










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkProducción + Limpia
Print version ISSN 1909-0455
Rev. P+L vol.8 no.2 Caldas July/Dec. 2013
Artículo de revisión / Review article / Artigo de revisão
Exergía en sistemas biológicos: Aproximación holística para el estudio de ecosistemas y el manejo ambiental*
Exergy in biological systems: a holistic approach to the study of ecosystems and environmental management
Exergia em sistemas biológicos: aproximação holística para o estudo de ecossistemas e o manejo ambiental
Carlos Federico Álvarez Hincapié**, Héctor Iván Velásquez Arredondo***
* Artículo derivado del proyecto de investigación "Diversidad como capital natural: mariposas, información molecular y análisis termodinámico en bosques altoandinos (2010-2015)", financiado por la Corporación Universitaria Lasallista y la Universidad Nacional de Colombia sede Medellín.
** MSc. Docente Corporación Universitaria Lasallista, Doctorado en Ecología Universidad Nacional de Colombia, sede Medellín - Colombia. E-mail: carlosfederico.lasallista@gmail.com.
*** PhD. Docente Universidad Nacional de Colombia, sede Medellín - Colombia. E-mail: hivelasq@unal.edu.co.
Artículo recibido: 03/06/2013; Artículo aprobado: 30/10/2013
Resumen
Los sistemas ecológicos operan a partir de las interacciones entre elementos bióticos y abióticos, lo que incluye flujos de masas, de energía y de información. La exergía es el componente de la energía que puede desarrollar la máxima capacidad de trabajo con respecto a un estado de referencia del ambiente. Su aplicación en el estudio de los ecosistemas permite desarrollar teorías, modelos y aproximaciones basadas en la termodinámica, que pueden incrementar el poder predictivo y analítico de las ciencias ecológicas y ambientales. Esta revisión presenta una sinopsis del concepto de exergía, así como de algunas aplicaciones en estudios ecológicos y ambientales.
Palabras clave: biodiversidad, conservación, energía, ecología, ecosistema, exergía gestión ambiental.
Abstract
Ecologic systems operate from the interaction of biotic and abiotic elements, including masses, energy and information flows. Exergy is the component of energy that can develop a maximum work capacity with respect to a reference state in the environment. Its application in the study of ecosystems allows the development of theories, models and approaches based on thermodynamics, thus increasing the prediction and the analysis power of environmental and ecological sciences. This revision introduces a synopsis of the concept of exergy and some applications on ecological and environmental studies.
Key words: biodiversity, conservation, energy, ecology, ecosystem, exergy environmental management.
Resumo
Os sistemas ecológicos operam a partir das interações entre elementos bióticos e abióticos, o que inclui fluxos de massas, de energia e de informação. A exergia é o componente da energia que pode desenvolver a máxima capacidade de trabalho com respeito a um estado de referência do ambiente. Sua aplicação no estudo dos ecossistemas permite desenvolver teorias, modelos e aproximações baseadas na termodinâmica, que podem incrementar o poder previsível e analítico das ciências ecológicas e ambientais. Esta revisão apresenta uma sinopse do conceito de exergia, bem como de algumas aplicações em estudos ecológicos e ambientais.
Palavras importantes: biodiversidade, conservação, energia, ecologia, ecossistema, exergia, gestão ambiental.
Introducción
Los ecosistemas son sistemas ubicados en un área determinada, donde interactúan organismos bajo la influencia continua de factores abióticos. A pesar de los diversos avances en la ciencia ecológica, aún persisten múltiples interrogantes sobre el funcionamiento de los ecosistemas y las formas de gestión sustentables de los mismos. Jørgensen y Svirezhev (2004) anotan que, dado que un ecosistema es un sistema muy complejo, no es una tarea fácil encontrar buenos indicadores ecológicos para dar información sobre su integridad. Diversas funciones objetivo (también denominadas principios extremos) han sido propuestas como reguladoras globales de la evolución de los ecosistemas, como aquellas asociadas al aumento en la distancia al equilibrio termodinámico, incluyendo la exergía (Gaucherel, 2006), la cual puede entenderse como la máxima cantidad de trabajo que puede desarrollar un sistema en relación con un estado de referencia del ambiente (Fath, Jørgensen, Patten & Straškraba, 2004).
Los sistemas biológicos, desde una célula, pasando por un organismo, una población, una comunidad, hasta un ecosistema, son sistemas abiertos, en continua interacción con su ambiente. Este puede considerarse a partir de cinco componentes o elementos en interacción: la biósfera, la litósfera, la atmósfera, la hidrósfera y la sociósfera. La exergía solar se consume en los flujos constantes de materia, energía e información que se dan dentro y entre estos componentes (Wall & Gong, 2001). Ahora bien, aún desconocemos con precisión la cuantificación de los flujos y reservorios energéticos en los ecosistemas y en los sistemas socio-ecológicos complejos, y existen limitantes en la información y modelamiento de los ecosistemas, especialmente en países tropicales (donde la diversidad biológica es abundante y altamente amenazada). El análisis de las redes tróficas (redes alimenticias: consumo de productores-plantas-algas por parte de consumidores primarios, secundarios y detritívoros / descomponedores) sugiere que a partir de los flujos de energía solar hacia los productores primarios y la posterior transferencia (de materia y energía) a consumidores y detritívoros, es posible el desarrollo de la vida. Desde tiempo atrás, se ha señalado que la única forma de que un sistema vivo se mantenga como tal (lejos del estado inerte de máxima entropía) es extrayendo entropía negativa de su medio ambiente; de esta forma un organismo se mantiene en un nivel elevado de orden (baja de entropía) absorbiendo continuamente orden de su medio ambiente (Schrödinger, 1944).
Jørgensen y Fath (2004a) proponen el uso de los principios termodinámicos para explicar el crecimiento y desarrollo de los sistemas ecológicos. En general el crecimiento significa un incremento en el tamaño del ecosistema, mientras que desarrollo es un incremento en la organización, independiente del tamaño. Termodinámicamente, el crecimiento y el desarrollo significan alejarse del equilibrio termodinámico, en donde el sistema no puede desarrollar ningún trabajo, sus componentes son inorgánicos, tienen cero exergía y los gradientes son eliminados.
El crecimiento se puede dar en un ecosistema a través de entradas (energía de baja entropía o biomasa), de crecimiento estructural (biomasa en el sistema al aumentar la cantidad, tamaño o número de sus componentes), del crecimiento de redes (componentes, conexiones, retroalimentaciones, ciclaje y flujos totales) y por medio del crecimiento en información (eficiencia energética asociada a incrementos en la complejidad genética) (Fath, Jørgensen, Patten & Straškraba, 2004). En este sentido, las variaciones de exergía expresan cambios en la estructura y componentes ecosistémicos (Fonseca, Marques, Paiva, Freitas, Madeira, Jørgensen, 2000). Si un sistema recibe una entrada de exergía, la utilizará para mantenerse lejos del equilibrio termodinámico, y parte de esta se destruirá por irreversibilidades inevitables del sistema. Si después del gasto de energía para el mantenimiento, todavía hay exergía disponible, entonces el sistema se alejará del equilibrio termodinámico, lo que se reflejará en el aumento de gradientes (Jørgensen y Fath, 2004a).
Este artículo presenta inicialmente una sinopsis introductoria al concepto de exergía y diversas aproximaciones al mismo. En la siguiente sección se muestra una síntesis de la aproximación de Jørgensen y colaboradores al concepto de eco-exergía, como un referente en cuanto a la exergía contenida en la biomasa y en la información de los organismos. A continuación, se presenta la aplicación de la exergía en el estudio de ecosistemas, el manejo ambiental y de recursos naturales, para finalmente señalar conclusiones y posibles líneas de trabajo futuro.
EXERGÍA: una sinopsis introductoria
El concepto de exergía tiene sus raíces en los trabajos de Carnot a inicios del siglo XIX asociados a la eficiencia de las máquinas de vapor. La exergía es un potencial termodinámico, es una medida general de trabajo, una "diferencia" o un contraste, es la parte de la energía que es posible convertir en trabajo (Wall & Gong, 2001). Una relación entre los conceptos de energía y exergía pueden apreciarse en la tabla 1.
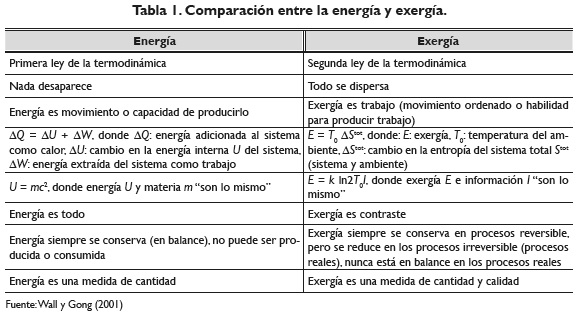
La exergía es también información o capacidad de información termodinámica acumulada en un sistema (en comparación a su punto de equilibrio termodinámico). Boltzman, en 1905, postuló que la energía libre de información es kTlnI, donde I es la información del sistema, T es la temperatura y k es la constante de Boltzman, aunque la exergía como información es usualmente una parte muy pequeña de la exergía total del sistema (Wall & Gong, 2001).
La entropía de un sistema cerrado aumenta continua e irrevocablemente, o sea, la energía disponible (exergía) es transformada continuamente en energía no disponible hasta que desaparece completamente, aunque algunos organismos (plantas) retrasan la degradación entrópica, almacenando parte de la radiación solar que sin ello se disiparía inmediatamente en forma de calor, en alta entropía (Georgescu, 1975). Los organismos, en aras de mantener su baja entropía constante, se alimentan constantemente de baja entropía de su entorno (Georgescu, 1971)9. El crecimiento y desarrollo ecológico organiza materiales en estructuras ordenadas y se requiere de trabajo para generar orden por fuera del "background" con menor orden (Jørgensen y Fath, 2004a).
En los sistemas biológicos la exergía incluye tanto la energía que puede ser convertida en trabajo (almacenada en la biomasa) como la información biológica que separa a los organismos de su entorno (Jørgensen y Fath, 2004a). La exergía refleja el grado de desarrollo o complejidad en un ecosistema, y su crecimiento y desarrollo son el resultado de un sistema de componentes y configuraciones que maximizan el flujo de energía útil y la cantidad de exergía almacenada (Silow & Mokry, 2010).
Por otra parte, los stocks en la forma de materia orgánica viva y muerta en la tierra representan formas diferentes de exergía almacenada. Este almacenamiento de exergía genera un potencial de exergía de crucial importancia para la evolución de la vida en diversidad y complejidad (Wall & Gong, 2001). En una red trófica, la biomasa y la exergía fluyen entre los compartimientos del ecosistema, soportando procesos por los que la exergía se degrada y se almacena en biomasa, en distintos niveles tróficos (Salas, Marcos, Pérez & Marques, 2005).
La exergía puede expresarse de formas diferentes. Se recomienda revisar el recuento histórico del concepto y sus aplicaciones presentado por Sciubba y Wall (2007), así como la relación que presentan sobre los diferentes componentes de la exergía y los flujos de energía. Diversas formulaciones matemáticas han sido presentadas en relación con el concepto, incluyendo a Gibbs (1928) citado por Wall (1993), Wall (1977), Szargut, Morris & Steward (1988), Wall y Gong (2001), Silow & In-Hye (2004), Jørgensen, Ladegaard, Debeljak, & Marques (2005), Sciubba y Ulgiati (2005), Sciubba, Bastianoni & Tiezzi (2008), Sciubba (2009), Jørgensen (2009). Estas se presentan en la tabla 2.
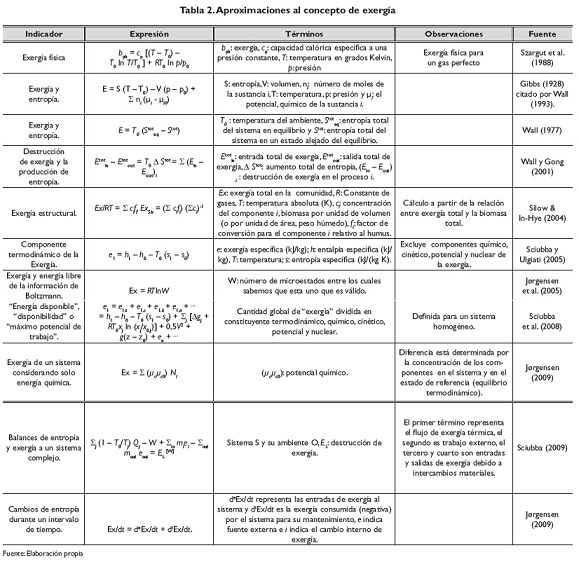
La exergía disponible para el hombre viene del stock de energía en los depósitos minerales y del flujo de radiación solar interceptado por la tierra (Georgescu, 1971). Dado que para la evolución de la naturaleza y de la vida los depósitos de exergía son esenciales, ellos representan una medida del valor del sistema y un indicador de sostenibilidad, ya que su reducción lleva a la destrucción de los sistemas de soporte de vida en la tierra (Wall & Gong, 2001).
Aun cuando para el cálculo de la exergía en sistemas ecológicos se ha propuesto el detritus como estado de referencia del ambiente (ver siguiente sección), otra posibilidad es hacerlo tomando áreas de control (Libralato, Torricelli & Fabio, 2006) (libres de perturbación), así:
Ecuación 1: Ext = ExIt - ExEt = RT ∑βt (CIi, t - CEi, t)
Donde: ExIt: Exergía interior de la comunidad-sistema; ExEt: Exergía externa a la comunidad-reservorio; CIi, t: Concentración de biomasa en la i-esima especie en la comunidad perturbada y CEi, t: Concentración de biomasa en la i-esima especie en la comunidad de control.
Ahora bien, los análisis de exergía estrictamente hablando solo aplican a sistemas que se mueven por una ruta en equilibrio (donde es posible dividir el sistema en subsistemas y cada uno de estos, si es aislado, asume un estado de equilibrio). Sin embargo, estos análisis proveen una suficiente aproximación a procesos en sistemas reales. En la termodinámica del equilibrio, la energía libre de Gibbs y la exergía juegan un papel análogo para conocer los cambios en el sistema, así como lo hace la producción de entropía en la termodinámica de no equilibrio (Ruth, 1995).
En síntesis, el concepto de exergía se asocia a la energía útil en un sistema, permite el crecimiento y desarrollo de los ecosistemas, y representa un amplio potencial en estudios ecológicos y ambientales. A continuación se presenta con más detalle la propuesta de eco-exergía de Jørgensen y colaboradores, referente obligado frente al análisis exergético en ecosistemas.
Eco-EXERGÍA: la propuesta de Jörgensen y colaboradores
Sven Erik Jørgensen, de la Universidad de Copenhagen, ha desarrollado una propuesta del uso de la exergía enriquecida a lo largo de las últimas décadas por trabajos con diversos colaboradores. En esta sección se presentan algunos elementos relevantes de su aproximación que, si bien no ha sido exenta de críticas (Sciubba, 2009; Sciubba y Wall, 2007; Christensen, 1995;Wagendorp; et al. 2006; Dewulf; et al. 2008; Gaudreau; et al. 2009) y presenta limitantes implícitos (Salas; et al. 2005; Jørgensen; et al. 2005; Jørgensen, 2009), contiene elementos de utilidad para la conservación de la biodiversidad y el manejo ambiental.
La exergía es una medida de la energía de un sistema en relación con su ambiente, y no se conserva en el sistema, sino que gradualmente se pierde en los procesos, desde la fijación por parte de los productores primarios hasta su destino final (Christensen, 1995). Es una medida de qué tan lejos está el ecosistema del equilibrio termodinámico, esto es, qué tanta organización del ecosistema ha sido creada en la forma de organismos, compuestos orgánicos o redes ecológicas complejas (Jørgensen, 2009). La exergía ha sido propuesta como una propiedad holística de los ecosistemas (Jørgensen, 1992) y, dado que aumenta con el incremento de biomasa, el desarrollo de la red ecológica y el crecimiento de la información, esta sirve como indicadora de la salud ecosistémica (Jørgensen, 2008a; Jørgensen, 2008b); y permite hacer comparaciones con ecosistemas similares (Jørgensen, 2006-2007).
Jørgensen y Mejer (1977) propusieron que la exergía podía usarse como una expresión de la capacidad ecológica buffer. Mejer y Jørgensen (1979) presentaron la estimación ecológica de exergía en términos de una suma global de los componentes de un ecosistema, donde se toma en cuenta la concentración relativa del componente correspondiente y su "distancia" de un estado de referencia (el mismo componente en el equilibrio termodinámico), proponiendo:
Ecuación 2: Ex = RT ∑ [ci ln(ci/cieq)- (ci - cieq)]
Donde: cieq = Ci0. R: constante de gases, T: temperatura absoluta, ci: concentración en el ecosistema del componente i, el índice 0 indica los componente inorgánicos del elemento químico en consideración y cieq: es la concentración del componente i en equilibrio termodinámico.
Una formulación similar es presentada por Christensen (1995) asi:
Ecuación 3: EX = RTC0 [ln (C0/Co, ref) - (l-C0, ref / C0) + ∑ xi ln(xi / xi, ref)]
Donde C0: concentración total de todos los grupos, C0, ref: concentración de referencia de todos los grupos, xi: proporción Ci / C0 y xi, ref: proporción Ci, ref / C0, ref
Se han sugerido cálculos que consideran la organización de los organismos y sus mayores contribuciones a la exergía al sistema bajo análisis. A partir de un estado de referencia (detritus o materia orgánica muerta), se estima la exergía en términos de la probabilidad (Pi) de producir materia orgánica y la probabilidad de "seleccionar" información genética correspondiente (Pia), así:
Ecuación 4: Ex ≈ -RT ∑ ci ln (Pi)
Donde (Pi)= PI*Pia, PI = [cI/c0eq] exp [-(μI-μIeq) / RT], μI = μIeq + RT ln cI/cIeq, μ: indica el potencial químico, Pia = 20-700 g, ci: concentración del componente i, 20: aminoácidos esenciales de los organismos, g: número de genes y 700: promedio de aminoácidos codificados en un gen de la especie i (Jørgensen, Nielsen & Mejer, 1995).
La exergía se basa en la distribución de la materia o de los nutrientes entre los diferentes compartimientos de un ecosistema (Christensen, 1995). En este sentido, la aproximación requiere estimaciones, como la cantidad de biomasa (ci).
Ecuación 5: Ex / R T ≈ ∑βi ci
Donde ci: concentración de biomasa de la especie i y βi: factor de ponderación que expresa la "calidad de información" contenida en la biomasa (Jørgensen & Nielsen, 1998).
Para Jørgensen y Bendoricchio (2001) la exergía total y la exergía estructural de una comunidad pueden hallarse así:
Ecuación 6: Ex = ∑ ci ln fi
Ecuación 7: ExStr = (∑ cifi) (∑ ci)-1
Donde Ex: Exergía total de la comunidad, ExStr: Exergía estructural de la comunidad, ci: concentración del componente i, fi: factor de conversión del componente i.
El cálculo de la exergía puede hacerse a partir de potenciales químicos (dominantes en los ecosistemas), así:
Ecuación 8: Ex = RT ∑ CilnCi/Ci0
Donde: 0: todos los compuestos inorgánicos, n=1: detritus, i≥2: organismos, Ci: concentración del i-esimo componente en el ecosistema, Ci0 el mismo componente en equilibrio termodinámico (Jørgensen et al., 2005).
La exergía específica ha sido definida como la eco-exergía dividida por la biomasa, y ha sido utilizada como un indicador holístico, aunque es necesario complementar su uso con otros indicadores (Jørgensen, 2008a). En este mismo sentido, Molozzi, Salas, Callisto, Marques (2013) indican que aunque la eco-exergía y la eco-exergía específica son indicadores efectivos para identificar la calidad de reservorios de agua, estos deben ser usados complementariamente y se debe combinar su uso conjuntamente con otros índices bióticos (como los de diversidad) y abióticos (asociados a las características físico-químicas).
Jørgensen (2009) señala que para el cálculo de la eco-exergía de un sistema completo se pueden combinar la exergía química y la exergía biológica, así:
Ecuación 9: Ex total = 18.7 ∑ ci 0, 0529 ∑ cigi [ML-1T-1].
Donde: g para el detritus (i=1) es 0.
Igualmente, este autor señala que es posible introducir un factor de ponderación β, para convertir la exergía de varios organismos en una unidad de detritus equivalente o equivalente de exergía química:
Ecuación 10: Ex total = ∑βi ci
Donde: ci: Concentración del componente i y βi: Información contenida en la biomasa.
De esta forma, el cálculo de la eco-exergía considera la energía química incorporada en la biomasa, así como la información genética contenida en el organismo (Jørgensen, 2009). Es posible encontrar aproximaciones a estos valores β en Marques, Pardal, Nielsen & Jørgensen (1997, p. 159); Jørgensen, Patten, Straškraba, (2000, p. 258); Fonseca et al. (2000, p. 185); Jørgensen, Odum & Brown (2004, p. 13); Jørgensen et al. (2005, p. 170, 173); Salas et al. (2005, p. 2512); Jørgensen (2009, p. 134). En cualquier caso, el detritus como punto de referencia tiene un valor β=1 Fonseca et al. (2000), y su exergía es 18, 7 kJ/g (Jørgensen, Odum & Brown, 2004).
Se ha propuesto la información genética como un estimador de β, como discriminador de la organización con respecto al estado de referencia del detritus. Aun cuando inicialmente se propuso usar el número de genes para determinar β (Jørgensen, Nielsen & Mejer, 1995), se encontró que la información disponible era insuficiente y se generaban estimados muy gruesos por lo que se adoptó al contenido de ADN del organismo (valor C / 2C) para determinar β (Marques et al., 1997).
El cálculo de los coeficientes β a partir del contenido del valor C (contenido de ADN), se puede hacer mediante (Libralato, et al. 2006):
Ecuación 11: βi = ln (20 -(Ci/2)(bp/3))I/ Exdet
Donde Ci: valor C; bp = 9, 8 * 108; Exdet: 7, 43 * 105
Jørgensen (2009) señala que la eco-exergía total de un ecosistema no se puede calcular exactamente, ya que no podemos medir las concentraciones de todos los componentes o determinar todas las posibles contribuciones de exergía al ecosistema. Dadas estas y otras limitaciones, se considera la eco-exergía como un índice relativo mínimo de la exergía (Jørgensen, 2006-2007). Gaudreau et al. (2009) anotan que las limitantes clave de la exergía que deben ser superadas incluyen:la incompatibilidad entre la calidad de la exergía y la calidad del recurso, la inhabilidad de la exergía para caracterizar recursos que no producen trabajo a través de la concentración de exergía, los limitantes asociados a la derivación de la exergía y los problemas para definir el ambiente de referencia. Aunque Jørgensen y Fath (2004b) proponen el uso de la exergía para modelar la selección adaptativa de los organismos, señalando que la maximización de la exergía puede considerarse como un orientador general de los procesos evolutivos, independientemente de los mecanismos subyacentes y de las rutas moleculares usadas para alcanzar el estado final, este es un punto controversial y discutible.
Jørgensen y Fath (2004a) han desarrollado una propuesta de ocho leyes generales de la ecología desde las implicaciones termodinámicas de procesos ecosistémicos en macroescala, destacándose a la luz del objeto de este artículo los siguientes: (1) Los ecosistemas son sistemas abiertos embebidos en un contexto de donde reciben entradas de energía-materia y en donde descargan salidas de energía-materia. Si un sistema recibe una entrada de exergía, la utilizará para desarrollar trabajo manteniendo el sistema lejos del equilibrio termodinámico, y si hay suficiente exergía se dará un aumento de los gradientes;(4) La masa (incluyendo la biomasa) y la energía se conservan;(7) Los procesos ecosistémicos son irreversibles. Los organismos necesitan energía para cubrir el mantenimiento de los procesos vitales; (8) Los procesos biológicos usan la energía capturada para alejarse del equilibrio termodinámico y mantener un estado de baja entropía relativo a su entorno. Después de una captura inicial de energía, el crecimiento y desarrollo de los ecosistemas es posible por un incremento en la estructura física (biomasa), incremento en las redes (más ciclaje) o por un incremento en la información.
Como síntesis de este apartado, la propuesta de Jørgensen y colaboradores parte de la evaluación de la biomasa y del contenido de ADN de los organismos para evaluar la exergía a partir del gradiente existente en comparación con el detritus. A continuación se presenta el concepto de exergía con el estudio de ecosistemas, se referencian trabajos desarrollados en el campo y desde una perspectiva ampliada, se hace una introducción a su aplicación al manejo ambiental y de los recursos naturales.
Aplicaciones en el estudio de ecosistemas, el manejo ambiental y los recursos naturales
Exergía y ecosistemas: elementos globales
Los mayores avances del uso de la exergía en sistemas ecológicos se han dado en ecosistemas acuáticos, posiblemente debido a factores metodológicos asociados a la medición de la biomasa en sistemas relativamente cerrados o a tendencias de investigación asociadas a escuelas académicas. Sean cuales fueren las causas, de esto resalta la importancia de avanzar en trabajos de campo, teóricos o de modelación, orientados a una mayor exploración de esta aproximación en ecosistemas terrestres (incluyendo organismos modelo, así como comunidades-ensambles completos).
Para conocer las características generales de los ecosistemas y para validar los modelos, es necesario conocer y combinar componentes estructurales y funcionales de los mismos (Müller, 2005). La evaluación de la exergía almacenada en los ecosistemas es un acercamiento a ambos aspectos, de ahí su utilidad como indicador ecológico y para el modelamiento (Jørgensen & Svirezhev, 2004). Así, estudios que relacionen la estructura de los ecosistemas con la capacidad de procesamiento de energía e información resultan de interés (Fonseca et al. 2000).
El crecimiento estructural (biomasa) se relaciona con la energía excedentaria disponible, aunque existen límites dados por las características de cada entorno y no es posible un crecimiento ilimitado ni en biomasa ni en complejidad (Anillo y Barrio, 2006). La equidad ("evenness") constituye una parte de la capacidad de información de los sistemas. Cuando se manipula la información, aumentando la equidad, se crea una estructura disipativa más eficiente, optimizando la captura de exergía e incrementando su almacenamiento en la biomasa (Jørgensen & Fath, 2004a).
Los ecosistemas se desarrollan para degradar los gradientes de energía más efectivamente, pues estructuras disipativas más desarrolladas degradan más energía. Por tanto, se espera que ecosistemas más maduros degraden la exergía de la energía que capturan de una forma más completa que los ecosistemas menos desarrollados (Schneider & Kay, 1994). En este mismo sentido se ha señalado que la proporción de generación de entropía / entropía acumulada (disipación específica) puede considerarse como indicador de la habilidad de autorganización de un sistema para mantenerse lejos del equilibrio. A mayor proporción, menor la capacidad del sistema para convertir entradas de energía en organización interna (Ludovisi, Pandolfi & Taticchi, 2005). A medida que los ecosistemas crecen y se desarrollan, deben incrementar su disipación total, desarrollar estructuras más complejas con más flujo de energía, incrementar la actividad de ciclaje, desarrollar mayor diversidad y generar más niveles jerárquicos, todo asociado a la degradación de energía (Schneider & Kay, 1994).
Desde otra perspectiva, Doka, Hillier, Kaila, Köllner, Kreiβig, Muys, Garcia-Quijano, Salpakivi-Salomaa, Schweinle, Swan & Wessman (2002) afirman que los ecosistemas construyen altos niveles de exergía y, como consecuencia, son capaces de disipar los flujos entrantes de energía. Así, es posible medir directamente el nivel de exergía de un ecosistema asociado a los regímenes de uso del suelo comparado a un ecosistema de referencia con las mismas condiciones locales. Igualmente es posible medir la exergía indirectamente, al medir la efectividad en la disipación ("dissipation/buffering performance") del ecosistema bajo regímenes de uso en comparación con un sistema de referencia.
El nivel de exergía de un ecosistema está generalmente referido a la complejidad o integridad del mismo; su componente de biomasa incluye la cantidad total de biomasa y su ubicación entre los compartimientos vivos, muertos y su capacidad de recuperación. Asimismo, su componente de información incluye la biodiversidad alfa, beta y gama, así como la red trófica (Doka et al., 2002).
Dewulf et al., (2008) revisan el potencial y limitaciones del concepto de exergía en el análisis: (1) de ecosistemas, describiendo el máximo almacenamiento y disipación de flujo de energía; (2) de sistemas industriales, desde procesos individuales hasta procesos encadenados; (3) termo-económico, con contabilidad extendida de exergía, y (4) de evaluación de impacto ambiental, a través del ciclo de vida completo con cuantificación de los efectos de la captura de recursos y la generación de emisiones.
Zhang, Gurkan & Jørgensen (2010) presentan una revisión de la aplicación de la eco-exergía en la evaluación de la salud de ecosistemas y en el desarrollo de modelos estructuralmente dinámicos. Por su parte, Gaucherel (2006), empleando una modelación tipo "Daisy world", demostró que los patrones espaciales de los ecosistemas parecen estar altamente correlacionados con la exergía y su evolución, anotando que se requiere considerar los efectos de la configuración espacial para lograr estimaciones más acertadas sobre aspectos relacionados con la exergía.
Exergía y estudio de ecosistemas
Silow y Mokry (2010) presentan una síntesis sobre la correlación entre índices de exergía y otros indicadores del estado de los ecosistemas. Los aspectos del ecosistema cubiertos por la eco-exergía específica como indicador incluyen: el tamaño de la red y eficiencia en el uso de la exergía, así como la información transportada por los organismos y también los efectos de la regulación y sinergia entre los organismos (Jørgensen, 2006). El análisis exergético ha sido empleado, por ejemplo, por Sciuba y Zullo (2012) que trabajaron en el análisis y modelamiento de relaciones depredador-presa.
A una escala más amplia, en el estudio de la exergía asociada a los ecosistemas y a comunidades bióticas, se han empleado diversas aproximaciones, incluyendo el índice de exergía (Salas et al., 2005; Libralato et al., 2006; Marques et al., 1997; Xu, 1997; Xu, Jørgensen & Tao, 1999; Ludovisi & Poletti, 2003; Marques, Nielsen, Pardal & Jørgensen, 2003; Fath & Cabezas, 2004; Munari & Mistri, 2006; Park, Lek, Scardi, Verdonschot & Jørgensen, 2006; Patricio, Marcos, Pardal, Pérez, Marques & Salas, 2006b; Patrício, Neto, Teixeira, Salas & Marques, 2009; Pusceddu & Danovaro, 2009), la exergía específica (Salas et al., 2005; Molozzi et al., 2013; Marques et al., 1997; Ludovisi & Poletti, 2003; Marques et al., 2003; Pusceddu & Danovaro, 2009; Fabiano, Vassallo, Vezzulli, Salvo & Marques, 2004; Patricio, Salas, Pardal, Jørgensen & Marques, 2006a; Austoni, Giordani, Viaroli & Zaldívar, 2007), la exergía estructural (Silow & In-Hye, 2004; Xu, 1997; Xu et al., 1999; Xu, Dawson, Tao, Li & Cao, 2002; Xu, Wang, Chen, Qin, Wu, He & Wang, 2011; Xu, Zhang, Huang & Su, 2012), la proporción exergía/flujo de emergía (Bastianoni & Marchettini, 1997, Bastianoni, Pulselli & Rustici, 2006), los componentes exergéticos (Zaleta, Ranz & Valero, 1998), el principio de máxima exergía (Marques & Jørgensen, 2002) y la exergía química (Chen & Ji, 2007). En la tabla 3 se referencian con detalle estas aplicaciones incluyendo el tipo de índice, el objeto de estudio, así como resultados y observaciones.
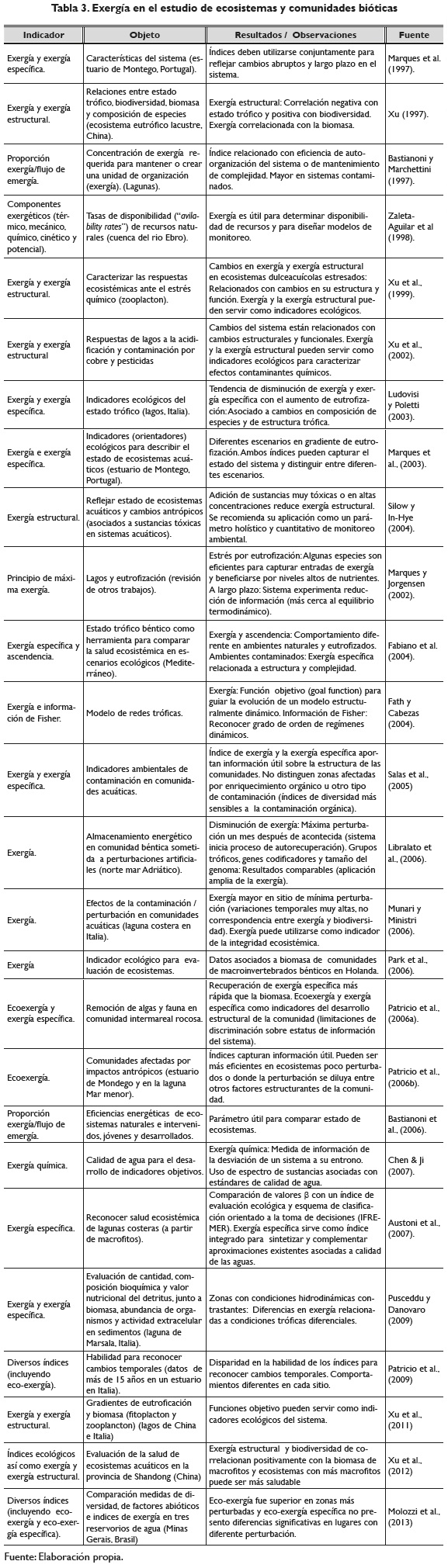
Ahora bien, es necesario considerar como lo señala Jenssen (2007), que la conversión de la diversidad de especies en un término exergético debería considerar explícitamente la estructura genética de las diferentes especies y sus poblaciones. En este sentido, un punto adicional a considerar está en la posibilidad de emplear otras fuentes de información diferentes al contenido de ADN para determinar el factor β. Por ejemplo, las filogenias obtenidas a partir de varios genes o, idealmente, a partir del genoma completo de las especies representan una fuente de información evolutiva, que no está siendo considerada por el valor C empleado desde la propuesta de Jørgensen y colaboradores. Asumir que el contenido total de ADN representa la información obtenida a través de los procesos evolutivos puede constituirse en un reduccionismo extremo, que desconoce aspectos fisiológicos, comportamentales y filogenéticos que hacen parte de la propia evolución y de la información de la que son portadores los organismos que hacen parte de los ecosistemas.
Exergía en el manejo ambiental y de recursos naturales
En el campo del uso de la exergía como indicador ambiental una referencia obligada es el trabajo de Silow y Mokry (2010), quienes presentan una amplia revisión del uso de la exergía en el modelamiento y el monitoreo, ambiental particularmente en ecosistemas acuáticos, y con referencia ampliada al estudio de caso del ecosistema del lago Baikal.
Los indicadores basados en la exergía pueden ayudar a conocer la conexión entre la calidad de los suministros y la demanda de energía, la ubicación y magnitud de los puntos de degradación de energía, el impacto ambiental asociado a producir, reutilizar y reciclar materiales de construcción, así como limitantes tecnológicas de los sistemas productivos, sus fuentes energéticas y las necesidades de avances, para el manejo de los sistemas complejos (Koroneos, Nanaki & Xydis, 2012).
La exergía proporciona un valor físico consistente sobre la energía y los recursos materiales con respecto al ambiente, que puede convertirse en un suplemento a los valores monetarios (Gong & Wall, 2001), y es un parámetro útil para reflejar el estado de los ecosistemas y estimar la severidad de los daños antropogénicos (Silow & Mokry, 2010). Las actividades humanas pueden disminuir el nivel de exergía de los ecosistemas a partir de la simplificación, la reducción de biomasa, la destrucción de la complejidad interna, la pérdida de especies y la pérdida subsecuente de funcionalidad ecosistémica (Wagendorp et al., 2006). Todos los procesos en la realidad son irreversibles, lo que significa que la exergía se pierde y se produce entropía, dos descripciones diferentes de la misma realidad (Jørgensen, 2009). La destrucción de exergía debido a las irreversibilidades se puede relacionar con la creación de caos, o la destrucción de orden en los sistemas organizados y el aumento de desorden (por la dispersión de contaminantes y pérdida de organización de los sistemas vivientes), y pude correlacionarse con el impacto ambiental (Rosen, 2002). Ahora bien, es importante considerar que para computar la destrucción de exergía en sistemas complejos, se necesita contabilizar apropiadamente las complejas interacciones entre sus componentes, por lo que las respuestas del sistema ante las variaciones de sus sub-unidades no siempre son fácilmente predecibles (Sciubba, 2009).
En el caso de la provisión de bienes y servicios ecosistémicos, Tundisi et al. (2012) sugieren la posibilidad que si la exergía expresa biomasa e información, ecosistemas hídricos más diversos y estables tendrán una mayor exergía y disponibilidad para ofrecer diversidad y valor de servicios ecosistémicos para el bienestar humano.
Asumiendo a las leyes físicas como herramientas objetivas para conocer los costos del agua, la hidronomía física ("physical hydronomics", PH) se ha desarrollado como una herramienta contable de base termodinámica para caracterizar físicamente la degradación de cuerpos de agua a través del cálculo de la pérdida de exergía. Con la aplicación en dos cuencas en España, esta aproximación ha sido validada, aunque los datos requeridos para la aplicación de PH son específicos y el uso de información secundaria es necesario, lo que lleva a algún nivel de imprecisión (Martínez, 2009).
El análisis energético puede usarse conjuntamente con otros indicadores biofísicos para analizar el impacto de actividades y obras de restauración ingenieril (Ma, Zhang, Zhang & Li, 2012). Aunque cada evaluación de un recurso natural requiere de un marco de análisis particular, el procedimiento general incluye: (1) identificar las características más relevantes del recurso y obtener una caracterización física y química, que lo diferencia de su entorno, (2) seleccionar el estado de referencia más adecuado y (3) calcular la exergía (Martínez, 2009).
Desde una perspectiva amplia, los flujos de exergía recibidos del sol y sus depósitos permiten el desarrollo de las actividades de las sociedades humanas en el planeta (Wall, 1977), aunque el aprovechamiento directo de la exergia proveniente del sol es muy pequeño en comparación con el uso que se hace de los depósitos. Así, el contenido de exergía de la entrada de recursos naturales a la economía puede ser interpretado como una medida general de su "utilidad" y, dado que el contenido de exergía en los desperdicios puede generar perjuicios al llevar a reacciones incontroladas en el ambiente, su exergía puede ser una medida del daño potencial. En este sentido, los diagramas de uso de exergía pueden servir como indicadores ecológicos asociados a la eficiencia de los procesos y a las consecuencias del uso de los recursos (Gong & Wall, 2001). Dichos diagramas pueden utilizarse para analizar procesos específicos como el flujo de exergía en la producción de diversos tipos de combustibles (Gong y Wall, 2001), hasta problemas complejos como el uso de exergía en una sociedad (Gong y Wall, 2001). En este mismo sentido, Chen et al. (2009) propusieron un marco general para el análisis exergético, el cual es ilustrado para la agricultura China, mientras que Zhang et al. (2012) analizaron insumos y emisiones de la industria China.
Sciubba y Ulgiati (2005) señalan que los elementos más importantes de la exergía desde la perspectiva del análisis de los sistemas energéticos son:(1) Definir un estado de referencia constante en todos los cálculos; (2) Un sistema S en un estado A puede entregar una cantidad máxima de trabajo útil igual a su exergía, e inversamente, un sistema S en un estado 0 puede llevarse a un estado A mediante un gasto de trabajo mecánico superior a su exergía (para transformaciones irreversibles); (3) Si un sistema se mueve de un estado A a un estado B, el máximo trabajo extraíble es eA - eB.; (4) Existe un límite en la eficiencia de Carnot; (5) La exergía es aditiva en los componentes del sistema; (6) Es posible definir un "contenido acumulativo de exergía" del sistema a partir de la suma de todas las entradas; (7) Es posible asignar un contenido exergético a los flujos materiales y procesos.
El análisis exergético puede servir para reducir el uso de recursos (o exergía) necesarios para los procesos, lo que implica menor extracción de recursos del ambiente (Rosen, 2002). Así, el análisis exergético permite reconocer aspectos termodinámicos de procesos y ofrece indicaciones cuantitativas sobre las irreversibilidades y la correspondencia entre los recursos utilizados y los flujos materiales o de energía al final del uso (Sciubba y Ulgiati, 2005).
Sciubba (2009), propone el uso de la contabilidad exegética extendida (Extended exergy accounting), como un indicador del uso de recursos primarios en el ciclo de vida de un material o de bienes inmateriales. Igualmente se propone el análisis extendido de exergía (Extended exergy analysis, EEA) para obtener balances de exergía en los niveles de componentes y del sistema, llegando a conocer los costos exergéticos asociados a procesos productivos (Jørgensen, 2009). Este análisis fue empleado por Bligh y Ugursal (2012) para caracterizar la extracción, conversión y uso energético en Nova Scotia (Canada), y se encontró una amplia disipación de energía de alta calidad (pérdida de exergía) a través de las actividades económicas. A su vez, Seckin y Bayulken (2012) lo utilizaron para analizar el manejo de residuos sólidos.
Se evidencia que en el campo de la aplicación de la exergía en el estudio de los ecosistemas y en el manejo ambiental existen diversas aproximaciones y aplicaciones, que han ido acumulando cada vez más información, útil para el desarrollo de trabajos futuros asociados a una mejor comprensión y uso del concepto en la toma de decisiones económicas, de gestión y de conservación.
Conclusiones. Posibles líneas de Interés
A pesar de las diferentes aproximaciones y enfoques asociados al concepto de exergía, esta puede considerarse como el combustible de los sistemas disipativos, que se sostienen por conversión de materia y energía, como una célula, un organismo, un ecosistema, la tierra con sus flujos materiales o una sociedad (Wall, 1993). El comportamiento de este y otros índices de orientación biofísica, ante las actividades humanas sobre los ecosistemas, es un área de trabajo de gran interés, no solamente para las ciencias básicas como la ecología, sino también para las aplicadas como la economía ecológica, la biología de la conservación y la gestión ambiental. Mayores avances se requieren en el estudio de ecosistemas terrestres (incluyendo los agroecosistemas). En estos sistemas, los avances logrados en la medición de biomasa y composición de comunidades vegetales, así como modelos de la distribución de las especies, redes tróficas y diversidad en diferentes niveles representan puntos de partida a relacionar con el análisis exergético.
Un área de interés es el uso de sensores remotos con base térmica, para evaluar el impacto ambiental midiendo la función del ecosistema en términos de disipación de energía (Wagendorp et al., 2006). Esto puede permitir un escalamiento de los cálculos de exergía obtenidos con comunidades y áreas particulares, hacia escalas mayores.La exergía es una herramienta útil en el análisis de impacto, aunque se encuentra en un estado menos avanzado en el análisis de los sistemas ecológicos y en su conexión con el análisis económico (Dewulf et al., 2008), lo que amerita el desarrollo de exploraciones futuras y el uso de diversos orientadores termodinámicos para cuantificar el desarrollo de los sistemas ecológicos (Ludovisi et al., 2005). En este sentido, se requieren mayores avances en la integración de los índices exergéticos con otro tipo de información (como los indicadores de diversidad a diferentes niveles o los análisis de base económica). Se recalca que los índices de exergía tienen limitaciones como indicadores para el manejo ambiental y se requiere desarrollar más trabajos prácticos y el diseño de experimentos para probar marcos teóricos, así como mejorar la estimación de los factores de ponderación para calcular la exergía (Salas et al., 2005). El conocimiento paulatino de los contenidos totales de ADN para los diferentes organismos, así como la obtención y organización de la información derivada (filogenias) permitirán mayores avances en el conocimiento de ecosistemas acuáticos y terrestres, lo que, a su vez, se traducirá en más información para apoyar la toma de decisiones de manejo.A futuro, los análisis de comunidades a partir de filogenias obtenidas de genomas completos serán de gran utilidad para refinar los aportes de dicha información.
En el contexto de la entropía, todo proceso en la naturaleza da como resultado un déficit para el sistema en su conjunto. Así, cuando producimos una lámina de cobre reducimos la entropía (el desorden) del mineral, pero solamente al coste de un aumento de entropía en el resto del universo (Georgescu, 1975). La definición precisa de los marcos analíticos asociados a la exergía / entropía (límites y contextos de los sistemas), aplicable a ambientes específicos (eventualmente modelable hacia sistemas en espacios y tiempos diferentes) es un útil campo de trabajo.
Un área de investigación de interés está en descubrir los nexos específicos entre la exergía y los diferentes impactos ambientales, en aras de reducir la producción de los mismos (Rosen, 2002). Esto resulta particularmente necesario en ecosistemas tropicales, incluyendo los acuáticos (donde se han logrado grandes avances en zonas templadas) y los ecosistemas terrestres (donde su aplicación es limitada). Retomando a Chen et al. (2009), el análisis exergético tiene una conexión directa con el paradigma de desarrollo sustentable, al posibilitar una medida unificada para medir recursos, flujos, procesos y productividades industriales, así como las transformaciones sistémicas y el estrés ambiental generado por las actividades humanas. Más que una herramienta de uso único para la evaluación de los ecosistemas y de los impactos ambientales, la evaluación exergética representa un potencial de información holística complementaria a las metodologías existentes para la toma de decisiones ambientales, así como para el manejo de las comunidades bióticas y de los ecosistemas.
Agradecimientos
A los revisores anónimos, al profesor Sergio Lopera por su entusiasmo y orientaciones desde GEYMA y de las discusiones desde el curso de capital natural. Al GSM y a sus integrantes por los aportes surgidos en las discusiones académicas. A la Corporación Universitaria Lasallista y a COLCIENCIAS (convocatoria 528/2011) por su apoyo para la formación doctoral de Carlos F. Álvarez-H.
Referencias bibliográficas
Austoni, M.; Giordani, G.; Viaroli, P. & Zaldívar, J. (2007). Application of specific exergy to macrophytes as an integrated index of environmental quality for coastal lagoons. Ecological Indicators, 7(2), 58-62. [ Links ]
Anillo, A. y Barrio, A. (2006). Los principios de la ecología. Análisis de la teoría de ecosistemas de Jørgensen y Fath. En: I Congreso Iberoamericano de Ciencia, Tecnología, Sociedad e Innovación CTS+I. México D.F. 19-23 Junio de 2006. Organización de Estados Iberoamericanos para la Educación la Ciencia y la Cultura (OEI), Agencia Española de Cooperación Internacional (AECI), Universidad Nacional Autónoma de México (UNAM), Instituto Politécnico Nacional (IPN), Universidad Autónoma Metropolitana (UAM), Academia Mexicana de Ciencias y Academia de Ingenieros, 8 p. [ Links ]
Bastianoni, S. & Marchettini, N. (1997). Emergy/exergy ratio as a measure of the level of organization of systems. Ecological Modeling, 99, 33-40. [ Links ]
Bastianoni, S.; Pulselli, F. & Rustici, M. (2006). Exergy versus emergy flow in ecosystems: Is there an order in maximizations?. Ecological Indicators, 6(1), 58-62. [ Links ]
Bligh, D. & Ugursal, V. (2012). Extended exergy analysis of the economy of Nova Scotia, Canada. Energy, 44(1), 878-890. [ Links ]
Chen, G. & Ji, X. (2007). Chemical exergy based evaluation of water quality. Ecological Modeling, 200(1-2), 259-268. [ Links ]
Chen, G.; Jiang, M.; Yang, Z.; Chen, B.; Ji, X. & Zhou, J. (2009). Exergetic assessment for ecological economic system: Chinese agriculture. Ecological Modeling, 220, 397-410. [ Links ]
Christensen, V. (1995). Ecosystem maturity - towards quantification. Ecological Modeling, 77(1), 3-32. [ Links ]
Dewulf, J., Van Langenhove, H.; Muys, B.; Bruers, S.; Bakshi, B.; Grubb, G.; Paulus, D. & Sciubba, E. (2008). Exergy: its potential and limitations in environmental science and technology. Environmental Science Technology, 42(7), 2221-2232. [ Links ]
Doka, G.; Hillier, W.; Kaila, S.; Köllner, T.; Kreiβig, J.; Muys, B.; Garcia-Quijano, J.; Salpakivi-Salomaa, P.; Schweinle, J.; Swan, G. & Wessman, H. (2002). The assessment of environmental impacts caused by land use in the life cycle assessment of forestry and forest products: Guidelines, hints and recommendations. Final report of working group 2 "Land Use" of COST Action E9. BFH Nr. 209, Hamburg, Germany, 92 p.
Fabiano, M.; Vassallo, P.; Vezzulli, L.; Salvo, V. & Marques, J. (2004). Temporal and spatial change of exergy and ascendency in different benthic marine ecosystems. Energy, 29(11), 1697-1712. [ Links ]
Fath, B. & Cabezas, H. (2004). Exergy and Fisher information as ecological indices. Ecological Modeling, 174, 25-35. [ Links ]
Fath, B.; Jørgensen, S.; Patten, B. & Straškraba, M. (2004). Ecosystem growth and development. Biosystems, 77; 213-228.
Fonseca, J.; Marques, J.; Paiva, A.; Freitas, A.; Madeira, V.; Jørgensen, S. (2000). Nuclear DNA in the determination of weighing factors to estimate exergy from organisms biomass. Ecological Modeling, 126, 179-189. [ Links ]
Gaucherel, C. (2006). Influence of spatial patterns on ecological applications of extremal principles. Ecological Modeling, 193(3-4), 531-542. [ Links ]
Gaudreau, K.; Fraser, R. & Murphy, S. (2009).The tenuous use of exergy as a measure of resource value or waste impact. Sustainability, 1, 1444-1463. [ Links ]
Georgescu, N. (1971). The entropy law and the economic problem. USA: Department of Economics, University of Alabama, 7 p. [ Links ]
Georgescu, N. (1975). Energía y mitos económicos. ICE, Revista de Economia, 94-122. [ Links ]
Gibbs, J. (1928). A Method of Geometrical Representation of the thermodynamic properties of substances by means of surface. Trans. Conn. Acad. II, 382-404 or The Collected Works, Yale University Press, 1. [ Links ]
Gong, M. & Wall, G. (2001). On exergy and sustainable development-Part 2: Indicators and methods. Exergy, an International Journal, 1(4), 217-233. [ Links ]
Jenssen, M. (2007). Ecological potentials of biodiversity modelled from information entro-pies: Plant species diversity of north-central european forests as an example. Ecological Informatic, 2(4), 328-336. [ Links ]
Jørgensen, S. (1992). Exergy and ecology. Ecological Modelling, 63(1-4), 185-214. [ Links ]
Jørgensen, S. (2006). Application of holistic thermodynamic indicators. Ecological indicators, 6, 24-29. [ Links ]
Jørgensen, S. (2006-2007). An integrated ecosystem theory. Annals of the European Academy of Science, 19-33. [ Links ]
Jørgensen, S. (2008a). Specific exergy as ecosystem Health Indicator. Sven Erik, Jorgensen y Brian Fath (Eds.) Encyclopedia of Ecology. Elsevier, 3332-3333. [ Links ]
Jørgensen. S. (2008b). Eco-exergy as an ecosystem health indicator. Sven Erik Jorgensen y Brian Fath (Eds.) Encyclopedia of Ecology, Elsevier. 977-979. [ Links ]
Jørgensen, S. (2009). Exergy. Jørgensen, S.E. (Ed.) Ecosystem ecology. Elsevier, Amsterdam. p. 128-141. [ Links ]
Jørgensen, S. & Bendoricchio, G. (2001). Fundamentals of ecological modeling. Elsevier, Amsterdam. 530 p. [ Links ]
Jørgensen, S. & Fath, B. (2004a). Application of thermodynamic principles in ecology. Ecological Complexity. 1(4); 267-280. [ Links ]
Jørgensen, S. & Fath, B. (2004b). Modelling the selective adaptation of Darwin's finches. Ecological Modeling, 176, 409-418. [ Links ]
Jørgensen, S.; Ladegaard, N.; Debeljak, M. & Marques, J. (2005). Calculations of exergy for organisms. Ecological Modeling, 185, 165-175. [ Links ]
Jørgensen, S. & Mejer, H. (1977). Ecological buffer capacity. Ecological Modelling, 3(1), 39-45. [ Links ]
Jørgensen, S. & Nielsen, S. (1998). Thermodynamic orientors: exergy as a goal function in ecological modeling and as an ecological indicator for the description of ecosystem development. In: Müller, F., Leupelt, M. (Eds.), Ecotargets, goal functions, and orientors. Springer-Verlag, Berlin. 63-86 p. [ Links ]
Jørgensen, S.; Nielsen, S. & Mejer, H. (1995). Emergy, environ, exergy and ecological modeling. Ecological Modelling, 77, 99-109. [ Links ]
Jørgensen, S.; Odum, H. & Brown, M. (2004). Emergy and exergy stored in genetic information. Ecological Modeling. 178, p. 11-16. [ Links ]
Jørgensen, S.; Patten, B. & Straškraba, M. (2000). Ecosystems emerging: 4. growth. Ecological Modeling, 126, 249-284.
Jørgensen, S. & Svirezhev, Y. (2004). Towards a Thermodynamic Theory for Ecological Systems. Elsevier, Amsterdam, 380 p. [ Links ]
Koroneos, C.; Nanaki, E. & Xydis, G. (2012). Sustainability Indicators for the Use of Resources-The Exergy Approach. Sustainability, 4(8), 1867-1878. [ Links ]
Libralato, S.;Torricelli, P. & Fabio, P. (2006). Exergy as ecosystem indicator: An application to the recovery process of marine benthic communities. Ecological Modeling, 192, 571-585. [ Links ]
Ludovisi, A.; Pandolfi, P. & Taticchi, I. (2005). The strategy of ecosystem development: specific dissipation as an indicator of ecosystem maturity. J Theor Biol., 235(1), 33-43. [ Links ]
Ludovisi, A. & Poletti, A. (2003). Use of thermodynamic indices as ecological indicators of the development state of lake ecosystems: 2. Exergy and specific exergy indices. Ecological Modeling, 159(2-3), 223-238. [ Links ]
Marques, J. & Jørgensen, S. (2002). Three selected observations interpreted in terms of a thermodynamic hypothesis. Contribution to a general theoretical framwork. Ecological Modeling, 158, 213-222. [ Links ]
Marques, J.; Nielsen, S.; Pardal, M. & Jørgensen, S. (2003). Impact of eutrophication and river management within a framework of ecosystem theories. Ecological Modeling, 166(1-2), 147-168. [ Links ]
Marques, J.; Pardal, M.; Nielsen, S. & Jørgensen, S. (1997). Analysis of the properties of exergy and biodiversity along an estuarine gradient. Ecological Modelling, 102, 155-167. [ Links ]
Ma, Ch.; Zhang, G.; Zhang, X. & Li, Y. (2012). Application of multiple indicators in environment evaluation of coastal restoration engineering: a case study in Bohal Bay in China. Applied Mechanics and Materials, 170-173, 2228-2232. [ Links ]
Martínez, A. (2009). Exergy cost assessment of water resources: physical hidronomics. Ph.D. Dissertation, Department of mechanical engineering, University of Zaragoza Recuperado de: http://zaguan.unizar.es/record/4551/files/TESIS-2010-026.pdf 2009. [ Links ]
Mejer, H. & Jorgensen, S. (1979). Energy and ecological buffer capacity. In: Jorgensen, S.E. (Ed.), State of the art of ecological modelling. Environmental Sciences and Applications, Proc. Conf. on Ecological Modelling, 28th August-2nd September 1978, Copenhagen. International Society for Ecological Modelling, Copenhagen. 829-846. [ Links ]
Molozzi, J.; Salas, F.; Callisto, M.; Marques, J. (2013). Thermodynamic oriented ecological indicators: Application of Eco-Exergy and Specific Eco-Exergy in capturing environmental changes between disturbed and non-disturbed tropical reservoirs. Ecological Indicators, 24, 543-551. [ Links ]
Müller, F. (2005). Indicating ecosystem and landscape organisation. Ecological Indicators, 5, 280-294. [ Links ]
Munari, C. & Mistri, M. (2006). Application of the exergy method to benthic assemblages of the Valli di Comacchio (Italy): Environmental gradient assessing. Italian Journal of Zoology, 73(3), 237-245. [ Links ]
Patricio, J.; Marcos, C.; Pardal, M.; Pérez, J.; Marques, J. & Salas, F. (2006b). Are taxonomic distinctness measures compliant to other ecological indicators in assessing ecological status?. Marine Pollution Bulletin, 52, 817-829. [ Links ]
Patricio, J.; Neto, J.; Teixeira, H.; Salas, F. & Marques, J. (2009). The robustness of ecological indicators to detect long-term changes in the macrobenthos of estuarine systems. Marine Environmental Research, 68(1), 25-36. [ Links ]
Patricio, J.; Salas, F.; Pardal, M.; Jørgensen, S. & Marques, J. (2006a). Ecological indicators performance during a re-colonisation field experiment and its compliance with ecosystem theories. Ecological Indicators, 6, 43-57. [ Links ]
Park, Y.; Lek, S.; Scardi, M.; Verdonschot, P. & Jørgensen, S. (2006). Patterning exergy of benthic macroinvertebrate communities using self-organizing maps. Ecological Modeling, 195 (1-2): 105-113. [ Links ]
Pusceddu, A. & Danovaro, R. (2009). Exergy, ecosystem functioning and efficiency in a coastal lagoon: The role of auxiliary energy. Estuarine, Coastal and Shelf Science, 84(2), 227-236. [ Links ]
Ruth, M. (1995). Information, order and knowledge in economic and ecological systems: implications for material and energy use. Ecological Economics, 13, 99-114. [ Links ]
Rosen, M. (2002). Can exergy help us understand and address environmental concerns. Exergy, an International Journal, 2, 214-217. [ Links ]
Salas, F.; Marcos, C.; Pérez-Ruzafa, A. & Marques, J. (2005). Application of the exergy index as ecological indicator of organically enrichment areas in the Mar Menor lagoon (southeastern Spain). Energy, 30, 2505-2522 [ Links ]
Sciubba, E. (2009). Exergy-based ecological indicators: a necessary tool for resource use assessment studies. Termotehnica, 2, 11-25. [ Links ]
Sciubba E.; Bastianoni S y Tiezzi, E. (2008). Exergy and extended exergy accounting of very large complex systems with an application to the province of Siena, Italy. J. Environ Manage, 86(2): 372-82. [ Links ]
Sciubba, E. & Ulgiati, S. (2005). Emergy and exergy analyses: Complementary methods or irreducible ideological options?. Energy, 30, 1953-1988. [ Links ]
Sciubba, E. & Wall, G. (2007). A brief commented history of exergy from the beginnings to 2004. Int. J. of Thermodynamics, 10(1), 1-26. [ Links ]
Sciubba, E. & Zullo, F. (2012). Stability and limit cycles in an exergy-based model population dynamics. Proceedings of ECOS 2012, 25 International Conference on Efficiency, Cost, Optimization, Simulation and Environmental Impact of Energy Systems. Perugia, Italy, 14 p. [ Links ]
Schneider, E. & Kay, J. (1994). Life as a manifestation of the second law of thermodynamics. Mathematical and Computer Modeling, 19 (6-8), 25-48. [ Links ]
Schrödinger, E. (1944). What is life?. Cambridge University Press. 184 p. [ Links ]
Seckin, C. & Bayulken, A. (2012). Determination of environmental remediation cost of municipal waste in terms of extended exergy. Proceedings of ECOS 2012, 25 International Conference on Efficiency, Cost, Optimization, Simulation and Environmental Impact of Energy Systems. Perugia, Italy, 14 p. [ Links ]
Silow, E.& In-Hye, O. (2004). Aquatic ecosystem assessment using exergy. Ecological Indicators, 4(3), 189-198. [ Links ]
Silow, E. & Mokry, A. (2010). Exergy as a tool for ecosystem health assessment. Entropy, 12, 902-925. [ Links ]
Szargut, J.; Morris, D. & Steward, F. (1988). Exergy analysis of thermal, chemical, and metallurgical processes. Hemisphere, 332 p. [ Links ]
Tundisi, J.; Tundisi, M. & Periotto, N. (2012). Exergy and ecological services in reservoirs. Braz. J. Biol, 72(4), 979. [ Links ]
Wagendorp, T.; Gulinck, H.; Coppin, P. & Muys, B. (2006). Land use impact evaluation in life cycle assessment based on ecosystem thermodynamics. Energy, 31(1), 112-125. [ Links ]
Wall, G. (1977). Exergy a useful concept within resource accounting, Report No. 77-42, Göteborg, Sweden: Institute of Theoretical Physics, 61 p. [ Links ]
Wall, G. (1993). Exergy, ecology and democracy - concepts of a vital society or a proposal for an exergy tax. ENSEC'93. International Conference on Energy Systems and Ecology, July 5-9, Krakow, Poland. Publ. Proc., Szargut, J., et al., eds., p. 111-121. [ Links ]
Wall, G. & Gong, M. (2001). On exergy and sustainable development-Part 1: Conditions and concepts. Exergy, an International Journal, 1 (3), 128-145. [ Links ]
Xu F. (1997). Exergy and structural exergy as ecological indicators for the development state of the Lake Chaohu ecosystem. Ecological Modelling, 99(1), 41-49. [ Links ]
Xu, F.; Dawson, R.; Tao, S.; Li, B. & Cao, J. (2002). System-level responses of lake ecosystems to chemical stresses using exergy and structural exergy as ecological indicators. Chemosphere, 46(2), 173-185. [ Links ]
Xu, F.; Jørgensen, S. & Tao, S. (1999). Ecological indicators for assessing freshwater ecosystem health. Ecological Modeling, 116: 77-106. [ Links ]
Xu, F.; Wang, J.; Chen, B.; Qin, N.; Wu, W.; He, W.; He, Q. & Wang, Y. (2011). The variations of exergies and structural exergies along eutrophication gradients in Chinese and Italian lakes. Ecological Modelling, 222(2), 337-350. [ Links ]
Xu, Y.; Zhang, H.; Huang, T. & Su, Y. (2012). A study on relationship between biomass of macrophytes and the health status of the lake ecosystem. Applied Mechanics and Materials, 209-211, 166-1169. [ Links ]
Zaleta, A.; Ranz, L. & Valero, A. (1998). Towards a unified measure of renewable resources availability: the exergy method applied to the water of a river. Energy Conversion and Management, 39(16-18), 1911-1917. [ Links ]
Zhang, J.; Gurkan, Z. & Jørgensen, S. (2010). Application of eco-exergy for assessment of ecosystem health and development of structurally dynamic models. Ecollogical Modeling, 221: 693-702. [ Links ]
Zhang, B.; Peng, B. & Liu, M. (2012). Exergetic assessment for resources input and environmental emissions by Chinese industry during 1997-2006. The Scientific World Journal, 13 p. [ Links ]
