










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInternational Journal of Psychological Research
Print version ISSN 2011-2084
int.j.psychol.res. vol.6 no.spe Medellín Oct. 2013
Probing Prejudice with Startle Eyeblink Modification: A Marker of Attention, Emotion, or Both?
Examinando el prejuicio mediante la modificación del parpadeo por sobresalto: ¿Un indicador de atención, emoción, o ambos?
Eric J. Vanmana,*, John P. Ryanb, William C. Pedersenc, Tiffany A. Itod,
a University of Queensland, School of Psychology, Brisbane, Australia.
b University of Pittsburgh, Department of Psychiatry, Pittsburgh, United States.
c California State University, Long Beach, Department of Psychology, Long Beach, United States.
d University of Colorado at Boulder Department of Psychology and Neuroscience,Boulder, United States.
* Corresponding author. Eric Vanman, School of Psychology, McElwain Building, Level 3, University of Queensland, St Lucia, QLD 4072, Australia. Email: e.vanman@uq.edu.au
Received: 24-07-2013-Revised: 26-09-2013-Accepted: 08-10-2013
ABSTRACT
In social neuroscience research, startle eyeblink modification can serve as a marker of emotion, but it is less clear whether it can also serve as a marker of prejudice. In Experiment 1, 30 White students viewed photographs of White and Black targets while the startle eyeblink reflex and facial EMG from the brow and cheek regions were recorded. Prejudice was related to facial EMG activity, but not to startle modification, which instead appeared to index attention to race. To test further whether racial categorizations are associated with differential attention, a dual-task paradigm was used in Experiment 2. Fifty-four White and fifty-five Black participants responded more slowly to a tone presented when viewing a racial outgroup member or a negative stimulus, indicating that both draw more attention than ingroup members or positive stimuli. We conclude that startle modification is useful to index differential attention to groups when intergroup threat is low.
Palabras Clave: Startle, Facial electromyography, Affect, Prejudice, Attention.
RESUMEN
En la investigación de la neurociencia social, la modificación del parpadeo de sobresalto puede servir como un marcador de emoción, pero no es tan claro si también puede servir como un marcador de prejuicios. En el experimento 1, 30 estudiantes blancos vieron fotografías de objetivos a blanco y Negro mientras se registraba el reflejo del parpadeo de sobresalto y las EMG faciales de las regiones de las cejas y las mejillas. El prejuicio se relacionó con la actividad EMG facial, pero no con la modificación de sobresalto, en cuyo lugar apareció la atención al índice de raza. A fin de probar si las categorizaciones raciales están asociadas con la atención diferencial, se utilizó un paradigma de doble tarea en el Experimento 2. 54 participantes blancos y 55 participantes negros respondieron más lento a un tono presentado al visualizar un miembro del grupo racial externo o de un estímulo negativo, lo que indica que ambos atraían más la atención que los miembros del mismo grupo o estímulos positivos. Llegamos a la conclusión de que la modificación de sobresalto es útil para indexar la atención diferencial a los grupos cuando la amenaza entre éstos es baja.
Key Words: Sobresalto, Electromiografía facial, Sentimientos, Prejuicios, Atención.
1. INTRODUCTION
Social categorization, especially for some well-developed categories (e.g., race, gender, age), is relatively effortless, unconscious, and occurs without intention (Brewer, 1988; Van Bavel & Cunningham, 2011). Typically, the resulting categorization automatically leads to ingroup bias, in which more positive evaluations are made toward ingroup members and more negative affect is directed toward outgroup members (Kubota, Banaji, & Phelps, 2012; Molenberghs, 2013; Mullen, Brown, & Smith, 1992). Social categorizations are advantageous because they enable us to identify others who may provide benefits to the self (i.e., ingroup members) while also helping us to recognize others who may pose a threat (i.e., outgroup members). People are therefore also likely to attend to ingroup and outgroup members differently. Humans, like other animals, are particularly prone to attend more to stimuli that are threatening than other kinds of stimuli (Öhman, 1997). This attentional bias may reflect an evolved module of fear and fear learning (Öhman & Mineka, 2001). Prior research suggests that we have a similar preparedness to associate outgroup members with danger, as they may be likely associated with a threat in the past (Mallan, Sax, & Lipp, 2009; Öhman, 2005). Therefore, outgroup members should also draw greater attention than do ingroup members.
The aim of the present research was to examine the extent that startle eyeblink modification can differentially index the emotional and attentional biases that accompany social categorizations. In a typical investigation of emotional modification of the startle reflex, a loud, startling burst of white noise is presented at some lead interval after picture onset while the research participant views a picture from the International Affective Picture System (IAPS; e.g., Bradley, Cuthbert, & Lang, 1993; Mallan & Lipp, 2007; Price, Dieckman, & Harmon-Jones, 2012; Vanman, Boehmelt, Dawson, & Schell, 1996). At longer lead intervals (i.e., 800 ms or longer), the presentation of negative pictures leads to augmentation of the startle response compared to instances when the participant views neutral and positive ones. However, the reflex can also be augmented or attenuated as a function of task-relevant or salient features of the stimulus, irrespective of arousal (Filion, Dawson, & Schell, 1993; Lipp, Siddle, & Dall, 1997; Simons & Zelson, 1985). Startle eyeblinks are generally attenuated for foreground stimuli that the participant finds especially salient or interesting, but are augmented for stimuli towards which the participant is deliberately directing his or her attention (see Filion, Dawson, & Schell, 1998, for a comprehensive review).
As a comparison, in this research we also used facial EMG to measure affect elicited by social categorizations. EMG recorded from the cheek and brow regions, corresponding to activity of the zygomaticus major (the muscle in the cheek that pulls up the lip corner) and corrugator supercilii (the muscle above the eye that pushes the brows together), respectively, can reliably index changes in positive and negative affect (Tassinary, Cacioppo, & Vanman, 2007). What is especially relevant here is that, facial EMG is a reliable marker of affective responses elicited by the presentation of group memberships of target individuals (e.g., Vanman, Paul, Ito, & Miller, 1997; Vanman, Saltz, Nathan, & Warren, 2004). In studies where both startle eyeblink modification and facial EMG have been employed, affective modification of the startle reflex was related to the amount of facial EMG activity in the respective cheek and brow regions (Bradley, Cuthbert, & Lang, 1996; Witvliet & Vrana, 1995).
1.1. Can Racial Biases Modify the Startle Reflex?
On the basis of prior research, it is plausible that startle modification might serve as a marker of prejudice when individuals view pictures of ingroup and outgroup members. That is, viewing pictures of outgroup members for which an individual holds antipathy should be accompanied with facilitation of the startle response, in comparison to viewing pictures of ingroup members (see Guglielmi, 1999, who made a similar hypothesis). This hypothesis, however, has garnered mixed support in studies of racial prejudice. For example, in one study (Amodio, Harmon-Jones, & Devine, 2003) White participants, who had completed measures of prejudice towards Blacks, viewed White, Black, and Asian photos in a startle modification paradigm. At the early lead interval (400 ms), the most prejudiced participants evidenced more inhibited startle to Black faces than they did to White faces, whereas at the late lead interval (4000 ms) the more prejudiced subgroups evidenced more facilitated startle to Black faces than to White faces. Interestingly, there were no ingroup-outgroup differences when participants viewed photos of Asians. Similarly, Phelps et al. (2000) included a startle modification measure in a study in which Whites also viewed pictures of Blacks and Whites while in an fMRI scanner. Although there were no significant startle modification effects when probes were presented, startle modification differences were correlated with differences in the activity of the amygdala. Finally, Brown, Bradley, and Lang (2006) presented White and Black participants photos of Whites and Blacks in either positive or negative contexts. Although startle modification was affected by the emotional valence of the photos, the race of the subjects in the photos had no effect on startle. This finding suggests that, in intergroup situations where contact with the outgroup is not characterized by threat or other strong emotions, stimuli representing the outgroup should be less likely to elicit effects of affective modification of startle, and be more sensitive to the attentional qualities of the stimulus. One might speculate that those with greater differences in amygdala activity in Phelps et al. (2000), or the high prejudiced groups in Amodio et al. (2003), were sufficiently aroused by the Black stimuli (but not by the Asian stimuli) to exhibit an emotional effect on startle, whereas participants were not sufficiently aroused by the racial stimuli in Brown et al. (2006). Note that none of these studies was designed to examine attentional effects on startle modification.
In this paper we report two studies that examined the affective and attentional consequences of social categorizations. In the first study, we hypothesized that the presentation of pictures of outgroup members, particularly for a social category against which an individual holds prejudice, should elicit facial EMG activity consistent with an affective bias against the outgroup. With respect to startle modification, however, we predicted that viewing an outgroup photo in a non-threatening context should elicit a pattern of startle consistent with other findings of attentional effects (i.e., attenuated startle for “interesting” stimuli at long lead intervals), but not consistent with affective modification of startle (i.e., enhanced startle for negative stimuli at long lead intervals). To test this hypothesis, we used a variation of the affect-directed attention paradigm (Vanman et al., 1996). The affect-directed attention paradigm, like other startle probe paradigms, allows one to track the time course of the processing of affect-laden pictures at both early and late stages of perceptual processing, but it may be more sensitive to any emotional processing of the stimulus that occurs just after its onset. The results from Experiment 1 led us to conduct a follow-up study designed to explore further the effects of racial categorization.
2. EXPERIMENT 1
2.1. OVERVIEW
A group of White university students viewed a set of pictures of White and Black students twice. During the first presentation of the set, the participants rated each target individual on apparent friendliness. During the second viewing of the picture set, a startle probe was presented on some trials at early and late lead intervals following picture onset. During trials in which no startle probe was presented, facial EMG from the cheek and brow regions was recorded. In addition, participants also completed a politically-based measure of prejudice against African Americans as a group.
Because the stimuli were pictures of Blacks and Whites, we predicted that participants would evidence intergroup bias in favor of their ingroup. However, we predicted that this ingroup favoritism would not be evident in the self-report ratings. In contemporary American society, such self-reports of evaluations based on race are typically susceptible to social desirability concerns, and thus may not serve as veridical indicators of a person's attitude toward members of a particular group. This dissembling of self-reports and physiological responses is an important difference between what is found when participants view pictures from the IAPS and when they view evaluate social stimuli based on race.
Finally, by using the affect-directed attention paradigm, we examined whether giving instructions to participants to attend to either the Black or White pictures would yield directed attention effects on startle modification--effects similar to those found in other studies that have included such instructions with non-emotional stimuli.
2.2. METHOD
Participants. Thirty White students (18 women, 12 men) enrolled in introductory psychology courses at a university in metropolitan Los Angeles participated for extra credit.
Materials. Stimuli consisted of 32 photos (8 Black males, 8 Black females, 8 White males, 8 White females) selected from a larger sample of photos that had been previously rated for attractiveness and familiarity by 96 students from an introductory psychology class in a previous semester. The photos were taken on two university campuses four years before, and showed students smiling slightly in a variety of settings, none of which identified the particular campus. The stimuli used in this study were selected because their attractiveness ratings were similar across the different racial and gender groupings.
Procedures. When the participants arrived at the laboratory, the experimenter told them that the study was concerned with physiological processes involved in person perception and ethnicity. They then completed a brief health questionnaire and a “student opinion survey,” which consisted of 40 items about student issues and social attitudes. To measure participants' general racial attitudes about African Americans, seven items were embedded in this questionnaire that comprised the Modern Racism Scale (MRS; McConahay, 1986) A 5-point rating scale (-2=strongly disagree, +2=strongly agree) was used. According to the MRS, the sample had moderately low levels of prejudice towards African Americans, M = -8.70, SD = 0.69 (on a -14 to +14 overall scale).
Electrodes (Ag/AgCl, 4 mm in diameter) were then attached to record surface EMG activity from the corrugator supercilii and zygomaticus major following previous recommendations regarding these sites (Fridlund & Cacioppo, 1986). To record the startle eyeblink as EMG activity, a pair of electrodes was attached directly over the region of the right orbicularis oculi inferior. A ground electrode was attached to the right earlobe.
Participants sat in a comfortable reclining chair wearing headphones for the remainder of the experiment. After the electrodes were attached, a 5-min resting period occurred, at the end of which three presentations of the startling white noise burst were presented as examples. The participants then viewed the entire set of 32 photos once without any physiological responses recorded. During this first viewing, each photo was presented for 3 s, followed by a 10-s intertrial interval. Participants rated each photo for how friendly the target appeared to be using a 7-point rating scale (1 = not at all friendly, 7 = very friendly). The photos were projected onto a white screen mounted approximately 5 m from the participant's chair.
When the first viewing was completed, the experimenter then told the participant that the photos would be presented again but this time they would differ in duration and order. The instructions indicated that most photos would be 5 s in duration, but some would be 7 s in duration. The task for the participant was to attend to the duration of the pictures of White (or Black) students, and to ignore the duration of the Black (or White) students (with the assignment of attend and ignore counterbalanced across participants). At 10 s following the offset of each to-be-attended photo, the participant was prompted on the computer monitor to indicate whether it was longer than usual by pressing one of two keys on the keypad. Twenty-four (12 of each target race) of the photos were displayed for 5 s, and eight (four of each target race) were displayed for 7 s. Intertrial intervals varied between 25 and 35 s. Customized software was used for the presentation and control of the data acquisition.
All physiological recordings were made during the second photo presentation. The 32 trials were organized into four blocks of eight trials. Four of these eight trials consisted of photos of White students (two males, two females), and the other four consisted of Black students (two males, two females). The startle probe (i.e., a 103-dB (A) SPL white noise burst, 40 ms in duration) was presented on three of the four trials that comprised each target race, whereas the remaining one was a “clear trial,” which was when zygomaticus and corrugator EMG activity was recorded. Each of the three startle probe trials contained a probe at either 300, 800, or 4500 ms following photo onset. The order of the events across the eight trials within each block was randomly determined and counterbalanced across participants, with the restriction that the first trial of the first block was always a clear trial.
In addition to the probes presented during the photo presentations, startle stimuli were also presented during 24 of the intertrial intervals, six occurring during each trial block. These probes were presented at random intervals between 10 and 20 s into the intertrial interval. Responses to these probes served as baseline measures with which to compare blink amplitudes elicited during the pictures. Upon completion of the last trial, participants were fully debriefed and dismissed.
Data acquisition and reduction. EMG signals were relayed through a shielded cable to one of three Grass 7P3 preamplifer/integrators, each with a bandpass of 10 Hz to 5 kHz. Signals were full wave rectified and smoothed using a contour-following integrator with a time constant of 0.05 s for zygomaticus and corrugator EMG and a time constant of 0.02 s for orbicularis oculi EMG. On clear trials, EMG activity from the zygomaticus and corrugator sites was digitized at 100 Hz for 5 s following photo onset. On probe trials, EMG activity from the orbicularisoculi site was digitized at 1000 Hz for 300 ms following the presentation of each startle probe.
During the experimental session the rectified and smoothed EMG recording were continuously displayed on a polygraph, and participants were monitored using a videocamera housed unobtrusively in a “message box” slightly above and directly in front of the participant. For the zygomaticus and corrugator data, mean amplitude of EMG activity was computed for all clear trials, and these mean amplitudes were averaged across trials within a condition and within participants to obtain more reliable and normally distributed estimates of treatment effects.
For the startle probe trials, the amplitude of each eyeblink elicited during a photo presentation was expressed as a percent change score from the mean of the amplitudes of the blinks elicited during the intertrial intervals in each block. A positive modification score indicates that the blink during a photo was larger than the average blink elicited during the intertrial intervals, whereas a negative blink modification score indicates that the blink during a photo was smaller than the intertrial blinks. Trials in which no blink occurred in response to the probe were scored as zero. In addition, blinks were discarded if the data were too noisy to be scored by the computer or the experimenter noted excessive movement at the time the startle probe was presented. Unscorable blinks occurred on less than 3% of the startle probe recording epochs.
2.3. RESULTS
For the analyses reported, repeated measures analyses of variance were performed using the multivariate approach, whenever appropriate. An alpha level of .05 was used for all statistical tests. Effect sizes are reported using η2 and Cohen's d. Due to equipment failure and experiment error, the zygomaticus data for one participant and the corrugator data for four participants were not collected, but the remaining data for those participants were included in other analyses.
Friendliness Ratings. Analyses of friendliness ratings revealed no main effects of target race or prejudice group, but there was a Target Race X Prejudice Group interaction, F(1,27) = 6.37, p = .018, η2 =.19. Post hoc comparisons revealed that the low prejudice participants evidenced no bias in their friendliness ratings for White targets (M = 4.87, SE = 0.10) and Black targets (M = 5.00, SE = 0.10), t(15) = 1.64, p = .122, d = 0.85, but high prejudice participants tended to show a bias favoring Whites targets (M = 4.94, SE = 0.16) over Black targets (M = 4.77, SE = 0.19), t(12) = -1.96, p = .074, d = -1.13.
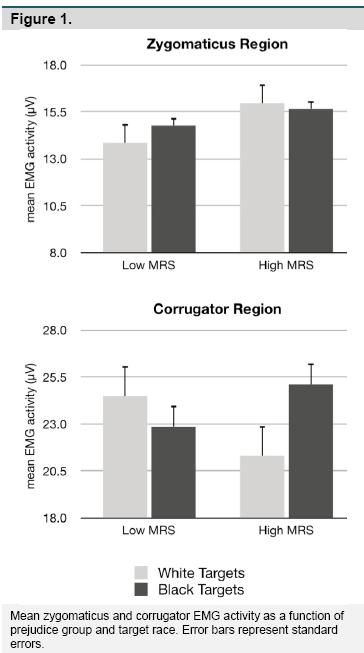
Facial EMG. Figure 1 presents the means and standard errors for the zygomaticus and corrugator EMG activity as a function of prejudice group and target race. For corrugator EMG, the analysis revealed no main effects of target race or prejudice group, but there was a Target Race X Prejudice Group interaction, F(1,24) = 5.50, p = .027, η2 η2 = .18. Post hoc comparisons revealed that the high-prejudiced participants tended to display more corrugator activity to Black targets, t(11) = 1.94, p = .08, d = 1.17, but low-prejudiced participants did not, t(13) = -1.24, p = .24, d = -0.69. For zygomaticus EMG, there were again no main effects of target race or prejudice group, but the analysis did reveal a Target Race X Prejudice Group interaction, F(1,27) = 4.70, p =.039, = .14. That is, low-prejudice participants displayed more zygomaticus activity to Black targets, t(15) = 2.29, p =.037, d = 1.18, but high-prejudiced participants did not, t(12) = -0.79, p =.443, d = -0.46. No other effects were found.
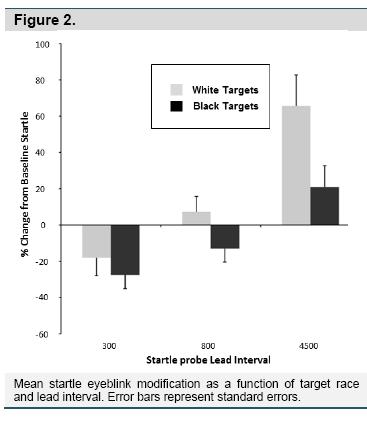
Startle Blink Modification. The startle analysis was first conducted as a 2 (Target Race) X 2 (Prejudice Group) X 3 (Lead Interval) mixed ANOVA, but this analysis revealed no main effects or interactions with prejudice group, so the remaining analyses collapsed across the prejudice group variable. Figure 2 depicts the resulting 2 (Target Race) X 3 (Lead Interval) interaction, F(2,50) = 3.28, p = .048, η2 =.11. A series of post hoctests revealed no differences between White and Black targets at 300 ms, but target race differed at 800 ms, t(26) = 2.23, p = .035, d = 0.88, and at 4500 ms, t(28) = 2.40, p =.023, d = 0.91. That is, for the 800 and 4500 lead intervals only, blinks elicited during the viewing of White targets were larger than those elicited during the presentation of Black targets. Ancillary analyses also examined participant sex and attention instructions (i.e., attend to White, ignore Black vs. attend to Black, ignore White), but they yielded no main effects or interactions associated with the target race manipulation in any analysis.
Other Relationships Between Measures. To explore the relationships between the Modern Racism Scale scores, friendliness ratings, startle eyeblink modification, and facial EMG, we computed bias scores (mean of Black targets- mean of White targets) for friendliness, zygomaticus and corrugator EMG activity, and startle modification at each lead interval. Of the correlations then tested between these variables, only a relationship between Modern Racism Scale scores and bias in corrugator EMG activity, r(23) = .481, p = .015, and between Modern Racism Scale scores and bias zygomaticus EMG activity, r(27) = -.415, p = .025, were found. Higher scores on the Modern Racism Scales were associated with more corrugator activity in favor of Black targets and more zygomaticus activity in favor of White targets. Indeed, bias in brow activity was negatively correlated with bias in zygomaticus activity, r(23) = -.402, p = .046. No association between Modern Racism Scale scores and biases in startle eyeblink modification was found at any lead interval.
2.4. DISCUSSION
The pattern of results for facial EMG measures in this study provides evidence that high-prejudiced participants found pictures of Blacks more unpleasant than pictures of Whites, and thus these particular results are consistent with previous research that has used facial EMG to measure racial bias (Brown et al., 2006; Vanman et al., 2004). In contrast, startle eyeblink modification appeared to index differential attention to ingroup and outgroup members. Blinks were smaller at the 800 and 4500 ms probes when participants viewed pictures of Blacks compared to pictures of Whites. As mentioned, attentional effects of startle modification have been demonstrated in several studies (see Filion et al., 1998), and these effects have differed, depending on whether attention was directed explicitly by instructions, or implicitly by intrinsic aspects of the stimuli. Experiments that have shown these instructional effects on attention have usually used fairly simple stimuli, such as colored light or a tone. In our experiment, the explicit attentional task (e.g., attend to the duration of Whites, ignore the duration of Blacks) had no direct effects on startle (see also Vanman et al., 1996, who found a similar lack of effect for task instructions with more interesting stimuli), perhaps because categorizing faces of ingroup and outgroup members on the basis of race is automatic and not interrupted by the demands of the task instructions. If startle modification is partially determined by the intrinsic interest of a stimulus, then one can infer that pictures of Black students in this study attracted more attention than did the pictures of White students.
Importantly, there was no evidence of affective modification of startle (i.e., the outgroup enhancing startle compared to the ingroup) at any lead interval, which is consistent with other research that used more emotional stimuli in an interracial context (Brown et al., 2006). Although Amodio et al. (2003) found, for at least a subset of their participants, larger blinks when White participants viewed the Black outgroup targets, they did not find this pattern for the Asian outgroup targets. Amodio et al. (2003) interpreted their late lead interval effects for the Black stimuli as consistent with an affective account of startle (i.e., the outgroup is associated with threat), but also concluded that the lack of effects for the Asian stimuli were likely due to the absence of threatening attributes in Americans' stereotypes of Asians. Because our results do not support the affective pattern of startle modification, we infer that participants were not sufficiently threatened by the stimuli in this context, but still found the outgroups more interesting because of novelty and/or a general negativity associated with outgroups. In other situations where participants have less contact with the outgroup (as perhaps was the case in the Amodio et al. study), startle modification may be more affected by the emotional intensity of the stimuli.
The findings of the Amodio et al. (2003) study raise the question about whether startle modification effects will be found for only those outgroups that have threatening stereotypes associated with them. That is, even if emotional arousal is not sufficiently evoked by the stimuli, the presentation of members of an outgroup associated with violence, for example, may still require more attentional resources. Indeed, all the participants in our study were White, and they may have associated threatening cultural stereotypes with Blacks in general, even though they held moderate attitudes towards African Americans. Other research has demonstrated that both low- and high-prejudiced students in the United States automatically activate a violence stereotype of African Americans when presented with instances of the group or a symbolic equivalent (Devine, 1989). Thus, the attenuation of the startle reflex that occurred for the 800 and 4500 ms probes during the presentation of Black targets in comparison to presentations of White targets may have indexed differential attention by the participants because of the automatic activation of negative stereotypes associated with Blacks. If it is a threatbased stereotype of African Americans in particular that draws more attention in White participants (Donders, Correll, & Wittenbrink, 2008; Trawalter, Todd, Baird, & Richeson, 2008), then African Americans should also attend more to photos of Blacks than Whites, as both Whites and Blacks appear to share the association of a violent, threatening stereotype more readily with African Americans than with Whites.
To examine this interpretation of the startle modification findings of Experiment 1, we conducted a second study that demonstrated convergent validity by using a behavioral measure sensitive to attentional manipulations. Dual-task paradigms allow one to assess the degree to which the attentional demands of two different tasks interfere with one another (Luck & Vecera, 2002). In particular, a secondary reaction-time paradigm requires that the participant attend to a primary task (e.g., categorizing red and blue squares) while a reaction time probe (e.g., a tone) is presented (Posner, 1978). If the primary task momentarily taxes limited attentional resources, the latency to the reaction time probe is longer. Moreover, the reaction time probe can be presented at different lead intervals following the onset of the primary task to track the time course of task processing. In Experiment 2, the primary task was to categorize the race of a target while responding to any tones (presented at early and late lead intervals) as quickly as possible. In addition, because the second study was conducted at a different university in a different city, we were able to recruit two different racial/ethnic samples (i.e., White and African American) to view photos of White and Black faces. We hypothesized that both Whites and African Americans would attend more (i.e., have longer response latencies to the probe) to their respective outgroup than to the ingroup. Alternatively, if the Black stimuli drew more attention because of threatening stereotypes of African Americans, then one would predict that only the White participants would attend differentially to the stimuli. As a comparison, we also presented pleasant and unpleasant IAPS photos. Consistent with our reasoning that negative information is more salient, we predicted that all participants would show evidence of greater attention to negative pictures.
3. EXPERIMENT 2
3.1. METHOD
Participants. One hundred nine (54 Black and/or African American, 55 White) university students, with nearly identical numbers of men and women within each ethnic group, enrolled in introductory psychology courses at a university in metropolitan Atlanta participated for extra credit.
Materials. Two sets of photos comprised the stimuli in this study. The first set consisted of 16 photos of senior high school students selected from yearbooks--eight White males and eight Black males. These stimuli were used by Vanman et al. (2004). The second set consisted of 16 photos selected from the IAPS (Lang, Bradley, & Cuthbert, 2005) that did not include people. Normative ratings that accompany the set were used to select eight rated as pleasant (e.g., ice cream sundae, beautiful scenery, cute animals) and eight rated as unpleasant (garbage, cockroach, a vicious dog), with each subset of eight being rated equivalent in arousal. All photos were presented on a computer monitor positioned approximately 0.75 m from the participant.
Participants also completed standard questionnaires regarding their racial attitudes. To measure racial attitudes about their respective outgroups, we used attitude measures that were originally standardized for White and African American participants, respectively. Thus, White participants completed the Symbolic Racism 2000 Scale (Henry & Sears, 2002), whereas Black participants completed the Johnson and Lecci (2003) Scale.
Procedure. Photos were presented to participants one at a time for 5 s. Each photograph was viewed four times (non-sequentially) over the course of the experiment. During one presentation of each photo, an auditory tone was presented 300 ms after picture onset. In a separate trial, an auditory tone was presented 1500 ms after picture onset. In other trials, no tone was presented. Based on random assignment, participants were instructed either to attend to the duration of the pictures of White students or the duration of the pictures of Black students. Stimuli were divided into two runs. At the end of each run, participants were asked to report the number of photos of their assigned racial category (Black or White) that had been shown for 7 s. In fact, all were shown for 5 s.
3.2. RESULTS
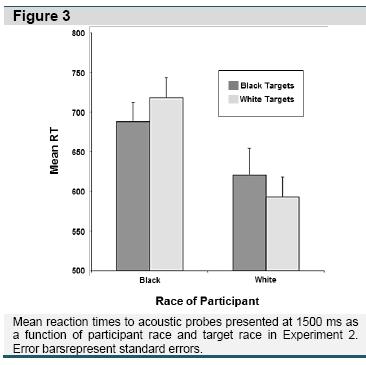
Intergroup Stimuli. A 2 (Participant Race) X 2 (Target Race) ANOVA revealed no effects at the 300 ms lead interval. However, analyses revealed a main effect of participant race, F (1,107) = 7.08, p = .009, η2 = .05, at the 1500 ms lead interval, which was qualified by a two-way Participant Race X Target Race interaction, F(1,107) = 4.42, p = .038, η2 = .04. As depicted in Figure 3, White participants had longer latencies to the acoustic probe when viewing Black targets than White targets t(54) = 2.619, p = .011, d = 0.71, whereas Black participants had longer latencies to the probe when viewing White targets than Black targets t(53) = -2.181, p = .034, d = -0.60.
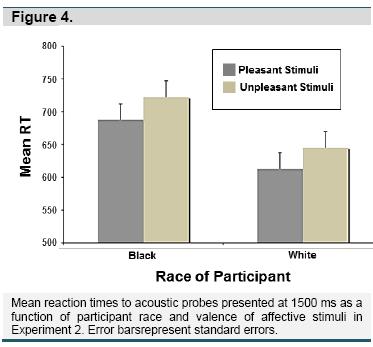
Affective Stimuli. Similar analyses were conducted for the IAPS stimuli. For each probe lead interval, a 2 (Participant Race) X 2 (Picture Valence) mixed factorial ANOVA was conducted on the mean latency for each condition. Again, no effects were observed at the 300-ms lead interval, but there was a main effect of Picture Valence at 1500 ms, F(1,107) = 12.80, p = .001, η2 = .11, with longer latencies to unpleasant photos (M = 678.50, SE = 28.15) than to pleasant photos (M = 649.72, SE = 26.44), regardless of the participant's race. Figure 4 depicts the means for each group for the 1500 lead interval.
Additional Analyses.To examine the effect of self-reports of prejudice on secondary reaction time to the intergroup stimuli, the latency data were regressed on scale scores for each measure separately (i.e., Johnson-Lecci, Symbolic Racism 2000). These analyses also revealed no effects.
3.3. DISCUSSION
The results from this experiment demonstrated that people do attend more to outgroup members than they do to ingroup members, as indicated by the longer latencies to the secondary reaction time probe when participants were viewing a picture of an outgroup member. This finding is consistent with the attentional interpretation of the startle eyeblink modification results of Experiment 1. This intergroup attentional effect, however, does not appear to be limited to groups with which cultural stereotypes of threat are associated, as both Black and White participants attended more to their respective outgroup. Moreover, none of these effects were moderated by attitudes about outgroup members as a whole. The two groups of participants differed in their mean reaction times, but the present research unfortunately does not readily provide an explanation for this group difference. It is possible, for example, that African American participants attended more than White participants to all classes of stimuli in this experiment because they were more anxious or vigilant due to the race-based context of the experiment.
Importantly, both Black and White participants attended more to negative than to positive IAPS stimuli. That is, when they viewed a picture of an animal carcass, for example, their reaction time to the acoustic probe was longer than when they viewed a picture of puppies. This result is consistent with other studies that have found that negatively valenced stimuli draw more attention than do positively valenced ones (Fiske, 1980; Ito & Cacioppo, 2000; Öhman & Mineka, 2001). Therefore, it is plausible to infer that the greater attention one exhibits towards an outgroup member is partly based on some sort of intergroup antipathy. In many instances outgroup members are not typically associated with full-on emotions such as fear. For example, there are occasions where intergroup relations are characterized by peaceful co-existence, or at least a tolerance of the others' presence. We might expect individuals to attend more to outgroup members in these situations perhaps because of the relative proportions of ingroup and outgroup members in the setting. For example, if one rarely encounters members of the outgroup in daily life, contact with an outgroup member should be relatively novel, and thus draw more attention (Oakes & Turner, 1986). Another possibility lies in research indicating that negative social information draws more attention than positive information (e.g., Carretié, Mercado, Tapia, & Hinojosa, 2001; Ito & Cacioppo, 2000). Indeed, the results of this study suggest that the antipathy associated with an outgroup leads to greater attention to outgroup than to ingroup members.
4. GENERAL DISCUSSION
This research examined whether startle eyeblink modification can serve as a marker of prejudice. Our findings suggest that among individuals whose prejudice towards a racial outgroup is low, startle modification effects are driven largely by attentional differences, but facial EMG is still sensitive to the affective component of prejudice. Together, the two experiments demonstrate that, at least during the early stages of social categorization, people attend differentially to race. In particular, they appear to attend more to racial outgroups than they do to ingroups. Although people frequently exhibit negative affect in response to outgroup members, attentional differences to intergroup stimuli may be the immediate precursor to those more elaborated emotional responses. Consistent with this view was the finding across the two experiments that the self-reported measures of racial prejudice failed to moderate the attentional effects, although prejudice were related to facial EMG patterns. Another important component of this research was that Experiment 2 showed that such attentional effects were not limited just to having Whites view pictures of Blacks (vs. Whites), but also that Black participants exhibited an identical pattern when viewing pictures of Whites (vs. Blacks). Studies of intergroup perception have frequently restricted their scope to looking at the social categorization processes of one particular ethnic group. Our results suggest that this more limited approach may overlook the apparent universality of many intergroup phenomena.
The attentional effects found for startle modification in Experiment 1 and the secondary reaction time paradigm in Experiment 2 are consistent with studies that have demonstrated that negative social information is more interesting. When approach and aversion motivations are low, startle modification is more influenced by basic attentional demands. It is only when activation of these motivations reach a threshold that they have an impact on startle in the manner (Lang, Bradley, & Cuthbert, 1997). When using natural social categories it may be difficult to determine where this threshold is, whereas it is probably easier to find cases at the extremes (e.g., in places where intergroup conflict involves violence). Although we have no data about the relative levels of arousal of the pictures of Blacks and Whites used in these studies, our assumption is that in this laboratory context pictures of students were not very arousing. If the participants instead had been extremely prejudiced (none of the participants in Experiment 1 had MRS scores above the scale's midpoint), the startle modification effects may have been just the opposite--greater startle facilitation for the disliked racial group and smaller blinks for the favored group--to the obtained effects here, which would be consistent with the findings of Amodio et al. (2003). For example, Mahaffey, Bryan, and Hutchison (2005) examined antigay bias on startle modification and found that male participants who reported more antigay bias also showed greater startle facilitation to male nudes at a 4000-ms lead interval. And, the Phelps et al. (2000) fMRI study of racial prejudice suggested it is only when the amygdala is sufficiently activated, as it is during states of fear and anxiety that affective modification of startle occurs in an intergroup context. If dual control of startle depends on some sort of threshold, the development of an explanatory model of startle modification that incorporates this fact might be capable of more fully integrating the sometimes conflicting literatures on attentional and emotional influences on startle as well (see Filion et al., 1998).
Moreover, the methods used to measure attention in our studies revealed information about the time course of attentional effects in intergroup perception. The startle modification data showed that by 800 ms people attended more to ingroup members, and this attentional bias was sustained at 1500 s (according to Experiment 2), and even enhanced at 4500 s. Replication of this paradigm using other methodologies sensitive to attention, such as eye-tracking, should provide further support for these effects. Indeed, consistent with our findings, Kubota and Ito (2007) found N100 and P200 event-related potential effects at midline sites on the head that also suggested greater attention to a negative outgroup at an early stage, regardless of the target's facial expression of emotion, which instead appeared to be processed later than, and independently from, a social categorization based on race. In addition, their data also suggested that a marginal interaction effect of race and facial expression on N100s reflected an early vigilance effect. This increased vigilance may be part of a filtering process that requires more processing of outgroup members initially, but which is quickly attenuated once potential threat has been dismissed (Ito & Bartholow, 2009).
The results of this experiment have implications for investigations of affect that rely on psychophysiological measures in social neuroscience. The number of publications that used startle eyeblink modification with humans has been relatively high in the past two decades, due in large part to its widespread use as a marker of emotion. In a literature search using PsycINFO, we found that the number of investigations that have used facial EMG in this way is much smaller. The reasons for this difference in popularity between the two classes of measures are probably varied. Regardless, the results from the present study provide additional support for the notion that facial EMG can reliably index the affective responses that occur as a result of categorizations based on race, whereas startle modification may not. This support is especially compelling given the fact that the participants in this case were relatively low in prejudice. Combining fMRI and other neuroimaging technique with facial EMG should provide a clear window to investigate affective processes associated with the neuroscience of intergroup biases.
5. FUNDING
This research was partially supported by a fellowship to John Ryan from the Department of Psychology at Georgia State University and the Center for Behavioral Neuroscience, under the STC Program of the National Science Foundation under Agreement No. IBN-9876754. In addition, the facial electromyography data and friendliness ratings have been reported for the first 25 White participants that were tested in this experiment (Vanman et al., 1997).
6. ACKNOWLEDGMENTS
We thank Kimberle Kelly, Sean Rossi, Stephanie Gates, Lorretta Abbott, John Gundry, and Liberty Dillinger for their assistance in data collection and reduction. We thank Ottmar Lipp and members of the Centre for Research on Group Processes at the University of Queensland for their helpful comments on earlier drafts of this paper.
7. REFERENCES
Amodio, D. M., Harmon-Jones, E., & Devine, P. G. (2003). Individual differences in the activation and control of affective race bias as assessed by startle eyeblink responses and self-report. Journal of Personality and Social Psychology, 84, 738-853. [ Links ]
Bradley, M. M., Cuthbert, B. N., & Lang, P. J. (1993). Pictures as prepulses: Attention and emotion in startle modification. Psychophysiology, 30, 541-545. [ Links ]
Bradley, M. M., Cuthbert, B. N., & Lang, P. J. (1996). Picture media and emotion: Effects of a sustained affective content. Psychophysiology, 33, 662-670. [ Links ]
Brewer, M. B. (1988). A dual process model of impression formation. In T. S. Srull & R. S. Wyer (Eds.), Advances in social cognition: Vol. I: A dual process model of impression formation (pp. 1-36). Hillsdale, NJ: Erlbaum. [ Links ]
Brown, L. M., Bradley, M. M., & Lang, P. J. (2006). Affective reactions to pictures of ingroup and outgroup members. Biological Psychology, 71, 303-311. [ Links ]
Carretié, L., Mercado, F., Tapia, M., & Hinojosa, J. A. (2001). Emotion, attention, and the 'negativity bias,' studied through event-related brain potentials. International Journal of Psychophysiology, 41, 75-85. [ Links ]
Devine, P. G. (1989). Automatic and controlled processes in prejudice: The role of stereotypes and personal beliefs. In S. J. B. A. R. Pratkanis, & A. G. Greenwald (Eds.), Attitude, structure, and function (pp. 181-212). Hillsdale, NJ: Lawrence Erlbaum. [ Links ]
Donders, N. C., Correll, J., & Wittenbrink, B. (2008). Danger stereotypes predict racially biased attentional allocation. Journal of Experimental Social Psychology, 44, 1328-1333. [ Links ]
Filion, D. L., Dawson, M. E., & Schell, A. M. (1993). Modification of the acoustic startle-reflex eyeblink: A tool for investigating early and late attentional processes. Biological Psychology, 35, 185-200. [ Links ]
Filion, D. L., Dawson, M. E., & Schell, A. M. (1998). The psychological significance of human startle eyeblink modification: A review. Biological Psychology, 47, 1-43. [ Links ]
Fiske S. T. (1980). Attention and weight in person perception: The impact of negative and extreme behavior. Journal of Personality and Social Psychology, 38, 889-906. [ Links ]
Fridlund, A. J., & Cacioppo, J. T. (1986).Guidelines for human electromyographic research. Psychophysiology, 23, 567-589. [ Links ]
Guglielmi, R. S. (1999). Psychophysiological assessment of prejudice: Past research, current status, and future directions. Personality and Social Psychology Review, 3, 123-157. [ Links ]
Henry, P. J., & Sears, D. O. (2002).The symbolic racism 2000 scale. Political Psychology, 23, 253-283. [ Links ]
Ito, T. A., & Bartholow, B. D. (2009).The neural correlates of race. Trends in Cognitive Sciences, 13(12), 524-531. [ Links ]
Ito, T. A., & Cacioppo, J. T. (2000). Electrophysiological evidence of implicit and explicit categorization processes. Journal of Experimental Social Psychology, 36, 660-676. [ Links ]
Johnson, J. D., & Lecci, L. (2003). Assessing anti-White attitudes and predicting perceived racism: The Johnson-Lecci Scale. Personality and Social Psychology Bulletin, 29, 299-312. [ Links ]
Kubota, J. T., & Ito, T. A. (2007). Multiple cues in social perception: The time course of processing race and facial expression. Journal of Experimental Social Psychology, 43, 738-752. [ Links ]
Kubota, J. T., Banaji, M. R., & Phelps, E. A. (2012). The neuroscience of race. Nature Neuroscience, 15, 940-948. [ Links ]
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997). Motivated attention: Affect, activation, and action. In P. J. Lang, R. F. Simons, & M. T. Balaban (Eds.), Attention and orienting: Sensory and motivational processes (pp. 97-135). Mahwah, NJ: Lawrence Erlbaum. [ Links ]
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2005). International Affective Picture System (IAPS): Affective ratings of pictures and instruction manual. Technical Report no.A-6. Gainesville, FL: University of Florida. [ Links ]
Lipp, O. V., Siddle, D. A. T., & Dall, P. J. (1997).The effect of emotional and attentional processes on blink startle modulation and on electrodermal responses. Psychophysiology, 34, 340-347. [ Links ]
Luck, S. J., & Vecera, S. P. (2002). Attention. In H. Pashler & S. Yantis (Eds.), Stevens' handbook of experimental psychology. Vol. I: Sensation and perception (pp. 235-286). New York, NY: Wiley. [ Links ]
Mahaffey, A. L., Bryan, A., & Hutchison, K. E. (2005). Using startle eye blink to measure the affective component of antigay bias. Basic and Applied Social Psychology, 35, 537-545. [ Links ]
Mallan, K. M., & Lipp, O. V. (2007). Does emotion modulate the blink reflex in human conditioning? Startle potentiation during pleasant and unpleasant cues in the picture-picture paradigm. Psychophysiology, 44, 737-748. doi:10.1111/j.1469-8986.2007.00541.x. [ Links ]
Mallan, K. M., Sax, J., & Lipp, O. V. (2009). Verbal instruction abolishes fear conditioned to racial out-group faces. Journal of Experimental Social Psychology, 45, 1303-1307. [ Links ]
McConahay, J. B. (1986). Modern racism, ambivalence, and the modern racism scale. In J. F. Dovidio & S. L. Gaertner (Eds.), Prejudice, discrimination, and racism (pp. 91-126). San Diego, CA: Academic Press. [ Links ]
Molenberghs, P. (2013). The neuroscience of in-group bias. Neuroscience and Biobehavioral Reviews, 37(8), 1530-1536. [ Links ]
Mullen, B., Brown, R., & Smith, C. (1992).Ingroup bias as a function of salience, relevance, and status: An integration. European Journal of Social Psychology, 22, 103-122. [ Links ]
Oakes, P. & Turner, J. C. (1986). Distinctiveness and the salience of social category memberships: Is there an automatic perceptual bias towards novelty? European Journal of Social Psychology, 16, 325-344. [ Links ]
Öhman, A. (1997). As fast as the blink of an eye: Evolutionary preparedness for preattentive processing of threat. In P. J. Lang, R. F. Simons, & M. Balaban (Eds.), Attention and orienting: Sensory and motivational processes (pp. 165-184). Mahwah, NJ: Erlbaum. [ Links ]
Öhman, A. (2005). Conditioned fear of a face. A prelude to ethnic enmity? Science, 309(5735), 711-713. [ Links ]
Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning. Psychological Review, 108(3), 483-522. [ Links ]
Phelps, E. A., O'Connor, K. J., Cunningham, W. A., Funayama, E. S., Gatenby, J. C., Gore, J. C., Banaji, M. R. (2000). Performance on indirect measures of race evaluation predicts amygdala activation. Journal of Cognitive Neuroscience, 12, 729-738. [ Links ]
Posner, M. I. (1978). Chronometric explorations of the mind. Hillsdale, NJ: Erlbaum. [ Links ]
Price, T. F., Dieckman, L., & Harmon-Jones, E. (2012). Embodying approach motivation: Body posture influences startle eyeblink and event-related potential responses to appetitive stimuli. Biological Psychology, 90, 211-217. [ Links ]
Simons, R. F., & Zelson, M. F. (1985). Engaging visual stimuli and reflex modification. Psychophysiology, 22, 44-49. [ Links ]
Tassinary, L. G., Cacioppo, J. T., & Vanman, E.J. (2007). The skeletomotor system: Surface electromyography. J. T. Cacioppo, L. G. Tassinary, & G. G. Berntson (Eds.), Handbook of Psychophysiology (pp. 267-299). Cambridge: Cambridge University Press. [ Links ]
Trawalter, S., Todd, A. R., Baird, A. A., & Richeson, J. A. (2008). Attending to threat: Race-based patterns of selective attention. Journal of Experimental Social Psychology, 44, 1322-1327. [ Links ]
Van Bavel, J. J., & Cunningham, W. A. (2011). A social neuroscience approach to self and social categorisation: A new look at an old issue. European Review of Social Psychology, 21, 237-284. [ Links ]
Vanman, E. J., Boehmelt, A. H., Dawson, M. E., & Schell, A. M. (1996). The varying time courses of attentional and affective modulation of the startle eyeblink reflex. Psychophysiology, 33, 691-697. [ Links ]
Vanman, E. J., Paul, B. Y., Ito, T. A., & Miller, N. (1997).The modern face of prejudice and structural features that moderate the effect of cooperation on affect. Journal of Personality and Social Psychology, 73, 941-959. [ Links ]
Vanman, E. J., Saltz, J., Nathan, L. R., & Warren, J. A. (2004). Racial discrimination by low-prejudiced Whites: Facial movements as implicit measures of attitudes related to behavior. Psychological Science, 15, 711-714. [ Links ]
Witvliet, C. V., & Vrana, S. R. (1995). Psychophysiological responses as indices of affective dimensions. Psychophysiology, 32, 436-443. [ Links ]
