










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInternational Journal of Psychological Research
Print version ISSN 2011-2084
int.j.psychol.res. vol.6 no.spe Medellín Oct. 2013
Potential interactions between oxytocin receptor system (OXTR) and candidate genes associated to psychopathy
Probables interacciones entre el receptor de oxitocina (OXTR) y genes candidatos asociados a la psicopatía
Jorge Mauricio Cuartas Ariasa,b,* and Carlos Alberto López Jaramillob
a Faculty of Psychology, Universidad de San Buenaventura Medellín, Colombia.
b Department of Psychiatry, Faculty of Medicine, Universidad de Antioquia, Medellín, Colombia.
* Corresponding author. Mauricio Cuartas, Faculty of Psychology, Universidad de San Buenaventura, Medellin, Colombia, Mauricio.cuartas@usbmed.edu.co Tel: (57-4) 514 56 00, etx. 4245.
Recibido: 15-08-2013-Revisado: 02-09-2013-Aceptado: 10-10-2013
ABSTRACT
Psychopathy is the result of a complex interaction between biological and environmental factors; both participate in the final expression of the phenotype and can modulate the variation regarding beginning, deterioration, and severity. Nevertheless, up to now it has not been possible to determine the circumscribed molecular track and its interaction with environmental factors that shape its expression. However, beyond the progresses regarding diagnostic evaluation, there are some difficulties in the construction of a neurobiological model that contributes to differentiate the antisocial behavior. That is why exploring the molecular-genetic, biochemical and hormonal mechanisms involved in the different neuro-physiological tracks modulated by candidate genes could contribute to determine significantly how the neurobiological findings impact the individual differences in the expression of psychopathy. This article aims to go in depth into those neurobiological mechanisms underlying psychopathy from the expression and regulation of the oxytocinergic system.
Key Words: OXT, OXTR, Oxytocine, Psychopathy, candidate genes, endophenotype.
RESUMEN
La psicopatía es el resultado de una compleja interacción de factores biológicos y ambientales, ambos participan en la expresión final del fenotipo y pueden modular la variación en inicio, deterioro y severidad; sin embargo, hasta ahora no se han logrado determinar vías moleculares circunscritas y su interacción con los factores ambientales que modelen su expresión. No obstante, más allá de los avances en la evaluación diagnóstica, hay dificultades en la construcción de un modelo neurobiológico que contribuya a discriminar la conducta antisocial. Por lo tanto, explorar los mecanismos genético-moleculares, bioquímicos y hormonales implicados en las diferentes vías neurofisiológicas moduladas por genes candidatos, podría contribuir significativamente en determinar cómo los hallazgos neurobiológicos impactan las diferencias individuales en la expresión de la psicopatía. Este artículo tiene el propósito de profundizar en aquellos mecanismos neurobiológicos que subyacen a la psicopatía a partir de la expresión y regulación del sistema oxitocinérgico.
Palabras Clave: OXT, OXTR, oxitocina, psicopatía, genes candidatos, endofenotipo.
1. INTRODUCTION
Oxytocin (OXT) is a neuropeptide with a hormonal and neurotransmitter role associated initially with milk secretion and the reproductive function in women. It also has a relevant role in the central nervous system and a higher expression in women than men (H. J. Lee, Macbeth, Pagani, & Young, 2009).
Up to now, several studies have indicated the OXT effects in prosocial behaviors, especially in affiliation and empathy (Poulin, Holman, & Buffone, 2012), stress regulation, social interaction, rise and feel of confidence in others (Kumsta & Heinrichs, 2013; Lucas-Thompson & Holman, 2013), and moral behaviour (Pfeiffer, 2013).
These findings about the variable expression of OXT in a cerebral level can configure a neurobiological way to differentiate personality disorders from putative molecular markers that constitute a particular gene and molecular network
2. PERSONALITY AS A DIMENSION
Clonniger (1993) defined personality as a dynamic organization of different psychobiological systems integrated in a neurobiological, interpersonal and affective maturation on the individual that favors the social adaptation. In this perspective, individual differences integrate a dynamic array that incorporates the relation with oneself, the others and the environment, which has an underlying physiological system regulated by genes and the interaction between them and the environment (Aron et al., 2010; Stemmler & Wacker, 2010). Recent researches have reported association between candidate genes and personality disorders. Nevertheless, the clinical analyses to determine a group of symptom, which in diagnosis are gathered into factors to identify circumscribed genetic associations, have not been consistent. Currently, the strategy aims to evaluate different dimensional features that can be determined by several genes, which interact additively or not (J. M. C. Arias, & C. A. P. Acosta, 2011; M. C. Arias, 2011; Cuartas Arias et al., 2011). This alternative may be an interesting strategy to tackle the complexity of behavior. When a genetic polymorphism has been associated with a population, it could not be associated with another ethnic group, which can explain a huge quantity of misunderstanding genetic discoveries. Besides, the presence of pleiotropic and epistatic effects can contribute critically and differentially to the molecular basis of personality. Up to now, some methods to mapping genes in complex diseases, like studies of genetic association, have been advantageous. Even though a large clinical heterogeneity coexists for psychopathology, these designs can offer a greater statistical robustness when controlling the bias of sample selection and experimental errors in allelic and genotypic frequencies for a particular gene variant.
3. GENETIC ASSOCIATION STUDIES
Association studies are used to determine if there is an epidemiologic relation between one or more genetic variants and a trait, starting from how the frequencies of these variants differ among a group of cases and another of control. However, from this biological point of view, the association has three different interpretations: (i) The polymorphism has a causal role (direct association), (ii) The polymorphism is not causal, but it is in a linkage disequilibrium with the causal variant (indirect association), or (iii) The association is a product of stratification or mixing of population (Spurious association) (Cordell & Clayton, 2005).
The designs for genetic association use different strategies, among them we find the recruitment of large or nuclear families, but this method is difficult, expensive and slow. There also are designs that incorporate unrelated individuals, who are much easier to recruit, but they are susceptible to population stratification. In this regard, there are not studies of families for Personality that have good experimental sensitivity or that have been replicated. These variations can be functional and related to the physiopathology of the disease, but in most cases, they are used to map and locate really relevant sites. A single nucleotide polymorphisms (SNPs) is a variation of the ADN sequence that affects just one nucleotide (adenine (A), thymine (T), cytosine (C), or guanine (G)) of the genome. Each one of these variations should be at least in 1% of the population to be considered as a SNP (Muse, 2004). The SNP candidates are a direct test of association between a functional putative variation and the risk of disease. In this case, it is established in advance a candidate gene based on previous studies or biological experimental evidence. (Tenesa & Dunlop, 2006).
4. OXT GENE AND BEHAVIOR TRAITS AND DISORDERS
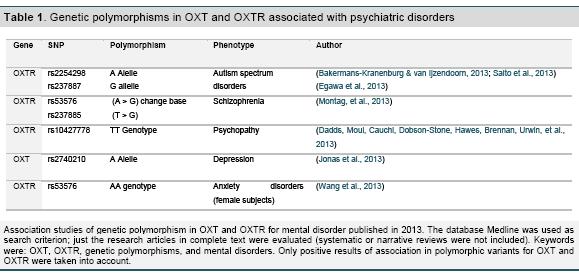
The OXT gene has been mapped in the large arm of the chromosome 20 (20p13), which codify for two proteins, oxytocin and neurophysin I. Both are packaged into the neurosecretory vesicles and carried by the axon to the terminal nerve of the neurohypophysis. OXT has three axons, two introns, and currently it has been documented just one receptor (OXTR), located in the chromosome 3 (3p25) that has four exons and three introns. Recently, different studies for OXT and OXTR genes have shown association in prosocial, emotional expression, individual differences and autism (Lerer et al., 2008; Tansey et al., 2010; Yrigollen et al., 2008), schizophrenia (Montag et al., 2013; Teltsh et al., 2011), psychopathy (Dadds, Moul, Cauchi, Dobson- Stone, Hawes, Brennan, Urwin, et al., 2013; Johansson et al., 2012), and so far it has been reported a complete genome for alcohol dependence with depressive syndrome in European - American descent (Edwards et al., 2012). Table 1 presents some of the key findings in 2013 for gene variants in OXT and OXTR in mental disorders and psychopathology (Table 1).
5. OXT GENE AND PSYCHOPATHY
In regard to the gene variants effects of OXT and OXTR, there are numerous studies that suggest the modeling of neurocognitive responses such as episodic and spatial memory (H. J. Lee, et al., 2009), decision making (Israel et al., 2009) and monitoring (Meyer-Lindenberg, 2008; Sala et al., 2011). The implications of the OXT system with the association of different genetic polymorphisms for different clinical phenotypes can delineate a way of molecular association that would allow infer a potential phenotype for psychological patterns closely related.
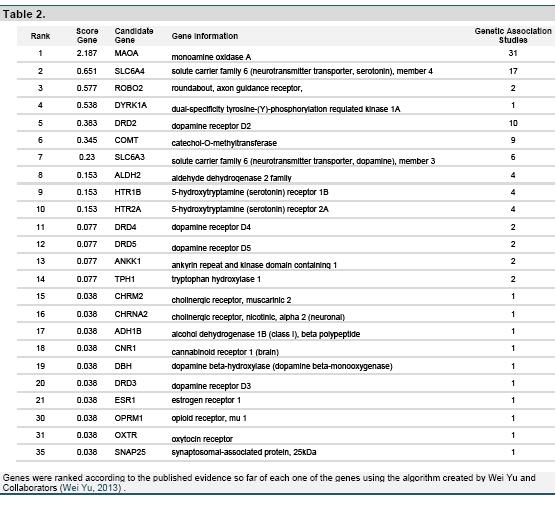
For psychopathology, from the dimensional perspective of personality and using as a model the observed finding for psychopathy, several studies have allowed to associate different SNPs in the OXTR gene and the differential expression of polymorphisms , in particular the G allele of rs 1042778 (Dadds, Moul, Cauchi, Dobson-Stone, Hawes, Brennan, & Ebstein, 2013; Dadds, Moul, Cauchi, Dobson-Stone, Hawes, Brennan, Urwin, et al., 2013), which can modulate processes of protein transduction and transcription in OXTR. This allelic variant in the receptor has been repeatedly associated with psychopathy and alterations in the inhibitory control (Johansson, et al., 2012; Malik, Zai, Abu, Nowrouzi, & Beitchman, 2012; Montag, Fiebach, Kirsch, & Reuter, 2011). The findings suggest that the presence of this polymorphism reduce the amygdala activation and affect the connectivity in the orbitofrontal and ventromedial cortex, both involved in the moral judgment and the emotional perception (Marsh et al., 2011; Marsh et al., 2008). However, several candidates genes have been referred to antisocial personality; until September 2013 in Huge Navigator 2.0 (An integrated, searchable knowledge base of genetic associations and human genome epidemiology), reported data for OXT and ORTR show their participation with a relatively marginal effect for the genetics associations studies (Wei Yu, 2013). Table 2. Nevertheless, the participation of the OXTR gene shows a pleiotropic effect that could suggest molecular regulation or modulation in the expression of phenotype, which contributes to evaluate the clinical dimension in psychopathy.
The neurobiological way that helps us to infer the participation of the OXT system in the modulation of the serotonergic and dopaminergic systems that participate directly in personality disorders is still complex. Currently, we can describe genes or molecules of reference that could refine the molecular networks to infer the expression of patterns set that are typically in the aggressive and antisocial behavior. Even though psychopathy has more literature regarding the androgens and their implications in personality, it is interesting to review the OXT system and the relation with the bioavailability of estrogens, which have been previously associated with the expression of aggressive behaviors.
Over 20 years ago, Caldwell and collaborators (1989) determined how estrogens that are frequently involved in plasticity and synaptogenesis increase the mRNA levels in the lateral preoptic area, which could explain the reproductive behavior, so the social bond as well. Additionally, Estrogen receptor beta (ERb) in the raphe nuclei, which have as function the serotonin release (5-HT) in the brain, have been identified. Besides, it has been reported their contribution with a greater density of the serotonin receptors 2A (5-HT2A) (Stahl, 2001), which is one of the receptors more associated with the impulsive behavior. Also, estrogens can decrease the expression of the monoamine oxidase A (MAOA), which is one of the candidate genes with best experimental relation to antisocial behaviour (Caspi et al., 2002; S. Y. Lee, Chen, Chang, & Lu, 2013; Wei Yu, 2013).
With these findings, analyzing the contribution of the OXT system and how it could influence in the expression of other genes that were associated with psychopathy could be useful to find a differential expression pattern that can modulate the clinical spectrum in psychopathy. This assertion is in accordance with the theory of Belsky, which affirms that the increase of susceptibility when developing a psychopathology can be related to polymorphic variations of the OXTR gene (Belsky, 1997). This explanation, has been supported by numerous findings that associate genetic polymorphism with the social pattern expression that have evolved in humans to favor efficient strategies of cooperation, empathy, and moral behavior, which is a constitutive base to differentiate clinical features related to mental and personality disorders (Cacioppo, Cacioppo, & Boomsma, 2013). A way to analyze the OXTR gene and the polymorphisms associated to this gene to increase or decrease the functional expression of the serotonergic network could allow to describe a path to delve into the findings about child abuse by Caspi and collaborators, which determined that individuals who show the genotype for low activity in the MAOA enzyme had more probabilities of developing psychopathy (5-HT levels are regulated by the action of monoamine oxidase, MAO) (Caspi, et al., 2002; Malik, et al., 2012).
Additionally, it is interesting the relation between the 5-HT system and OXT for antisocial and aggressive behavior, several candidate genes that predominate in the serotonergic network (SLC6A4, HTR2A, HTR1B, HTR3A, HTR3B, HTR2C) are involved in psychopathy (Wei Yu, 2013), and the OXT system influences physiologically in the hormonal regulation in the hypothalamic- pituitary- adrenal and neurophysiologically in the bioavailability of serotonin.
6. OXTR AS A POTENTIAL ENDOPHENOTYPE
Endophenotypes are constituted as a quantitative pattern of disadvantage, vulnerability, susceptibility or risk of showing a particular phenotype that can contribute directly to the probability of developing a particular syndrome or pathology (J. M. C. Arias, & C. A. P. Acosta, 2011). At the beginning, the endophenotype concept was coined by Dr. Irving Gottesman to explain the presence of an intermediate phenotype or vulnerability marker (clinical, biochemical, neurophysiological, genetic or cognitive) that is relatively stable in the expression of a specific psychopathological pattern (Gottesman & Gould, 2003).
OXTR has a crucial role in the expression of critical traits strongly associated with psychopathy. Perhaps, the more important one in its diagnosis is impulsivity, and its pathophysiological correlative is in accordance with the aforementioned candidate genes. Impulsivity entails aggressive behaviors, affiliation and prosocial difficulties, and lack of empathy. Currently, it has been reported that OXTR participates in the regulation of receptors in HTR2A, which have been associated with impulse control changes and aggressive behavior (Zalsman et al., 2011).
In the other hand, OXTR comply with the criteria for heritability, stability and replicability that should have an endophenotype, which suggests that OXTR can be considered as an intermediate molecular phenotype that predominates or interacts in a relevant way in the neurobiology and the etiopathogeny of psychopathy: there is evidence of molecular expression with OXTR in this disorder. Although OXT levels are variable between genders, they establish a relatively stable biomolecular pattern modulating the expression of the most bizarre clinical patterns that are closely linked to psychopathy and impulsivity.
At this moment, more research is needed. There are not yet clear studies to assess if the changes in OXTR cosegregate with this disruptive pattern. However, we may infer that OXTR polymorphisms can modulate a biochemical and genetic network associated with typical psychopathy characteristics, especially those that show lack of empathy, moral construct deficits and increased impulsivity. Furthermore, although the oxytocin effect has been frequently associated with prosocial and altruistic behaviors, there are others (such as the altruistic punishment practiced in social contexts and clung to human cooperation in cooperative patterns searching) that display aggressive behaviors and negative emotions in social competence.
Summarizing, it might be necessary to remodel a gene network that incorporates the functional load of the OXTR gene from its implications in genetic architecture of neural systems. Currently, there are few findings that allow discriminating how a specific polymorphism in the OXTR gene affects positively or negatively the expression or interacts directly with the molecular dynamic of other genes involved in antisocial behavior. It is possible that a best approach to the impact of the OXT system requires refining more dimensional diagnosis methods in psychopathology, analyzing putative molecular networks that could modulate the transcriptional efficiency of genetic polymorphisms reported, and incorporating methods of systems biology that allow refining epistatic processes into a molecular level underlying the differential expression of the phenotype of interest.
7. REFERENCES
Arias, J. M. C., & Acosta, C. A. P. (2011). Psychobiology and candidate endophenotypes in psychopathy. International Journal of Psychological Research, 4(1), 92-101. [ Links ]
Arias, M. C. (2011). Functional enviromics and personality disorders. International Journal of Psychological Research, 4(2), 5-8. [ Links ]
Aron, A., Ketay, S., Hedden, T., Aron, E. N., Rose Markus, H., & Gabrieli, J. D. (2010). Temperament trait of sensory processing sensitivity moderates cultural differences in neural response. [Research Support, N.I.H., Extramural]. Social cognitive and affective neuroscience, 5(2-3), 219-226. doi: 10.1093/scan/nsq028. [ Links ]
Bakermans-Kranenburg, M. J., & van Ijzendoorn, M. H. (2013). A sociability gene? Meta-analysis of oxytocin receptor genotype effects in humans. Psychiatric genetics. doi: 10.1097/YPG.0b013e3283643684. [ Links ]
Belsky, J. (1997). Variation in susceptibility to rearing influences: an evolutionary argument. Psychol Inquiry, 8, 182-186. [ Links ]
Cacioppo, J. T., Cacioppo, S., & Boomsma, D. I. (2013). Evolutionary mechanisms for loneliness. Cognition & emotion. doi: 10.1080/02699931.2013.837379. [ Links ]
Caldwell, J. D., Brooks, P. J., Jirikowski, G. F., Barakat, A. S., Lund, P. K., & Pedersen, C. A. (1989). Estrogen alters oxytocin mRNA levels in the preoptic area. Journal of neuroendocrinology, 1(4), 273-278. doi: 10.1111/j.1365-2826.1989.tb00115.x. [ Links ]
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., ... Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children. [Research Support, Non-U.S. Gov't Research Support, U.S. Gov't, P.H.S.]. Science, 297(5582), 851-854. doi: 10.1126/science.1072290. [ Links ]
Clonniger, C., Rakic, S. y Pezybeckt, D. (1993). A psychobiological model of temperament and character. Archives of General Psychiatric., 50(975-990). [ Links ]
Cordell, H. J., & Clayton, D. G. (2005). Genetic association studies. Lancet, 366(9491), 1121-1131. doi: S0140-6736(05)67424-7 [pii]10.1016/S0140-6736(05)67424-7. [ Links ]
Cuartas Arias, J. M., Palacio Acosta, C. A., Valencia, J. G., Montoya, G. J., Arango Viana, J. C., Nieto, O. C., ... Ruiz-Linares, A. (2011). Exploring epistasis in candidate genes for antisocial personality disorder. [Research Support, Non-U.S. Gov't]. Psychiatric genetics, 21(3), 115-124. doi: 10.1097/YPG.0b013e3283437175. [ Links ]
Dadds, M. R., Moul, C., Cauchi, A., Dobson-Stone, C., Hawes, D. J., Brennan, J., & Ebstein, R. E. (2013). Methylation of the oxytocin receptor gene and oxytocin blood levels in the development of psychopathy. Development and psychopathology, 1-8. doi: 10.1017/S0954579413000497. [ Links ]
Dadds, M. R., Moul, C., Cauchi, A., Dobson-Stone, C., Hawes, D. J., Brennan, J., ... Ebstein, R. E. (2013). Polymorphisms in the oxytocin receptor gene are associated with the development of psychopathy. Development and psychopathology, 1-11. doi: 10.1017/S0954579413000485. [ Links ]
Edwards, A. C., Aliev, F., Bierut, L. J., Bucholz, K. K., Edenberg, H., Hesselbrock, V., ... Dick, D. M. (2012). Genome-wide association study of comorbid depressive syndrome and alcohol dependence. [Research Support, N.I.H., Extramural]. Psychiatric genetics, 22(1), 31-41. doi: 10.1097/YPG.0b013e32834acd07. [ Links ]
Egawa, J., Watanabe, Y., Endo, T., Tamura, R., Masuzawa, N., & Someya, T. (2013). Association between OXTR and clinical phenotypes of autism spectrum disorders. [Letter]. Psychiatry research, 208(1), 99-100. doi: 10.1016/j.psychres.2012.11.007. [ Links ]
Gottesman, II, & Gould, T. D. (2003). The endophenotype concept in psychiatry: etymology and strategic intentions. [Research Support, U.S. Gov't, P.H.S. Review]. The American journal of psychiatry, 160(4), 636-645. [ Links ]
Israel, S., Lerer, E., Shalev, I., Uzefovsky, F., Riebold, M., Laiba, E., ... Ebstein, R. P. (2009). The oxytocin receptor (OXTR) contributes to prosocial fund allocations in the dictator game and the social value orientations task. [Research Support, Non-U.S. Gov't]. PloS one, 4(5), e5535. doi: 10.1371/journal.pone.0005535. [ Links ]
Johansson, A., Westberg, L., Sandnabba, K., Jern, P., Salo, B., & Santtila, P. (2012). Associations between oxytocin receptor gene (OXTR) polymorphisms and self-reported aggressive behavior and anger: Interactions with alcohol consumption. [Research Support, Non-U.S. Gov't]. Psychoneuroendocrinology, 37(9), 1546-1556. doi: 10.1016/j.psyneuen.2012.02.009. [ Links ]
Jonas, W., Mileva-Seitz, V., Girard, A. W., Bisceglia, R., Kennedy, J. L., Sokolowski, M., ... Steiner, M. (2013). Genetic variation in oxytocin rs2740210 and early adversity associated with postpartum depression and breastfeeding duration. Genes, brain, and behavior. doi: 10.1111/gbb.12069. [ Links ]
Kumsta, R., & Heinrichs, M. (2013). Oxytocin, stress and social behavior: neurogenetics of the human oxytocin system. [Research Support, Non-U.S. Gov't Review]. Current opinion in neurobiology, 23(1), 11-16. doi: 10.1016/j.conb.2012.09.004. [ Links ]
Lee, H. J., Macbeth, A. H., Pagani, J. H., & Young, W. S., 3rd. (2009). Oxytocin: the great facilitator of life. [Research Support, N.I.H., Extramural Research Support, N.I.H., Intramural Review]. Progress in neurobiology, 88(2), 127-151. doi: 10.1016/j.pneurobio.2009.04.001. [ Links ]
Lee, S. Y., Chen, S. L., Chang, Y. H., & Lu, R. B. (2013). Variation of types of alcoholism: Review and subtypes identified in Han Chinese. Progress in neuro-psychopharmacology & biological psychiatry, 48C, 36-40. doi: 10.1016/j.pnpbp.2013.09.013. [ Links ]
Lerer, E., Levi, S., Salomon, S., Darvasi, A., Yirmiya, N., & Ebstein, R. P. (2008). Association between the oxytocin receptor (OXTR) gene and autism: relationship to Vineland Adaptive Behavior Scales and cognition. [Research Support, Non-U.S. Gov't]. Molecular psychiatry, 13(10), 980-988. doi: 10.1038/sj.mp.4002087. [ Links ]
Lucas-Thompson, R. G., & Holman, E. A. (2013). Environmental stress, oxytocin receptor gene (OXTR) polymorphism, and mental health following collective stress. [Research Support, Non-U.S. Gov't Research Support, U.S. Gov't, Non-P.H.S.]. Hormones and behavior, 63(4), 615-624. doi: 10.1016/j.yhbeh.2013.02.015. [ Links ]
Malik, A. I., Zai, C. C., Abu, Z., Nowrouzi, B., & Beitchman, J. H. (2012). The role of oxytocin and oxytocin receptor gene variants in childhood-onset aggression. [Research Support, Non-U.S. Gov't]. Genes, brain, and behavior, 11(5), 545-551. doi: 10.1111/j.1601-183X.2012.00776.x. [ Links ]
Marsh, A. A., Finger, E. C., Fowler, K. A., Jurkowitz, I. T., Schechter, J. C., Yu, H. H., ... Blair, R. J. (2011). Reduced amygdala-orbitofrontal connectivity during moral judgments in youths with disruptive behavior disorders and psychopathic traits. [Research Support, N.I.H., Intramural]. Psychiatry research, 194(3), 279-286. doi: 10.1016/j.pscychresns.2011.07.008. [ Links ]
Marsh, A. A., Finger, E. C., Mitchell, D. G., Reid, M. E., Sims, C., Kosson, D. S., ... Blair, R. J. (2008). Reduced amygdala response to fearful expressions in children and adolescents with callous-unemotional traits and disruptive behavior disorders. [Research Support, N.I.H., Intramural]. The American journal of psychiatry, 165(6), 712-720. doi: 10.1176/appi.ajp.2007.07071145. [ Links ]
Meyer-Lindenberg, A. (2008). Impact of prosocial neuropeptides on human brain function. [Review]. Progress in brain research, 170, 463-470. doi: 10.1016/S0079-6123(08)00436-6. [ Links ]
Montag, C., Brockmann, E. M., Bayerl, M., Rujescu, D., Muller, D. J., & Gallinat, J. (2013). Oxytocin and oxytocin receptor gene polymorphisms and risk for schizophrenia: a case-control study. [Research Support, Non-U.S. Gov't]. The world journal of biological psychiatry : the official journal of the World Federation of Societies of Biological Psychiatry, 14(7), 500-508. doi: 10.3109/15622975.2012.677547. [ Links ]
Montag, C., Fiebach, C. J., Kirsch, P., & Reuter, M. (2011). Interaction of 5-HTTLPR and a variation on the oxytocin receptor gene influences negative emotionality. [Research Support, Non-U.S. Gov't]. Biological psychiatry, 69(6), 601-603. doi: 10.1016/j.biopsych.2010.10.026. [ Links ]
Muse, G. (2004). A Primer of Genome Science. Massachusetts: Sinauer Associates. [ Links ]
Pfeiffer, U. J. (2013). Oxytocin-not always a moral molecule. Frontiers in human neuroscience, 7, 10. doi: 10.3389/fnhum.2013.00010. [ Links ]
Poulin, M. J., Holman, E. A., & Buffone, A. (2012). The neurogenetics of nice: receptor genes for oxytocin and vasopressin interact with threat to predict prosocial behavior. Psychological science, 23(5), 446-452. doi: 10.1177/0956797611428471. [ Links ]
Saito, Y., Suga, M., Tochigi, M., Abe, O., Yahata, N., Kawakubo, Y., ... Yamasue, H. (2013). Neural correlate of autistic-like traits and a common allele in the oxytocin receptor gene. Social cognitive and affective neuroscience. doi: 10.1093/scan/nst136. [ Links ]
Sala, M., Braida, D., Lentini, D., Busnelli, M., Bulgheroni, E., Capurro, V., ... Chini, B. (2011). Pharmacologic rescue of impaired cognitive flexibility, social deficits, increased aggression, and seizure susceptibility in oxytocin receptor null mice: a neurobehavioral model of autism. [Research Support, Non-U.S. Gov't]. Biological psychiatry, 69(9), 875-882. doi: 10.1016/j.biopsych.2010.12.022. [ Links ]
Stahl, S. M. (2001). Effects of estrogen on the central nervous system. The Journal of clinical psychiatry, 62(5), 317-318. [ Links ]
Stemmler, G., & Wacker, J. (2010). Personality, emotion, and individual differences in physiological responses. Biological psychology, 84(3), 541-551. doi: 10.1016/j.biopsycho.2009.09.012. [ Links ]
Tansey, K. E., Brookes, K. J., Hill, M. J., Cochrane, L. E., Gill, M., Skuse, D., ... Anney, R. J. (2010). Oxytocin receptor (OXTR) does not play a major role in the aetiology of autism: genetic and molecular studies. [Research Support, Non-U.S. Gov't]. Neuroscience letters, 474(3), 163-167. doi: 10.1016/j.neulet.2010.03.035. [ Links ]
Teltsh, O., Kanyas-Sarner, K., Rigbi, A., Greenbaum, L., Lerer, B., & Kohn, Y. (2011). Oxytocin and vasopressin genes are significantly associated with schizophrenia in a large Arab-Israeli pedigree. The international journal of neuropsychopharmacology / official scientific journal of the Collegium Internationale Neuropsychopharmacologicum, 1-11. doi: 10.1017/S1461145711001374. [ Links ]
Tenesa, A., & Dunlop, M. G. (2006). Validity of tagging SNPs across populations for association studies. Eur J Hum Genet, 14(3), 357-363. doi: 5201554 [pii] 10.1038/sj.ejhg.5201554. [ Links ]
Wang, J., Qin, W., Liu, B., Zhou, Y., Wang, D., Zhang, Y., ... Yu, C. (2013). Neural mechanisms of oxytocin receptor gene mediating anxiety-related temperament. Brain structure & function. doi: 10.1007/s00429-013-0584-9. [ Links ]
Wei Yu, A. Y., Mindy Clyne, Anja Wulf, Marta Gwinn and Muin J. Khoury. (2013). HuGE Literature Finder. HuGE Navigator. Retrieved from http://www.hugenavigator.net/HuGENavigator/startPagePubLit.do/. [ Links ]
Yrigollen, C. M., Han, S. S., Kochetkova, A., Babitz, T., Chang, J. T., Volkmar, F. R., ... Grigorenko, E. L. (2008). Genes controlling affiliative behavior as candidate genes for autism. [Research Support, N.I.H., Extramural Research Support, Non-U.S. Gov't]. Biological psychiatry, 63(10), 911-916. doi: 10.1016/j.biopsych.2007.11.015. [ Links ]
Zalsman, G., Patya, M., Frisch, A., Ofek, H., Schapir, L., Blum, I., ... Tyano, S. (2011). Association of polymorphisms of the serotonergic pathways with clinical traits of impulsive-aggression and suicidality in adolescents: a multi-center study. [Multicenter Study Research Support, Non-U.S. Gov't]. The world journal of biological psychiatry : the official journal of the World Federation of Societies of Biological Psychiatry, 12(1), 33-41. doi: 10.3109/15622975.2010.518628. [ Links ]
