b* tonalidad de amarillo a azul) en tres tiempos de maduración (a) y asociaciones de genotipos (CAST) con el color de la carne en dos tipos de músculos (b). Líneas de color negro en la terminación de cada barra indica error estándar de la media. Adaptado de Pinto et al. (2011) 62 y Reardon et al. (2010) 66
CAPN1, CAST y jugosidad de la carne
La jugosidad de la carne se evidencia durante la masticación (seca o no seca). Esta característica está determinada por una sensación de liberación de agua y luego es influenciado por la acción de los lípidos en la liberación de la saliva. Las virtudes sensoriales de la carne se ven afectadas y reduce su aceptabilidad cuando en una carne tiene jugosidad limitada. Existe evidencia en la que se asocia el SNP CAST2959 del gen CAST con la jugosidad de carne de animales cruzados de ascendencia Bos taurus. Sin embargo, el patrón no está muy claro debido a que bovinos con genotipo CT exhiben una carne con menor jugosidad respecto a bovinos homocigotos (CC o TT) 10. La carne del músculo Longissimus de bovinos de descendencia Bos taurus, presentó una puntuación de 5,30, cruce de Bos taurus con Bos indicus presentó 5,53 y Bos indicus presentó 5,25, de una escala de 1 al 8, donde 1 fue extremadamente duro y 8 fue extremadamente jugoso 10 (Figura 4). La selección asistida podría mejorar ciertos atributos de calidad de carne, pero también podría tener consecuencias no deseadas en otros rasgos de calidad de la carne.
CAPN1, CAST y sabor de la carne
Los principales precursores del sabor se dividen en componentes solubles en agua y lípidos. El sabor se deriva de la fracción muscular soluble en agua, pero las diferencias puntuales de la especie en el aroma de carne cocida se deben principalmente a concentración y composición de sustancias aromatizantes derivadas de los lípidos 60,74. Una autooxidación de los lípidos conlleva a producir un sabor rancio, pero una adecuada oxidación de los lípidos en la cocción de la carne da como resultado aromas deseables y por ende sabor deseable. Los compuestos volátiles generados a partir de grasa bovina calentada y la reacción de Maillard proporcionan aromas que producen componentes volátiles que dan sustento al sabor de carne cocida 60,74.
La carne del músculo Longissimus de bovinos de descendencia Bos taurus, presentó una puntuación de 4,89, cruce de Bos taurus con Bos indicus presentó 4,62 y Bos indicus presentó 5,88, de una escala de 1 al 8, donde 1 fue extremadamente soso y 8 fue extremadamente intenso 10 (Figura 4). Existen reportes de asociación del SNP CAST-2959 del gen CAST con el sabor de la carne de bovinos cruzados de Bos taurus con Bos indicus 10. Lo mismo ocurre en SNPs CAPN-530 (AF248054:g.4558A>G) del gen CAPN1, donde bovinos con genotipo CC y CT producen carne con un sabor más intenso respecto a bovinos de genotipo TT 10. Bovinos con herencia de genotipo CC producen carnes más tiernas y sabrosas (C alelo favorable para el marcador CAPN1, SNP CAPN-530) respecto a animales con genotipo TT; y los bovinos con genotipo TT producen carnes más duras o menos tiernas, pero los bovinos con genotipos TT en el SNP CAST-2959 del gen CAST presentan carne más tierna respecto a los bovinos de genotipo CC. Un sabor intenso se produce en el músculo Longissimus de bovinos Bos taurus, cruces de Bos taurus con Bos indicus y Bos indicus con genotipo CC en CAST-2959, y genotipo TT en CAPN-530 (10. Para la implementación de un plan de mejoramiento genético en un hato ganadero, se deben considerar las exigencias del mercado y evaluar el potencial de interacción de todos los genotipos posibles, para lograr una adecuada selección del plantel reproductor. Esto permitirá una producción de carne con características deseables y acorde a lo que busca el consumidor.
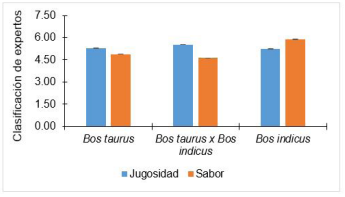
Figura 4 Media de mínimos cuadrados de jugosidad, evaluado por panelistas y clasificado en 1 = Extremadamente seco; 4 = ligeramente seco; 5 = ligeramente jugoso; 8 = extremadamente jugoso (barra azulada) e intensidad de sabor, 1 = extremadamente soso; 4 = ligeramente soso; 5 = ligeramente intenso; 8 = extremadamente intenso (barra anaranjada) de carne a 14 días post mortem. Líneas de color negro en la terminación de cada barra indicar error estándar de la media. Adaptado de Casas et al. (2006) 10.
CAPN1, CAST y marmoleo (marbling) de la carne
Uno de los factores más importantes que influye en la calidad cárnica es el marmoleo. El marmoleo se refiere a la percepción de calidad de la carne que están en función a la proporcionalidad de la cantidad y distribución de la grasa intramuscular 83. Se describe como la proporción de manchas de grasa intramuscular en los haces musculares 32. El gen CAPN1 está localizado en el cromosoma 29 72 y está formado por 21 exones y 20 intrones 23. Los exones 5 y 6 del gen CAPN1 en bovinos de raza Bali tienen ocho SNPs, pero solo CAPN-3669 y CAPN-3899 (NW_001494538) se asocian con puntuación del marmoleo 63 (Figura 5). Del mismo modo, se han reportado asociaciones del gen CAPN1 con la puntuación de marmoleo en bovinos de raza Angus, Jinnan, Simmental, Qinchuan y cruce de Simmental con Menggu. La sustitución de A/G en la posición 3553 del CAPN1 hace que los animales con genotipo AA tengan una mayor puntuación de marmoleo respecto a genotipos AG y GG 33 (Figura 5). Sin embargo, los índices de marmoleo pueden variar entre razas y de acuerdo a otros factores como las hormonas tiroideas que juegan un rol importante en la regulación del metabolismo y pueden afectar la homeostasis de los depósitos de grasa 71 y afectar el marmoleo. El estudio de ciertos SNPs pueden brindar la certeza de mejorar el marmoleo de la carne. A mayor y mejor distribución del marmoleo en el músculo, mayor es la aceptabilidad y palatabilidad de la carne. Para lograr este propósito es necesario comprender la arquitectura molecular del marmoleo.
La infiltración de grasa es un factor determinante en la calidad de la carne. El consumidor prefiere carne con mayor marmoleo debido a que está asociado a una buena palatabilidad 59. El tamaño de partículas y distribución de marmoleo ha logrado llamar la atención del productor porque se mejora las ventas y los consumidores buscan carne con marmoleo intermedio 44. El tamaño y la distribución de las partículas de marmoleo están relacionados positivamente con el precio de carne de bovinos 5. El marmoleo está asociado también con otros factores sensoriales como la terneza, jugosidad y palatabilidad de la carne 79. Existe diferencias significativas de marmoleo entre individuos de una misma raza, entre razas, el tipo de músculo y haces de fibras musculares 39,2. El uso de marcadores moleculares que se asocien a estas características importantes para selección de animales con ciertos atributos, constituyen una vía alterna a la selección genética tradicional. Ejemplo, los SNPs que se asocian con el marmoleo de la carne pueden servir para investigaciones con selección asistida por marcadores 33 (Tabla 1). Sin embargo, estudios con pocos animales podrías dar como resultado asociaciones falsas positivas 16. En consecuencia, se necesitan más estudios sobre las asociaciones de ciertos SNPs con parámetros de calidad de la carne 47.
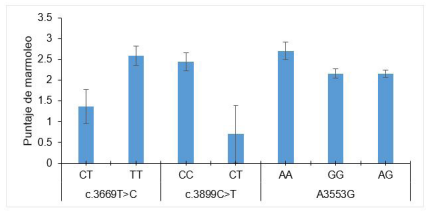
Figura 5 Asociación de SNP del gen CAPN1 con el marmoleo de la carne (puntuación de marmoleado realizada de acuerdo con el estándar de referencia de marmoleado Australian Meat). Líneas de color negro en la terminación de cada barra indicar error estándar de la media. Adaptado de Pratiwi et al. (2016) 63 y Hou et al. (2011) 33.
CAPN1, CAST y el pH de la carne
El estrés en bovinos impacta negativamente en las características del producto final, debido a que existe mayor agotamiento del glucógeno previo al sacrificio y menor producción de ácido láctico, traduciéndose en la disminución insuficiente del pH. En bovinos Bos taurus (Charolais, Limousin y Retinta), el pH promedio a 24 h post mortem en músculo Longissimus dorsi es inferior a 5,8 4. La carne de bovinos con registro de pH límite de 5,9 a 6,1 suelen ser muy duras 50.
En el músculo Longissimus thoracis et lumborum de terneros cruzados de Angus con Hereford se confirma la importancia de las enzimas (calpaína y calpastatina) sobre la terneza de la carne frente a la disminución del pH y la temperatura en la etapa temprana post mortem 35. La carne no siempre muestra niveles de µ-calpaína y calpastatina a 4 h post mortem, mostrando una mayor reducción de µ-calpaína respecto a calpastatina, es decir la calpastatina sería menos susceptible a cambios en pH y temperatura en periodos tempranos post mortem. Una adecuada maduración de la carne depende de los días de maduración y una disminución óptima de pH. Por ejemplo, se logra obtener una carne con mayor terneza a una disminución media de 5,9 a 6,2 de pH en 1,5 horas post mortem o de 29 a 30 °C a un pH de 6, después de 14 días de maduración de la carne 35. En bovinos cruzados irlandeses, el SNP CAST-282 del gen CAST se asocia significativamente con el pH en el músculo Longissimus thoracis et lumborum66. Los músculos de bovinos con herencia de genotipo GG presentan pH más alto respecto a bovinos de genotipo CC o CG (5,68 respecto a 5,57 y 5,59). Estos valores están dentro del rango aceptable, pero bovinos de genotipo GG podrían ser más propensos a producir carnes de categoría oscura, firme y seca respecto a bovinos de otros genotipos 81,66.
Los SNPs c.1735+794CTC, c.1735+920A>G y c.1735+951T>C (AF252504S1 y AF252504S2) ubicados en el intrón 16 del gen CAPN1 se asocian con el pH de la carne al día 0, 1 y 3 post mortem y el SNPs c.1795A>G ubicado entre el intrón 16 y el exón 17 del mismo gen, se asocian con el pH al 7 día post mortem 82. La importancia de validar SNPs depende de la naturaleza específica de la población bovina a examinar 82. Por lo tanto, un determinado polimorfismo no siempre puede asociarse a todos los parámetros de calidad de la carne. El fracaso de la validación está dado por la falta de asociaciones verdaderas entre el rasgo de calidad de la carne y el SNP, aunque también podría deberse a las diferencias en las frecuencias de los SNPs, las interacciones del genotipo por el entorno o epistasis, el tamaño de la muestra y la manera de medir el rasgo de calidad de carne 82.
CAPN1, CAST y capacidad de retención de agua de la carne
La capacidad de retención de agua (CRA) tiene valor productivo y económico, debido a la pérdida de peso en el producto final, que puede ocurrir tanto en el almacenamiento como en la cocción. La CRA está determinada por la degradación de ciertas proteínas citoesqueléticas del miocito 75. Los genes que influyen en la degradación de la estructura del miocito dependen del sistema proteolítico de µ-calpaína y calpastatina, que permite una interacción más estable de la matriz proteica con el agua, llegando a disminuir pérdidas de líquido de la carne 42. El SNP CAST-282 del gen CAST se asocia con la CRA en el músculo Longissimus thoracis et lumborum de la carne de bovinos irlandeses cruzados, al considerarse al pH a las 48 h post mortem 66. La conexión de la calpastatina y la CRA podría construir un vínculo a través de la concentración de iones de Ca++ y el ritmo de la contracción muscular 20. Asimismo, si la calpastatina está involucrada en la velocidad y alcance de la glucólisis, este fenómeno podría influir en la CRA 66. En cerdos se ha reportado correlaciones de la actividad de la calpaína con la retención de agua 52. Estas correlaciones probablemente se deban a que la CRA está influenciada positivamente, debido al agua retenida en la célula muscular (degradación rápida de filamentos intermedios) en el periodo inicial post mortem 40. Sin embargo, el SNP CAPN-947 del gen CAPN1 no se asocia con la CRA de carne de bovino Angus, Charolais, Hereford, Limousin y Simmental 47 (Tabla 1). Asimismo, no existe una relación significativa del SNP CANP-4751 y CAPN-316 del gen CAPN y el SNP CAST-282 del gen CAST con valores de CRA en bovinos Turco gris 38.
La CRA de la carne puede variar por la raza del bovino (Figura 6). El factor raza es significativo al evaluar la CRA en el músculo Longissimus dorsi de carne cruda. Las razas que presentan mayores valores son Braunvieh y Limousin 43. La CRA está relacionada con el nivel de la jugosidad de la carne. La carne que retiene mayor cantidad de agua en las fibras musculares ya sea de forma libre o inmovilizada conlleva a una carne más jugosa.
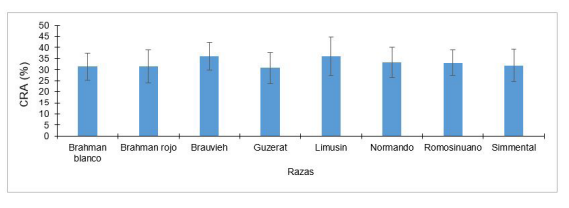
Figura 6 Valores promedio de capacidad de retención de agua en carne (Longissimus dorsi) en diferentes grupos genéticos, líneas de color negro en la terminación de cada barra indicar el error estándar de la media. Adaptado de Leal-Gutiérrez et al. (2014) 43
Tabla 1 Polimorfismos de CAPN y CAST asociados a parámetros de calidad de la carne bovina.
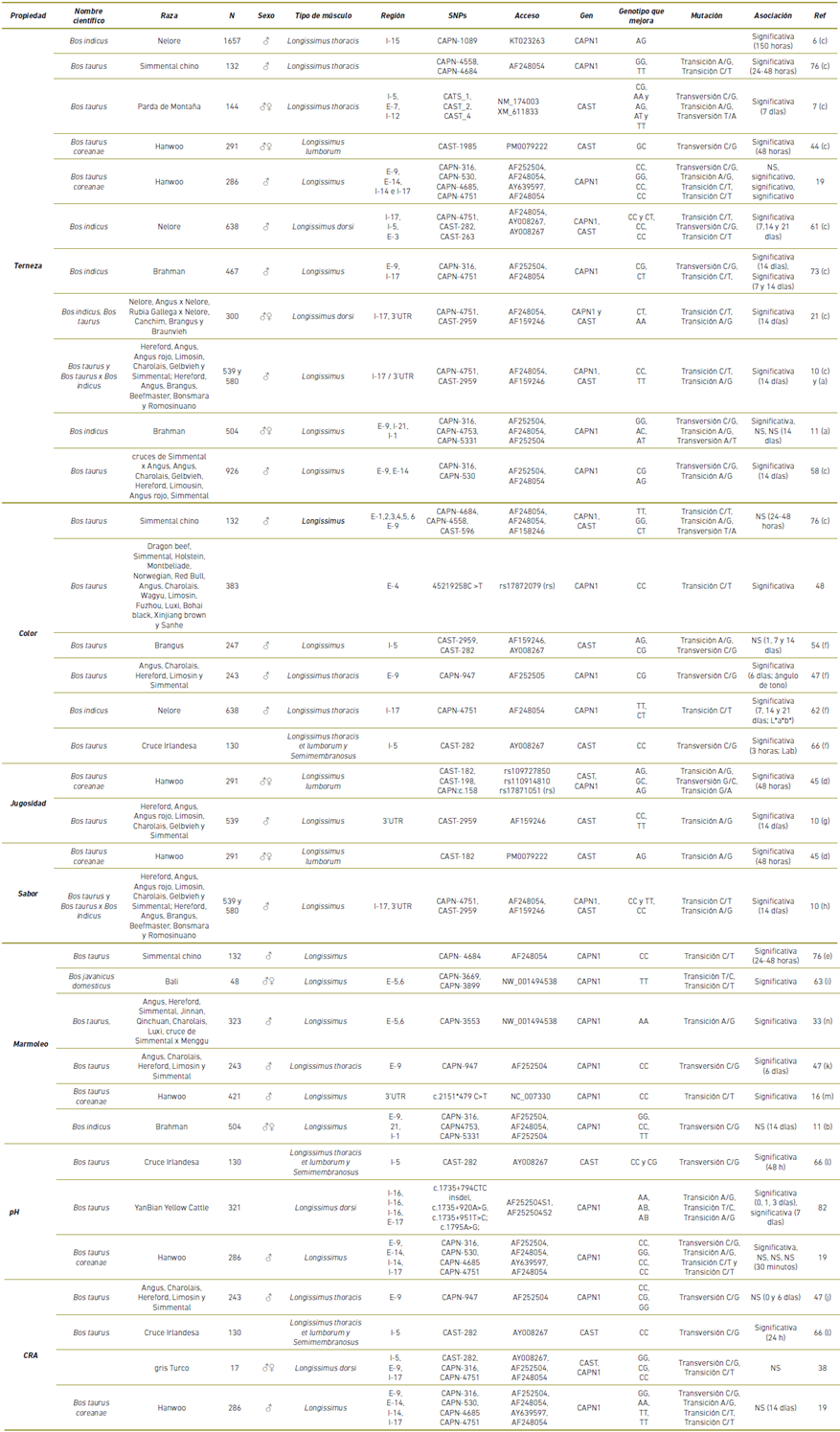
N: número de individuos usados para el estudio. I-: Intrón; E-: Exón, (a) Terneza de filete de 14 días medido con panel sensorial, usando escala de puntuación: (1 = extremadamente resistente; 4 = levemente resistente; 5 = levemente tierno; 8 = extremadamente tierno). (b) Medición del marmoleo: Devoid = 100 a 199; Trazas = 200 a 299; Ligero = 300 a 399; Pequeño = 400 a 499; Modesta = 500 a 599; y moderado = 600 a 699. (c) Terneza evaluada por la fuerza de corte Warner Bratzler. (d) Determinación de jugosidad y sabor con panelistas, usando escala: (1 = muy seco, no me gusta a 10 = muy jugoso, me gusta extremadamente. (e) Uso de estándares de fotografía para determinación de marmoleo. (f) Determinación del color con el sistema CIEL* a* b* y HurterL* a* b*. (g) Determinación de jugosidad con panel sensorial: (1 = extremadamente seco; 4 = ligeramente seco; 5 = ligeramente jugoso; 8 = extremadamente jugoso). (h) Determinación de sabor con panel sensorial: (1 = extremadamente soso; 4 = ligeramente soso; 5 = ligeramente intenso; 8 = extremadamente intenso). (i) Determinación del marmoleo según Australian Meat. (j) Determinación de la capacidad de retención de agua fue el porcentaje de pérdida de peso de la muestra de carne durante la congelación y la cocción. (k) Determinación de marmoleo con panelista bajo la observación de grasa intramuscular en fotografía y clasificado en 1 a 5 con intervalos de 0.5 (1 = sin grasa intramuscular visible, 5 = cantidad media de grasa intramuscular ya que no se encontró ningún individuo con alto nivel de marmoleado en el grupo actual de toros jóvenes). (l) determinación de la capacidad de retención de agua considerando el valor de pH a las 48 h post mortem. (m) Determinación de marmoleo según estándar coreano (1 = traza, 7 = muy abundante, considerando el grado de veteado en la superficie de corte de la carne). (n) según el criterio del estándar de corte GB/T17238-1998 de carne de res fresca y refrigerada de EE. UU. (U.S. Standard Publishing House). (NS) no significativa. (rs) Números de referencia de SNP identificados utilizando Ensembl versión 70 (Wellcome Trust Genome Campus - Hinxton, Reino Unido), (*) ID de base de datos dbSNP del NCBI. (rs) Código de SNP
Conclusiones
SNPs de CAPN1 y CAST se asocian positivamente, pero también de manera negativa con indicadores de calidad de carne. La información publicada y disponible sobre las asociaciones de SNPs con indicadores de calidad de la carne revela que, el estudio de marcadores moleculares está permitiendo mayores avances en el entendimiento de los atributos sensoriales de la carne.
El uso de marcadores moleculares para la identificación de regiones polimórficas del genoma bovino que estén asociadas a características deseables, podría brindar información del animal deseado, basado en propiedades fisicoquímicas de la carne, lo que permitirá la evaluación de la carne en lotes completos de animales. Sin embargo, no siempre es posible lograr animales con todas las características de calidad cárnica deseadas. La selección asistida con marcadores mucho más efectivos podría lograr controlar ciertos rasgos característicos de la canal y carne, pero también podría tener consecuencias no deseadas en otros rasgos de calidad.
Con este estudio proporcionamos detalles importantes de los SNP de genes CAPN1 y CAST que subyacen a los rasgos económicamente importantes para la industria de la carne bovina para mejorar la eficiencia de la producción y calidad de la carne.

