










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInternational Journal of Psychological Research
Print version ISSN 2011-2084
int.j.psychol.res. vol.9 no.2 Medellín Jul./Dec. 2016
https://doi.org/10.21500/20112084.2320
DOI: http://dx.doi.org/10.21500/20112084.2320
Research
Assessing Symmetry by Comparing the Acquisition of Symmetric and Nonsymmetric Conditional Relations in a Capuchin Monkey
Evaluación de Simetría por la Comparación de Relaciones Condicionales Simétricas y No-simétricas en Monos Capuchinos
Paulo S. D. Soares Filhoa,b*, Álvaro J. M. e Silvac, Saulo M. Velascob,d, Romariz S. Barrosc and Gerson Y. Tomanarib
a Universidad de San Buenaventura, Bogotá, Colombia
b Universidade de São Paulo, Brasil
c Universidade Federal do Pará, Brasil
d Centro Paradigma de Ciências do Comportamento, Brasil
* Corresponding author: Paulo Sergio Dillon Soares Filho, Programa de Psicología, Universidad de San Buenaventura Bogotá, B&aaáááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááááute;gotá, Colombia. Carrera 8H #172 -20. Email address: mailto: paulodillon@gmail.com.
Article history:
Received: 15-04-2016. Revised: 29-06-2016. Accepted: 12-08-2016
ABSTRACT
The present study presents a procedure to assess the property of symmetry by comparing the acquisition of conditional relations that are consistent and inconsistent with this property in a capuchin monkey (Sapajus spp.). One young male monkey underwent arbitrary matching-to-sample training. The experiment had four phases: Phase 1.1 (establishing A1B1 and A2B2 relations), Phase 1.2 (reinforcing B1A1 and B2A2 relations, consistent with the property of symmetry), Phase 2.1 (establishing A3B3 and A4B4 relations), and Phase 2.2 (reinforcing B3A4 and B4A3 relations, inconsistent with the property of symmetry). A comparison between Phase 1.2 (consistent) and Phase 2.2 (inconsistent) showed faster acquisition of consistent relations (B1A1 and B2A2) than inconsistent relations (B3A4 and B4A3). The results suggest that the established conditional discriminations may have the property of symmetry and confirm the potential of comparative analysis between the acquisition of conditional discriminations as a promising procedure to evaluate equivalence class formation in nonhuman subjects.
Key words: conditional discrimination; equivalence class formation; symmetry; Sapajus spp.
RESUMEN
Este estudio presenta un procedimiento para evaluar la propiedad de simetría a través de la comparación entre curvas de adquisición de relaciones condicionales consistentes y inconsistentes con la propiedad de simetría en un mono capuchino (Sapajus spp.). La investigación fue realizada en cuatro fases: Fase 1.1 (establecimiento de las relaciones A1B1 y A2B2), Fase 1.2 (reforzamiento de las relaciones B1A1 y B2A2, consistentes con la simetría), Fase 2.1 (reforzamiento de las relaciones A3B3 y A4B4), y Fase 2.2 (Reforzamiento de las relaciones B3A4 y B4A3, inconsistentes con la simetría). Al comparar las Fases 1.2 (consistente) y 2.2 (Inconsistente) fue observada una adquisición máápida de las relaciones consistentes que de las relaciones inconsistentes, sugiriendo que las relaciones condicionales establecidas poseen la propiedad de simetría. Reafirmando el procedimiento de comparación entre las curvas de adquisición de las relaciones condicionales como promisor para evaluar las propiedades de las clases de equivalencia.
Palabras clave: relaciones condicionales; formación de clases de equivalencia; simetría; sapajus spp.
1. INTRODUCTION
Equivalence class formation, as defined by Sidman and Tailby (1982; see also Sidman, 1994, 2000), is demonstrated by the emergence or establishment of new conditional relations that reflect the properties of reflexivity, symmetry, and transitivity. For example, given the conditional discriminations AB and BC, equivalence class formation would be demonstrated by the emergence of conditional relations AA, BB, and CC (Reflexivity), BA and CB (Symmetry), and AC (Transitivity).
The emergence of conditional relations, such as those that define equivalence class formation, have been repeatedly demonstrated in verbal humans (e.g., Devany, Hayes, & Nelson, 1986; Sidman & Cresson, 1973; Sidman & Tailby, 1982). However, the emergence of conditional relations has rarely been documented in nonhuman subjects or humans with severe developmental delays or significant impairments in verbal functioning (D'Amato, Salmon, Loukas, & Tomie, 1985; Devany et al., 1986; Hogan & Zentall, 1977; Sidman, Rauzin, Lazar, Cunningham, Tailby, & Carrigan, 1982).
Despite the different demonstrations of equivalence class formation between human and nonhuman subjects, some studies with nonhuman subjects have reported a few instances of emergent performance, such as generalized identity, which is usually treated as evidence of reflexivity1 (e.g., Barros, Galvão, & McIlvane, 2002; Kastak & Schusterman, 1994) and transitivity (e.g., D'Amato et al., 1985; Schusterman & Kastak, 1993; Yamamoto & Asano, 1995). Symmetry has received special attention in the literature because the results with nonhuman subjects have mostly been negative (Barros, Galvão, & Fontes, 1996; D'Amato et al., 1985; Dugdale & Lowe, 2000; Gray, 1966; Hogan & Zentall, 1977; Holmes, 1979; Lionello-DeNolf & Urcuioli, 2002; Lipkens, Kop, & Matthijs, 1988; Sidman et al., 1982; for a review see Lionello-DeNolf, 2009) and because some experiments have shown that one can facilitate the emergence of the other properties by directly training this property (e.g., Schusterman & Kastak, 1993; Yamamoto & Asano, 1995).
Recently, some studies that used a successive (Go/No-Go) matching procedure (e.g., Frank, 2007, Frank & Wasserman, 2005; Urcuioli, 2008; Vasconcelos & Urcuioli, 2011) reported the property of symmetry in pigeons, supporting the argument that procedural issues may be responsible for several of the prior failures to document symmetry in nonhumans. Following Sidman's proposition (Sidman, 1994, 2000), Urcuioli argued that such difficulties in demonstrating equivalence class formation in nonhuman subjects may be attributable to inappropriate control by stimulus locations during training and testing and thus may reflect more of a procedural issue than dependency on any direct multiple exemplar (or language) training (Urcuioli, 2008; Vasconcelos & Urcuioli, 2011).
In addition to issues concerning adventitious control by stimulus locations, another procedural issue that may be partially responsible for negative results with nonhuman subjects is the presentation of probe trials without programmed reinforcement (Brino, Galvão, & Barros, 2009; Galvão, Calcagno, & Sidman, 1992; Schusterman & Kastak, 1993). Emergent conditional relations are generally tested in the absence of programmed reinforcement (i.e., extinction) to ensure that performance is not explicitly trained during the test. However, the absence of programmed reinforcement may be a possible cause of digressions in stimulus control during the tests (Dube & McIlvane, 1996; Galvão et al., 1992) and may establish a discriminated repertoire that produces negative results during testing (Brino et al., 2009; Galvão et al., 2005). Extinction can also lead to the occurrence of resurgent behaviors (Epstein, 1983, 1985; Villas-Bôas, Murayama, & Tomanari, 2005; Wilson & Hayes, 1996), aggression (Azrin, Hutchinson, & Hake, 1966; Keller & Schoenfeld, 1950; Skinner, 1938), and an increase in behavioral variability (Antonitis, 1951; Lerman & Iwata, 1996). Additionally, test trials without reinforcement replicate the programmed consequences for incorrect responses during baseline training.
To avoid interference with the emergence of conditional relations by the absence of reinforcement, some alternative procedures have been developed, such as evaluating equivalence class properties under permanent differential reinforcement. One of these strategies consists of comparing the acquisition of conditional relations that are consistent with the conditional relations that are supposed to emerge (i.e., experimental condition) vs. inconsistent relations (i.e., control condition; e.g., D'Amato et al., 1985; Hogan & Zentall, 1977).
For example, Hogan and Zentall (1977) tested conditional relations that were consistent and inconsistent with the property of symmetry to assess such a property of conditional relations in pigeons. After training baseline conditional discriminations (A1B1 and A2B2), they divided the subjects into two groups. The first group was trained to choose the comparison stimulus that was consistent with the property of symmetry (A1, given the sample B1; A2, given the sample B2). The second group was trained to choose the comparison stimulus that was inconsistent with the property of symmetry (A2, given the sample B1; A1, given the sample B2). Evidence of symmetry would be confirmed by more precise and faster acquisition in the group that was given consistent training compared with the group that was given inconsistent training. The average group performance revealed differences between groups that were consistent with symmetry only in the first "testing" trials but not throughout the entire "testing" condition.
D'Amato et al. (1985, Experiment 1) also evaluated symmetry by utilizing consistent and inconsistent training in a single-subject experimental design. Each subject underwent training of conditional relations that were consistent and inconsistent with previously established baseline relations. Strong evidence of symmetry was found for two of the six subjects. The subjects responded with higher accuracy in the consistent training. Similar results were observed with one of the four remaining subjects in a follow-up study.
If group data are taken into account, such as in the study by Hogan and Zentall (1977), then the results that were reported by D'Amato et al. (1985, Experiment 1) would not have indicated symmetry. However, an examination of each subject's data individually suggests symmetry for two monkeys in the first evaluation and one monkey in the second evaluation. Despite the relatively small number of subjects, such results are noteworthy when considering the great amount of negative evidence of symmetry that has been reported in the literature with animals (c.f., Lionello-DeNolf, 2009).
Despite the benefits of a single-subject experimental design, exposure of the same subject to both consistent and inconsistent conditions with the same stimuli can create difficulties. The reinforcement of inconsistent relations (e.g., B1A2, B2A1, C1B2, and C2B1) may collapse the potentially established classes (e.g., A1B1C1 and A2B2C2) into a single large class (e.g., A1A2B1B2C1C2), resulting in levels of performance that are close to the level of chance during assessment procedures. This is what apparently happened with some subjects in the study by D'Amato et al. (1985, Experiment 1).
Velasco, Huziwara, Machado, and Tomanari (2010) developed an alternative strategy to assess the property of symmetry while reinforcing conditional relations in a single-subject design without collapsing the potential classes. They trained two separate sets of two baseline relations (A1B1 and A2B2; C1D1 and C2D2) with four pigeons. After reaching the accuracy criterion, they gave each subject symmetry-consistent training of one conditional relation that belonged to Set 1 (B1A1 and D1C1) together with training one new conditional relation that recombined stimuli that belonged to Sets 2 (D2A2 and B2C2). Such novel training was planned as a control condition without collapsing the potentially formed classes. Evidence of symmetry (i.e., faster acquisition of the symmetry-consistent relation) was not initially found. A second evaluation involved the symmetry-consistent relations B2A2 and D2C2 and novel relations D1A1 and B1C1. One pigeon showed strong evidence of symmetry, two pigeons showed some evidence of symmetry, and one pigeon did not show any evidence of symmetry. Velasco et al. argued that symmetry may not have emerged in the first evaluation because the subjects lacked a prerequisite discriminative repertoire, such as the absence of explicit stimulus-location control (as pointed out by Frank & Wasserman, 2005; Urcuioli, 2008). The prerequisite that might have been partially trained during symmetry training (i.e., shifting the order of the sample and comparison) may explain the improvement in performance during the second evaluation.
The main characteristic of the procedure that was reported by Velasco et al. (2010) is the exposure of each subject to both experimental (symmetry-consistent) and control (new) training conditions, without the possibility of collapsing classes. However, such a procedure required the training of four baseline conditional relations simultaneously, which can be difficult with some populations. Difficulty in training multiple conditional relations is one of the major limitations in research on stimulus class formation in nonhumans, particularly monkeys.
In summary, both studies (D'Amato et al., 1985; Velasco et al., 2010) reported evidence of symmetry in nonhuman subjects using a single-subject design with programmed reinforcement for the test trials. The goal of the present study was to evaluate a procedure to assess the property of symmetry in capuchin monkeys by combining elements of the procedures reported by D'Amato et al. and Velasco et al. The present study was part of a research program entitled Experimental School for Primates, which is dedicated to developing stimulus control procedures and evaluating the relational capabilities of capuchin monkeys (see Barros, Galvão, & McIlvane, 2002; Galvão, Soares Filho, Barros, & Souza, 2008).
We report data from symmetry-consistent and symmetry-inconsistent tests (with programmed reinforcement) using (i) a single-subject design, (ii) a limited number of conditional relations, and (iii) a modified matching-to-sample (MTS) procedure, in which sample and comparison stimuli can be displayed in any of nine locations in a 3 x 3 matrix (i.e., no correlation between stimulus position [e.g., central key] and stimulus function [sample]). Such procedural features were intended to overcome the following difficulties, respectively: (i) the limited number of experimental subjects available for research, considering that capuchin monkeys are wild animals that have been maintained in captivity, (ii) the difficulty in training more than three conditional relations simultaneously using standard MTS procedures in nonhuman subjects (Barros, Galvão, & McIlvane, 2002; McIlvane, Serna, Dube, & Stromer, 2000), and (iii) adventitious control by stimulus location.
After training the two baseline conditional relations (A1B1 and A2B2), one subject (a capuchin monkey) underwent training of symmetry-consistent relations (B1A1 and B2A2). Subsequently, two additional baseline relations (A3B3 and A4B4) were trained. The monkey then underwent training of symmetry-inconsistent relations (B3A4 and B4A3). The acquisition of symmetry-consistent and symmetry-inconsistent relations was compared.
2. METHOD
2.1 Subject
One adult male capuchin monkey (Sapajus spp.; Cotoh, M12) was included in the study. The monkey underwent simple and conditional discrimination training (identity and arbitrary MTS) but had no history of symmetry or any bidirectional relational training. The monkey was housed in a cage (2.5 x 2.5 x 2.5 m) with other monkeys of the same species at the Experimental School for Primates research program (Federal University of Pará, UFPA, Brazil).
The animal was fed once per day, approximately at 3:00 PM, and had free access to water. The balanced diet consisted of fruit, roots, protein, vegetables, and industrialized feed (Megazoo P18), providing all nutritional needs for monkeys. The housing conditions and animal handling were approved by the Ethics Committee in Research with Animals of the Institute of Biological Sciences of UFPA and by IBAMA (authorization no. 207419; unit code 381.201-4), in agreement with local and international ethical regulations.
2.2 Apparatus
The sessions were performed in an experimental chamber that was connected to a computer equipped with a touch screen monitor and an automatic food dispenser (Research Diets Dustless Precision Pellets, 190 mg). The food pellets were sugar-based with banana flavor. Stimulus presentation, response recording, and consequence presentation were automatically managed by software that was specifically designed for this purpose. An interface (developed in our laboratory) allowed the activation of devices, such as the automatic food dispenser, contingent on responding on the computer screen. The monkey's behavior was also video-recorded during the experimental sessions using a commercial surveillance and home monitoring system that was adapted for use in the laboratory.
We used a total of 10 two-dimensional black and white stimuli in bitmap format (alphanumerically referred to as A1, A2, A3, A4, A5, B1, B2, B3, B4, and B5) with dimensions of 93 x 93 pixels (see Figure 1).

2.3 General Procedure
Conditional discrimination training was performed using a zero-delay arbitrary MTS procedure. The sessions consisted of 48 trials and were conducted 5 days per week. Sample and comparison stimuli were presented randomly in any of nine positions in a 3 x 3 matrix on the computer screen. The monkey performed the discrimination tasks by directly touching the stimulus on the screen.
Each trial began with presentation of the sample stimulus in any of the nine positions. Touching the sample stimulus (initially under a fixed-ratio 3 [FR3] schedule and subsequently under a variable ratio 3 [VR3] schedule) produced removal of the sample stimulus and immediate (zero delay) presentation of two or three comparisons in any of the eight remaining positions. Touching the comparison stimulus (FR3) that was arbitrarily determined as correct (S+) produced food delivery, ended the trial, and initiated a 10 s intertrial interval (ITI). Touching the incorrect comparison stimulus (S-) produced the end of the trial and the ITI, without food delivery.
The experiment was divided into two phases, and each phase consisted of two sub-phases. The performance accuracy criterion in each sub-phase during the entire experiment was one session with a maximum of one error per conditional relation that was trained.
2.4 Specific Procedures
2.4.1 A1B1 and A2B2 baseline training
The monkey underwent training of two conditional relations (A1B1, A2B2). To obtain a three-choice arbitrary MTS baseline, after some training sessions we introduced a third conditional relation (A5B5) that had already been trained during the monkey's prior experimental history.
2.4.2 Symmetry-consistent (B1A1 and B2A2) evaluation
The monkey underwent training of symmetry-consistent conditional relations (B1A1 and B2A2). Additionally, training comprised A5B5 trials. Sample-comparison functions were not inverted for the third relation to maintain a three-choice matching procedure with minimal changes compared with baseline.
2.4.3 A3B3 and A4B4 baseline training
The monkey underwent training of two new arbitrary conditional relations (A3B3 and A4B4). The three-choice matching procedure was maintained by retaining the A1B1 discrimination as the third discrimination that was mastered in sub-phase 2.4.1 with discrimination A5B5.
2.4.4 Symmetry-inconsistent (B3A4 and B4A3) evaluation
The monkey underwent training of two new symmetry-inconsistent conditional relations (B3A4 and B4A3). Additionally, training comprised A1B1 trials (as in sub-phase 2.4.2).
After completing the four experimental sub-phases, we compared the acquisition of symmetry-consistent and symmetry-inconsistent conditional discriminations (sub-phases 2.4.2 and 2.4.4). Faster acquisition of symmetry-consistent conditional discrimination (sub-phase 2.4.2) compared with the acquisition of symmetry-inconsistent conditional discrimination (sub-phase 2.4.4) would demonstrate the property of symmetry in conditional baseline relations.
3. RESULTS AND DISCUSSION
The monkey successfully learned all forward (baseline) and backward (assessment) conditional relations. The monkey completed sub-phase 2.4.1 (A1B1 and A2B2 training) in 38 sessions, sub-phase 2.4.2 (symmetry-consistent training, B1A1 and B2A2) in 10 sessions, sub-phase 2.4.3 (A3B3 and A4B4 training) in six sessions, and sub-phase 2.4.4 (symmetry-inconsistent training, B4A3 and B3A4) in 28 sessions. Errors were equally distributed among stimuli throughout training (see Figure 2).
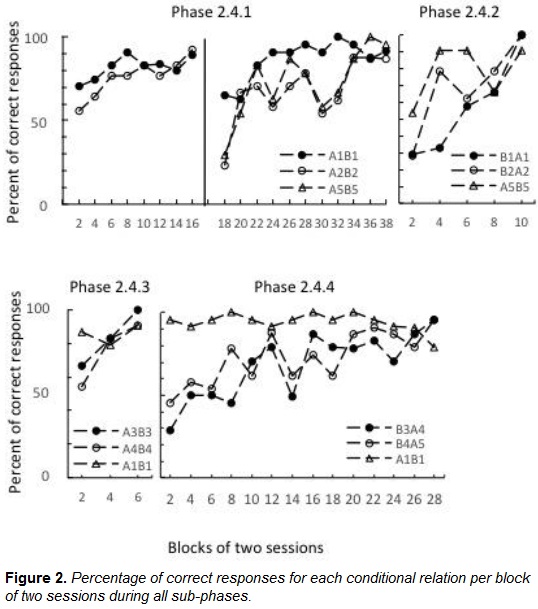
The monkey also exhibited faster acquisition of the symmetry-consistent relations than the symmetry-inconsistent relations (Figure 3). Such data provide evidence that the conditional relations that were initially trained had the property of symmetry and confirmed that the assessment procedure was able to evaluate the properties of equivalence classes in a nonhuman subject.
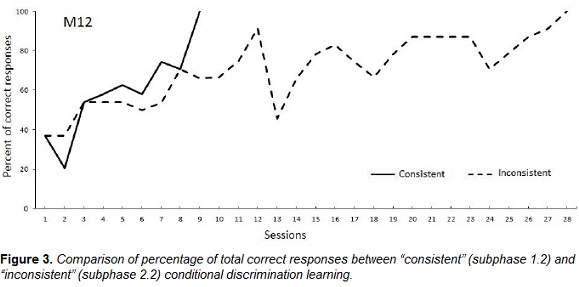
The present results document the property of symmetry in standard (zero-delay) MTS performance by a new-world primate (capuchin monkey) and extend previous findings of symmetry in other species that used assessment procedures that differed from standard testing in extinction (e.g., D'Amato et al., 1985; Hogan & Zentall, 1977; Velasco et al., 2010). Symmetry has also been successfully demonstrated in pigeons using successive (Go/No-Go) matching procedures, in which identity matching was trained together with arbitrary relations (e.g., Frank, 2007; Frank & Wasserman, 2005; Urcuioli, 2008; Vasconcelos & Urcuioli, 2011). The present study also extends the findings of Picanço and Barros (2015), with symmetry-compatible performance in a capuchin monkey in a successive (Go/No-Go) procedure. The study by Picanço and Barros (2015) was the only one that had previously documented the property of symmetry in a capuchin monkey by comparing symmetry-consistent and symmetry-inconsistent discrimination acquisition.
The present study, together with Picanço and Barros (2015), extend the evaluation of a novel procedure to assess the property of symmetry in a capuchin monkey using a single-subject design. Comparisons of the acquisition of symmetry-consistent and symmetry-inconsistent conditional relations were made across different baseline conditional relations. Such an approach, which differed from D'Amato et al. (1985), prevented collapse of the potential classes during symmetry evaluation (e.g., Velasco et al. 2010) and avoided the use of a high number of conditional discriminations.
Additionally, as part of the Experimental School for Primates research program, the experimental procedure described herein was designed to encourage coherence between trained stimulus control relations and those that are required during the tests (McIlvane et al., 2000). Among the procedures that may achieve this are three-choice MTS and the elimination of correlations between stimulus position and stimulus location (Barros, Galvão & McIlvane, 2002; Galvão et al., 2008).
One may suggest that the symmetry-consistent sub-phase 1.2 in the present study may configure multiple exemplar training and thus be responsible for the difficult in acquisition during the subsequent symmetry-inconsistent sub-phase 2.4.4 However, in contrast to Schusterman and Kastak (1993; who used a sea lion) and Yamamoto and Asano (1995; who used a chimpanzee that needed to train six symmetric relations to obtain any evidence of symmetry), the present study provided evidence of symmetry after having trained only two symmetric relations. The limited number of symmetric relations that were trained in the present study may suggest that equivalence class formation in nonhuman subjects is possible without multiple exemplar training (Devany et al., 1986; Hayes, 1989; Hayes, Barnes-Homes, & Roche, 2001).
The experimental procedure reported herein can clearly be improved to provide stronger evidence of symmetry in nonhuman primates. These results were obtained with only one subject and need to be replicated with more subjects and other species. Another experimental control that could be implemented in further related experiments is a control of the training order. This control might be done with the inclusion of a different training order group control (nonsymmetric training first, followed by symmetric training) or with the training of all relations together (symmetric and nonsymmetric; see Picanço and Barros, 2015). Additionally, we believe that a learning-set effect (Harlow, 1949) does not explain the results, considering that performance was less accurate in the fourth discrimination training condition (symmetry-inconsistent relations) after two baseline training conditions and one symmetry-consistent condition. If any learning-set effect occurred, then acquisition should have been faster as successive training conditions were implemented.
As mentioned above, some studies reported positive results of symmetry assessment by controlling the stimulus location during training and restricting the stimuli to only one possible location during testing using a successive Go/No-Go procedure (e.g., Frank, 2007; Frank & Wasserman, 2005; Urcuioli, 2008; Vasconcelos & Urcuioli, 2011). The present study implemented such stimulus-location control using multiple sample and comparison locations in a 3 x 3 matrix (as used by Brino et al., 2009; Barros et al., 2002; Barros, Galvão, & McIlvane, 2002; Galvão et al., 2005).
Our findings encourage the use of acquisition curve comparisons instead of traditional tests in extinction as a valid procedure to assess symmetry and other properties of equivalence relations in conditional and discrimination in nonhuman subjects (see Huziwara, Velasco, Tomanari, de Souza, & Machado, 2012; Velasco et al., 2010).
Foot notes
1 Generalized identity has traditionally been considered evidence of reflexivity (Dube, McIlvane, & Green, 1992; Sidman & Tailby, 1982; Sidman et al., 1982). However, its validity has recently been questioned, and conceptual theories and findings have been produced with the objective of distinguishing the two phenomena: generalized identity and reflexivity (e.g., Sweeney & Urcuioli, 2010; Urcuioli & Swisher, 2012).
4. REFERENCES
Antonitis, J. J. (1951). Response variability in the white rat during conditioning, extinction, and reconditioning. Journal of Experimental Psychology, 42, 273-281. doi: 10.1037/h0060407. [ Links ]
Azrin, N. H., Hutchinson, R. R., & Hake, D. F. (1966). Extinction-induced aggression. Journal of the Experimental Analysis of Behavior, 9, 191-204. doi: 10.1901/jeab.1966.9-191. [ Links ]
Barros, R. S., Galvão, O. F., & Fontes, J. C. (1996). Um teste de simetria após treino de discriminações condicionais de posição com macaco Ateles paniscus paniscus [A symmetry test after training position conditional discriminations with macacus Ateles paniscus paniscus]. Acta Comportamentalia, 4, 181-204. [ Links ]
Barros, R. S., Galvão, O. F., & McIlvane, W. J. (2002). The search for relational learning capacity in Cebus apella: a programmed "educational" approach. In: S. Soraci, Jr. & K. Murata-Soraci (Eds.), Visual information processing (pp. 223-245). Westport: Praeger. [ Links ]
Brino, A. L., Galvão, O. F., & Barros, R. S. (2009). Successive identity matching to sample tests without reinforcement in Cebus apella. Ciências e Cognição, 14, 2-11. Retrieved from: http://cienciasecognicao.org/revista/index.php/cec/article/view/19. [ Links ]
D'Amato, M. R., Salmon, D. P., Loukas, E., & Tomie, A. (1985). Symmetry and transitivity of conditional relations in monkeys (Cebus apella) and pigeons (Columbia livia). Journal of the Experimental Analysis of Behavior, 44, 35-47. doi: 10.1901/jeab.1985.44-35. [ Links ]
Devany, J. M., Hayes, S. C., & Nelson, R. O. (1986). Equivalence class formation in language-able and language-disabled children. Journal of the Experimental Analysis of Behavior, 46, 243-257. doi: 10.1901/jeab.1986.46-243. [ Links ]
Dube, W. V. & McIlvane, W. J. (1996). Some implications of a stimulus control topography analysis for emergent behavior and stimulus classes. In: T. R. Zentall & P. M. Smeets (Eds.), Stimulus class formation in humans and animals (pp. 197-218). Amsterdam: Elsevier. [ Links ]
Dube, W. V., McIlvane, W. J., & Green, G. (1992). An analysis of generalized identity matching-to-sample test procedures. Psychological Record, 42, 17-28. [ Links ]
Dugdale, N. & Lowe, C. F. (2000). Testing for symmetry in conditional discriminations of language-trained chimpanzees. Journal of the Experimental Analysis of Behavior, 73, 5-22. doi: 10.1901/jeab.2000.73-5. [ Links ]
Epstein, R. (1983). Resurgence of previously reinforced behavior during extinction. Behaviour Analysis Letters, 3, 391-397. [ Links ]
Epstein, R. (1985). Extinction-induced resurgence: preliminary investigations and possible applications. Psychological Record,35, 143-153. [ Links ]
Frank, A. J. (2007). An examination of the temporal and spatial stimulus control in emergent symmetry in pigeons. Unpublished doctoral dissertation, University of Iowa. Retrieved from: http://ir.uiowa.edu/cgi/viewcontent.cgi?article=1342&context=etd&unstamped=1. [ Links ]
Frank, A. J. & Wasserman, E. A. (2005). Associative symmetry in the pigeon after successive matching-to-sample training. Journal of the Experimental Analysis of Behavior, 84, 147-165. doi: 10.1901/jeab.2005.115-04. [ Links ]
Galvão, O. F., Calcagno, S., & Sidman, M. (1992). Testing for emergent performance in extinction. Experimental Analysis of Human Behavior Bulletin, 10, 18-20. [ Links ]
Galvão, O. F., Barros, R. S., Santos, J. R., Brino, A. L. F., Brandão, S., Lavratti, C. M., Dube, W. V., & McIlvane, W. J. (2005). Extent and limits of the matching concept in Cebus apella: a matter of experimental control? Psychological Record, 55, 219-232. [ Links ]
Galvão, O. F., Soares Filho, P. S. D., Barros, R. S., & Souza, C. B. A. (2008). Matching-to-sample as a model of symbolic behavior for bio-behavioral investigation. Reviews in the Neurosciences, 19, 149-156. [ Links ]
Gray, L. (1966). Backward association in pigeons. Psychonomic Science, 4, 333-334. [ Links ]
Harlow, H. F. (1949). The formation of learning sets. Psychological Review, 56, 51-65. doi: 10.1037/h0062474 [ Links ]
Hayes, S. C. (1989). Nonhumans have not yet shown stimulus equivalence. Journal of the Experimental Analysis of Behavior, 51, 385-392. doi: 10.1901/jeab.1989.51-385. [ Links ]
Hayes, S. C., Barnes-Holmes, D., & Roche, B. (2001). Relational frame theory: a post-Skinnerian account of human language and cognition. New York: Plenum. [ Links ]
Hogan, D. E. & Zentall, T. R. (1977). Backward associations in pigeon. American Journal of Psychology, 90, 3-15. doi: 10.2307/1421635. [ Links ]
Holmes, P. W. (1979). Transfer of matching performance in pigeons. Journal of the Experimental Analysis of Behavior, 31, 103-114. Doi: 10.1901/jeab.1979.31-103. [ Links ]
Huziwara, E. M., Velasco, S. M., Tomanari, G. Y., de Souza, D. G., & Machado, A. (2012). Emergent relations in pigeons following training with temporal samples. Learning and Behavior, 40, 192-204. doi: 10.3758/s13420-012-0097-z. [ Links ]
Kastak, D. & Schusterman, R. J. (1994). Transfer of visual identity matching-to-sample in two California sea lions (Zalophus californianus). Animal Learning and Behavior,22, 427-435. Retrieved from: http://link.springer.com/article/10.3758/BF03209162. [ Links ]
Keller, F. S. & Schoenfeld, W. N. (1950). Principles of psychology: a systematic text in the science of behavior. New York: Appleton-Century-Crofts. [ Links ]
Lerman, D. C. & Iwata, B. A. (1996). Developing a technology for the use of operant extinction in clinical settings: an examination of basic and applied research. Journal of Applied Behavior Analysis,29, 345-382. doi: 10.1901/jaba.1996.29-345. [ Links ]
Lionello-DeNolf, K. M. (2009). The search for symmetry: 25 years in review. Learning and Behavior,37, 188-203. doi: 10.3758/LB.37.2.188. [ Links ]
Lionello-DeNolf, K. M. & Urcuioli, P. J. (2002). Stimulus control topographies and tests of symmetry in pigeons. Journal of the Experimental Analysis of Behavior, 78, 467-495. doi: 10.1901/jeab.2002.78-467. [ Links ]
Lipkens, R., Kop, P. F. M., & Matthijs, W. (1988). A test for symmetry and transitivity in the conditional discrimination performances of pigeons. Journal of the Experimental Analysis of Behavior, 49, 395-409. doi: 10.1901/jeab.1988.49-395. [ Links ]
McIlvane, W. J., Serna, R. W., Dube, W. V., & Stromer, R. (2000). Stimulus control topography coherence and stimulus equivalence: reconciling test outcomes with theory. In: J. Leslie & D. E. Blackman (Eds.), Experimental and applied analysis of human behavior (pp. 85-110). Reno: Context Press. [ Links ]
Picanço, C. R. & Barros, R. S. (2015). Symmetry evaluation by comparing acquisition of conditional relations in successive (Go/No-Go) matching-to-sample training. Psychological Record, 65, 131-139. Retrieved from: http://link.springer.com/article/10.1007%2Fs40732-014-0096-x. [ Links ]
Schusterman, R. J. & Kastak, D. (1993). A California sea lion (Zalophus californianus) is capable of forming equivalence relations. Psychological Record,43, 823-839. [ Links ]
Sidman, M. (1994). Equivalence relations and behavior: a research story. Boston: Authors Cooperative. [ Links ]
Sidman, M. (2000). Equivalence relations and the reinforcement contingency. Journal of the Experimental Analysis of Behavior,74, 127-146. doi: 10.1901/jeab.2000.74-127. [ Links ]
Sidman, M. & Cresson, O., Jr. (1973). Reading and crossmodal transfer of stimulus equivalences in severe retardation. American Journal of Mental Deficiency,77(5), 515-523. Retrieved from: http://www.ncbi.nlm.nih.gov/pubmed/4267398. [ Links ]
Sidman, M. & Tailby, W. (1982). Conditional discrimination vs. matching to sample: an expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior,37, 5-22. doi: 10.1901/jeab.1982.37-5. [ Links ]
Sidman, M., Rauzin, R., Lazar, R., Cunningham, S., Tailby, W., & Carrigan, P. (1982). A search for symmetry in the conditional discriminations of rhesus monkeys, baboons, and children. Journal of the Experimental Analysis of Behavior,37, 23-44. doi: 10.1901/jeab.1982.37-23. [ Links ]
Skinner, B. F. (1938). The behavior of organisms: an experimental analysis. New York: Appleton-Century-Crofts. [ Links ]
Sweeney M. M. & Urcuioli, P. J. (2010). Reflexivity in pigeons. Journal of the Experimental Analysis of Behavior,94, 267-282. doi: 10.1901/jeab.2010.94-267. [ Links ]
Urcuioli, P. J. (2008). Associative symmetry, antisymmetry, and a theory of pigeons' equivalence-class formation. Journal of the Experimental Analysis of Behavior, 90, 257-282. doi: 10.1901/jeab.2008.90-257. [ Links ]
Urcuioli, P. J. & Swisher, M. (2012). Emergent identity matching after successive matching training: II. Reflexivity or transitivity? Journal of the Experimental Analysis of Behavior,97, 5-27. doi: 10.1901/jeab.2012.97-5. [ Links ]
Vasconcelos, M. & Urcuioli, P. J. (2011). Associative symmetry in a spatial-response paradigm. Behavioural Processes,86, 305-315. doi: 10.1016/j.beproc.2011.01.002. [ Links ]
Velasco, S. M., Huziwara, E. M., Machado, A., & Tomanari, G. Y. (2010). Associative symmetry by pigeons after few-exemplar training. Journal of the Experimental Analysis of Behavior,94(3), 283-295. doi: 10.1901/jeab.2010.94-283. [ Links ]
Villas-Bôas, A., Murayama, V. K., & Tomanari, G. Y. (2005). Ressurgência: conceitos e métodos que podem (ou não) contribuir para a análise do comportamento. In: H. J. Guilhardi & N. C. Aguirre (Eds.), Sobre comportamento e cognição: Vol. 15. Expondo a variabilidade (pp. 18-28). Santo André: ESETec. [ Links ]
Wilson, K. G. & Hayes, S. C. (1996). Resurgence of derived stimulus relations. Journal of the Experimental Analysis of Behavior,66, 267-281.doi: 10.1901/jeab.1996.66-267. [ Links ]
Yamamoto, J. & Asano, T. (1995). Stimulus equivalence in a chimpanzee (Pan troglodytes). Psychological Record,45, 3-21. [ Links ]
5. ACKNOWLEDGMENT
This research was supported by a Master Student Scholarship (FAPESPA) to Paulo S. D. Soares Filho, an Undergraduate Scholarship (PIBIC-CNPq UFPA) to Álvaro J. M. e Silva, a Post-Doctoral Scholarship (FAPESP) to Saulo M. Velasco, research support (CNPq) to Gerson Y. Tomanari, and a Research Productivity Grant (CNPq) to Romariz S. Barros. All of the authors are members of the National Institute of Science and Technology on Behavior, Cognition, and Teaching, supported by FAPESP (grant no. 08/57705-8) and CNPq (grant no. 573972/2008-7). The authors declare no conflicts of interest. We thank Klena Sarges for veterinarian assistance and Adilson Pastana for the excellent homecage care given to our monkeys.

