










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInternational Journal of Psychological Research
Print version ISSN 2011-2084
int.j.psychol.res. vol.9 no.2 Medellín Jul./Dec. 2016
https://doi.org/10.21500/20112084.2646
DOI: http://dx.doi.org/10.21500/20112084.2646
Research
Social context modulates food hoarding in Syrian hamsters
El contexto social modula el almacenamiento de comida en hamsters Sirios
Bibiana Montoyaa and Germán Gutiérreza*
a Departamento de Psicología, Universidad Nacional de Colombia, Bogotá, Colombia.
* Corresponding author: Germán Gutiérrez, Departamento de Psicología, Universidad Nacional de Colombia, Bogotá, Colombia Email Address: gagutierrezd@unal.edu.co
Article history:
Received: 24-05-2016. Revised: 13-06-2016. Accepted: 21-06-2016.
ABSTRACT
The effect of the presence of a con-specific in the temporal organization of food hoarding was studied in two varieties of Syrian hamster (Mesocricetus auratus): golden and long-haired. Four male hamsters of each variety were used. Their foraging behavior was observed during four individual and four shared trials in which animals were not competing for the same food source or territory. During individual trials, long-haired hamsters consumed food items directly from the food source, transporting and hoarding only remaining pieces. During shared trials, the long-haired variety hoarded food items before consumption, and increased the duration of hoarding trips, food handling in the storage, and cache size. Golden hamsters maintained the same temporal organization of hoarding behavior (i.e., hoarding food items before consumption) throughout both individual and shared trials. However, the golden variety increased handling time at the food source and decreased the duration of hoarding trips, the latency of hoarding and storing size throughout the shared trials. In Syrian hamsters, the presence of a con-specific may signal high probability of food source depletion suggesting that social pressures over food availability might facilitate hoarding behavior. Further studies are required to evaluate cost-benefit balance of food hoarding and the role of cache pilferage in this species.
Key words: hamster, foraging, social behavior, larder hoarding, food hoarding.
RESUMEN
Se estudió el efecto de la presencia de un conespecífico en la organización temporal en el almacenamiento de comida en dos variedades de hamsters sirios (Mesocricetus auratus): dorados y de pelo largo. Se utilizaron cuatro hamsters machos de cada variedad. Se observó su comportamiento de forrajeo durante cuatro ensayos individuales y cuatro compartidos, en los que los animales no competían por la misma comida o territorio. Durante los ensayos individuales, los hamsters de pelo largo consumieron comida directamente de la fuente, y transportaron y almacenaron la comida restante. Durante los ensayos compartidos, la variedad de pelo largo almacenó la comida antes de su consumo e incrementó la duración de sus viajes de almacenamiento, manipulación de comida en el almacén y el tamaño de la reserva. Los hamsters dorados mantuvieron la misma organización temporal de la conducta de almacenamiento (i.e., almacenamiento de comida antes de su consumo) tanto en los ensayos individuales como en los compartidos. Sin embargo, la variedad dorada incrementó el tiempo de manipulación en la fuente de comida y decrementó la duración de los viajes de almacenamiento, la latencia del almacenamiento y el tamaño de la reserva durante los ensayos compartidos. En los hamsters Sirios, la presencia de un conespecífico puede señalar una alta probabilidad de agotamiento de la fuente de comida lo que sugiere que las presiones sociales sobre la disponibilidad de comida pueden facilitar la conducta de almacenamiento. Se requieren otros estudios para evaluar el balance de costo-beneficio del almacenamiento y el papel del hurto de reservas en esta especie.
Palabras clave: hamster, forrajeo, conducta social, almacenamiento de comida.
1. INTRODUCTION
Food hoarding is a behavioral adaptation used by social insects like ants and bees (Holldobler & Wilson, 1990 ; Rinderer, Bolten, Harbo, & Collins, 1982), birds (Brodin, 2005 ; Shettleworth, 2001), rodents (Beck & Vander Wall, 2010 ; Yasuda, Miura & Nor Azman, 2000), and some marsupials (Dennis, 2003 ; Forget & Vander Wall, 2001) to cope with fluctuations in the availability of nutritional resources. Food hoarding requires the capacity to delay food consumption and, in turn, provides privileged access to food in both time and space, releasing the hoarder from pressures exerted by fluctuations in their physical and social environment (Jansen & Forget, 2001 ; Vander Wall, 1990; Vander Wall & Jenkins, 1987).
Some spatial variation has been observed within and between species regarding their food hoarding strategies. Food storage can be built in one place or several scattered places. Territorial species such as squirrels, marsupial rats, and beavers frequently use a central place located in the burrow or near it, where all the food items are stored (Smith & Reichman, 1984). Using the burrow for food storage reduces costs associated with searching for a new safe place and eventually relocating the items stored there. However, larder hoarding entails at least two risks: food stored there can be exposed to microbes, fungus and insects due to the relative humidity level of the borrow; and competitors using con-specific or hetero-specific nests as clues during food searching would be able to empty the individual only food storage (Jansen & Forget, 2001 ; Jenkins, Rothstein & Green, 1995). Some individuals aggressively defend their storage from the invasion of con-specifics which may entail energetic costs (Briggs & Vander Wall, 2004 ; Vander Wall & Jenkins, 1987). However, at least 26% of the seeds consumed by an individual during a season may come from foreign deposits and 25% of the seeds from its own deposits are taken by competitors (Gerhardt, 2005); even some individuals build false food storages in presence of con-specifics to reduce the stealing rate (Steele et al., 2001). Therefore, one of the variables that can determine food availability is the action of competitors (Alexander et al., 2005 ; Vander Wall & Jenkins, 1987).
To minimize the costs of building a central storage in the nest, food can also be stored in multiple scattered places located near the burrow, around the food source, or in the area between these two points (Jenkins, Rothstein & Green, 1995). Interestingly, individuals may exhibit flexibility in their hoarding strategies depending on environmental factors. Scatter hoarding can also help to counteract the impact of pilferage (Male & Smulders, 2007). Accordingly, willow tits (Parus montanus) use different spatial strategies to store food depending on social variables. Subordinates prefer to build deposits in multiple areas distant from their nest, while dominants tend to store food in only one place within their nesting area (Van Horik & Burns, 2007). Additionally, these birds make deposits near their food source in the absence of con-specifics but prefer locations far from their food source when con-specifics are present (Lahti et al., 1998). However, building scattered storages entails extra costs such as longer foraging trips, increased predation risk and seed loss by germination despite its potential usefulness against the actions of competitors (Steele et al., 2001).
In natural conditions adult Syrian hamsters (Mesocricetus auratus) are solitary, highly aggressive with con-specifics and the presence of males in a female's burrow could only be tolerated during the reproductive season. Syrian hamsters build larder hoards and defend them against competitors (Gismonoli, 1999 ; Lupfer, Frieman & Coonfield, 2003). Golden Syrian hamsters are the domestic variety most similar to the wild type at the phenotypic and behavioral level; while long-haired Syrian hamsters are a domestic variety selected by its long hair and low aggression level (Yoshizawa et al., 2015). The aim of the present study was to evaluate the extent to which changes in the social environment affect food hoarding strategies in two varieties of Syrian hamster. If intra-specific competence is one of the pressures promoting food hoarding in hamsters, it was expected that the presence of a con-specific would have an effect on the temporal organization of their food hoarding strategy.
2. METHODS
2.1 Subjects
Four golden and four long-haired Syrian male hamsters were provided by a commercial supplier. Animals were 30 days old upon their arrival to the Laboratory of Animal Learning and Behavior, Universidad Nacional de Colombia, and were housed in same variety groups for 60 days. At the beginning of the experiment hamsters were 90 days old. Animal had ad libitum access to laboratory rodent food and water. Additionally, 10 gr of peanuts (6 items) were provided to stimulate alimentary behavior during observational sessions.
At the arrival to the laboratory, experimental animals were exposed to 24 h darkness to revert the light-dark cycle. Reversed cycle during the study was 14:10 h light:dark with light starting at 22:00 hours.
2.2 Apparatus
Animals were housed in individual glass boxes 40 cm x 60 cm x 60 cm (width x length x height). One of the glass ends of the box was replaced by a wire wall. An identical glass box was attached to the wire end. The attached box was empty during individual observations and contained a con-specific during the shared observations. Food and water were available in plastic containers, water was placed in the middle of the housing box and the food container was hanged from the wire wall, 5 cm from the floor. Housing boxes were provided with a 2 cm bed of wood chips.
For behavioral records, housing boxes were divided into three equal adjacent rectangles (20 cm x 40 cm). In the first rectangle (zone A) the food source was located; in the middle of the second rectangle (zone B) water was available, and the third one was empty (zones C; Fig. 1).
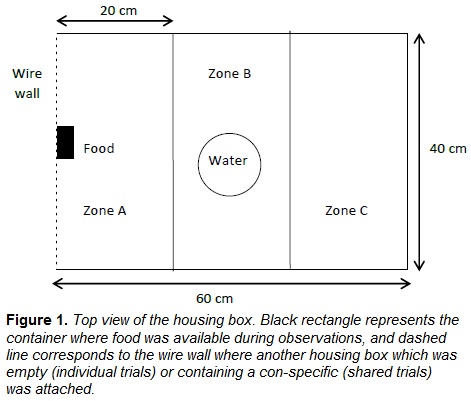
2.3 Procedure
Pilot observations were carried out twice a day (morning and afternoon) during 10 days with a different set of animals to establish the maximum time animals spent interacting continously with food resources. Pilot and experimental trials were performed exactly in the same way. Additionally, animals were exposed to laboratory conditions (i.e., food type, housing, photoperiod, temperature, humidity, background odors and noise) during 15 days previously starting the experimental trials to promote habituation to stimuli present in the new environment.
Experimental trials began the last hour of the light cycle when rodent food was removed and another housing box was attached to the focal animal's arena. At the beginning of the dark cycle, 10 gr of peanuts were available in the food container and foraging behavior was continuously recorded during 30 min. After this observation period, the remaining peanuts were removed and specific location and weight were recorded, food hoarded in the individual's cheek pouches was also removed. Finally, the housing box was cleaned with 50% acetic acid diluted in water to neutralize odors, the wood-chip bed was replaced and rodent food was placed once again in the food container. The second observation in a day took place four hours after the first one was finalized and was carried out using exactly the same procedure described above. A day of individual observations was followed by a day of shared observations where both attached boxes contained each a subject. Each type of observation was repeated once during non-consecutive trials.
2.4 Behavioral records
During individual and shared observations, trained observers recorded: i) the latency of food searching (period of time between the beginning of the observation and the first time the animal handled available food), ii) the latency of food hoarding (period of time between the beginning of the observation and the first time the animal transported food to a place different from the food source), iii) the latency of food consumption (period of time between the beginning of the observation and the first time the animal consumed a food item), iv) the duration of food handling (time spend by the animal handling food items with or without consuming them), v) the duration of hoarding trips (time spend by the animal transporting food items from the source to elsewhere) and vi) the size of food storage (weight in grams of all the food storage built by the subject).
Statistical analyses were performed using R 3.1.2 for Windows. Considering the small sample size, paired t tests were used to evaluate within subject differences when observed alone or with a con-specific and t tests were used to compare behavior between the golden versus the long-haired varieties during each observation phase.
3. RESULTS
3.1 Within subjects social effect
In the presence of a con-specific, long-haired Syrian hamsters reduced the latency of food searching and hoarding, while the latency of food consumption did not significantly change (Fig. 2 ; Table 1). Therefore, in presence of a con-specific, long-haired hamsters prioritized hoarding over food consumption. Food handling in the food source in absence of a con-specific (mean = 3.69 ± 0.24 min) did not differ when a con-specific was present (mean = 3.60 ± 0.22 min; Table 2). However, food handling in the storage was higher in shared conditions (individual mean = 2.68 ± 0.25 min, shared mean = 4.61 ± 0.30 min; Table 2). Long-haired hamsters increased the total duration of hoarding trips during shared conditions (Fig. 3 ; Table 2), storage size was larger during this condition (Fig. 3 ; Table 2).
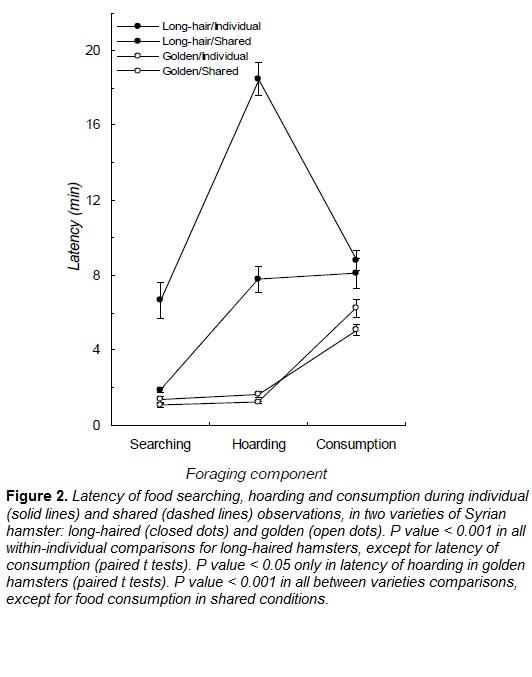
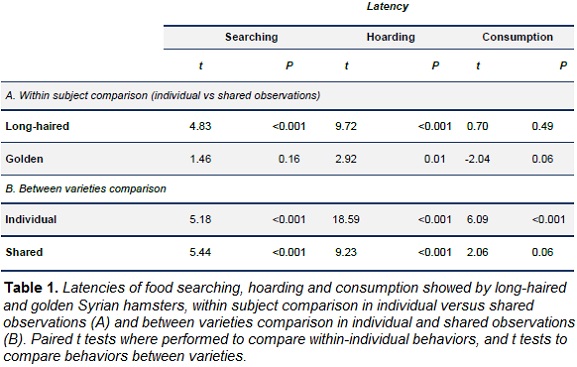
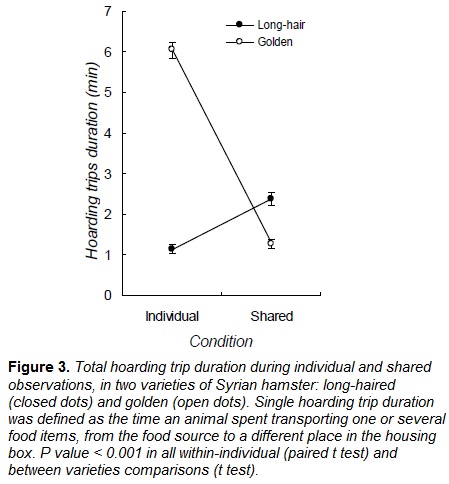
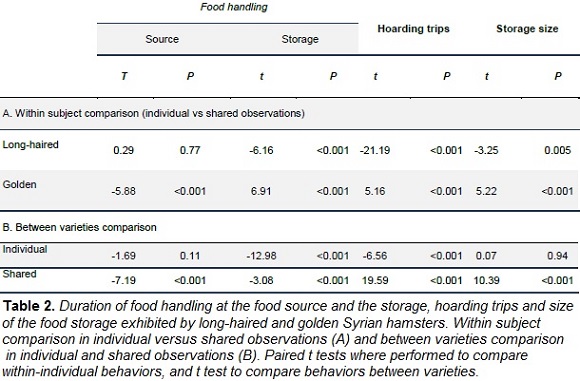
During shared conditions, golden Syrian hamsters did not change the latency of food searching or consumption, however, latency of hoarding was lower when a con-specific was present (Fig. 2, Table 1). In the presence of a con-specific, food handling in the food source was higher (individual mean = 4.42 ± 0.41, shared mean = 8.74 ± 0.57; Table 2) and lower in the storage (individual mean = 7.77 ± 0.34, shared mean = 4.77 ± 0.26, Table 2). The total duration of hoarding trips was lower in presence of a con-specific (Fig. 3; Table 2), and similarly happened with food storage size (Fig. 3; Table 2).
3.2 Between varieties comparison
During individual conditions, the latency of food searching, hoarding and eating was higher in long-haired hamsters than in golden ones (Fig. 2, Table 1). Complementarily, long-haired hamsters spent less time in hoarding trips than golden subjects (Fig. 3; Table 2) and handled the food items in the storage during shorter periods (Table 2). Food handling in the food source did not statistically differ between the two varieties of hamster (Table 2)
When a con-specific was present, long-haired hamsters took longer to start food searching and hoarding than subjects from the golden variety (Fig. 2, Table 1). Time spent in hoarding trips by long-haired hamsters was higher than time allocated by golden subjects (Fig. 3; Table 2), yet time spent handling food items was shorter for long-haired hamsters (Table 2). Consistently, storage of long-haired hamsters was larger compared to the deposits of the golden variety (Table 2).
4. DISCUSSION
The presence of a con-specific had an impact on the temporal organization of the food hoarding strategy in both long-haired and golden Syrian hamsters. The latency of food searching, hoarding and consumption were reduced in both varieties when exposed to a con-specific. Additionally, only in presence of a con-specific, long-haired hamsters hoarded food items before starting food consumption, whereas golden hamsters maintained the same sequence in individual and shared conditions (i.e. searching, hoarding and consumption). Although suggestive, these findings would need to be taken in perspective within a study using a larger sample size, more observational trials, but mainly wider housing facilities to allow individuals to perform more distant hoarding trips and build separate scatter hoards. The use of wider housing facilities could modify the effects found here by reducing stress associated with the presence of a con-specific, but might also better represent the characteristics of the environment where this behavior has evolved.
Long-haired and golden hamsters may resort to exhaustive exploitation of the food source while competing against a con-specific. The reduction of the latencies in food searching, hoarding and consumption when a con-specific was present suggests that, regardless of the artificial selection for highly sociable individuals in long-haired hamsters, both varieties respond to the presence of a con-specific as a potential hazard for the food source availability. In this context, low searching latencies confer a competitive advantage, because the first individual to detect a food source can exhaustively exploit it, transporting food items to one or multiple storages for posterior consumption. In presence of a con-specific, golden hamsters reduced the duration of hoarding trips, while increasing handling time in the food source, suggesting the exploitation of the food source by transporting several food items in fewer trips. In turn, during shared conditions, long-haired hamsters changed the strategy observed in individual conditions, i.e. direct item consumption in the food source, to a hoarding strategy, increasing thereby duration of the hoarding trips. Therefore, food hoarding may increase the competitive advantage of individuals when exploiting a food source susceptible to monopolization, as has been suggested for other rodent hoarder species where individuals additionally increase larder hoarding in the presence of other subjects (Zhang et al. 2014).
Typical social behavior of this species, in natural conditions, may explain changes in the food hoarding strategy in the presence of a con-specific. Syrian hamsters are solitary and do not exhibit gregarious behavior while foraging (Lupfer, Frieman & Coonfield, 2003 ; Gismonoli, 1999). Consequently, when competing for food in natural conditions, Syrian hamsters probably exhibit an exploitation strategy, rather than interfering with the contender's source use. In that case, is likely intensively once an individual finds a food source, such source is intensively exploited and the next forager might not be able to take advantage of it (Begon, Harper & Townsend, 2006). In a laboratory arrangement of classical conditioning, when a con-specific (conditioned stimulus) was presented just before the release of a ephemeral food source (unconditioned stimulus), opposite to what was found in rats, Syrian hamsters did not show any conditional response (i.e., approaching response towards the conditioned stimulus before the release of food items; Lupfer, Frieman & Coonfield, 2003). Interestingly, rats increase food-carrying behavior in the presence of a signal preceding a risky stimulus (Onuki & Makino, 2005). As a whole, the results of this study support the idea that, for Syrian hamsters, the presence of a con-specific may be a clue for food sources depletion. Hence, hoarding food items, from a recently found food source, might be a good strategy to avoid source depletion by a con-specific.
Social context affects food hoarding strategies in Syrian hamsters giving priority to food hoarding over item consumption. It seems likely that intra-specific competence has been one of the pressures favoring food hoarding behavior in hamsters. Further studies are necessary to evaluate whether hoarding food, in the presence of a con-specific, do provide an adaptive advantage to individuals after considering the potential cost-benefit balance, and to explore the role of cache pilferage on the spatial-temporal organization of food hoarding strategies.
5. REFERENCES
Alexander, L., Duthie, C., Fyfe, J., Haws, Z., Hunt, S., Montoya, I., Ochoa, C., Siva, A., Stringer, J. & Burns, K. (2005). An experimental evaluation of food hoarding by North Island robins (Petroica australis longipes). Nortonis, 52, 138-142. [ Links ]
Beck, M. J. & Vander Wall, S. B. (2010). Seed dispersal by scatter-hoarding rodents in arid environments. Journal of Ecology, 98, 1300-1309. [ Links ]
Begon, M., Harper, J. & Townsend, C. (2006). Ecology: From individuals to ecosystems. Malden, MA: Blackwell. [ Links ]
Briggs, J. & Vander Wall, S. (2004). Substrate type affects caching and pilferage of pine seeds by chipmunks. Behavioral Ecology, 15, 666-672. [ Links ]
Brodin, A. (2005). Mechanisms of cache retrieval in long-term hoarding birds. Journal of Ethology, 23, 77-83. [ Links ]
Dennis, A. (2003). Scatter-hoarding by musky rat-kangaroos, Hypsiprymnodon moschatus, a tropical rain-forest marsupial from Australia: implications for seed dispersal. Journal of Tropical Ecology, 19, 619-627. [ Links ]
Forget, P. M. & Vander Wall, S. (2001). Scatter-hoarding rodents and marsupials: convergent evolution on diverging continents. Trends in Ecology & Evolution, 16, 65-67. [ Links ]
Gerhardt, F. (2005). Food pilfering in larder-hoarding red squirrels (Tamiasciurus hudsonicus). Journal of Mammalogy, 86, 108-114. [ Links ]
Gismonoli, E. (1999). El Manual del Hámster. Barcelona: DeVecchi. [ Links ]
Holldobler, B. & Wilson, E.O. (1990). The ants. Cambridge, MA: Belknap/Harvard. [ Links ]
Jansen, P. A., & Forget, P. M. (2001). Scatterhoarding rodents and tree regeneration. Nouragues, 80, 275-288. Springer Netherlands [ Links ]
Jenkins, S., Rothstein, A. & Green, W. (1995). Food hoarding by Merriam's kangaroo rats: a test of alternative hypotheses. Ecology, 76, 2478-2481. [ Links ]
Lahti, K., Koivula, K., Rytkonen, S., Mustonen, T., Welling, P., Pravosudov, V. & OrcD, M. (1998). Social influences on food caching in willow tits: a field experiment. Behavioral Ecology, 9, 122-129. [ Links ]
Lupfer, W., Frieman, J. & Coonfield, D. (2003). Social transmission of flavor preferences in two species of hamsters (Mesocricetus auratus and Phodopus campbelli). Journal of Comparative Psychology, 117, 449-455. [ Links ]
Male, L. & Smulders, T. (2007). Hyperdispersed cache distributions reduce pilferage: a field study. Animal Behaviour, 73, 717-726. [ Links ]
Onuki, Y. & Makino, J. (2005). Food-carrying behavior increased under risk-approaching signal in rats (Rattus norvegicus). Physiology and Behavior, 84, 141-145. [ Links ]
Rinderer, T. E., Bolten, A. B. Harbo, J. R., & Collins, A. M. (1982). Hoarding behavior of European and Africanized Honeybees (Hymenoptera: Apidae). Journal of Economic Enthomology, 75, 714-715. [ Links ]
Shettleworth, S. J. (2001). Spatial behavior, food storing and the modular mind. In M. Bekoff, C. Allen & G. Burhardt (Eds.) The Cognitive Animal. USA: MIT Press. [ Links ]
Smith, C. & Reichman, O. (1984). Evolution of food caching by birds and mammals. Annual Review of Ecology and Systematics, 15, 329-351. [ Links ]
Steele, M., Turner, G., Smallwood, P., Wolf, J. & Radillo, J. (2001). Cache management by small mammals: Experimental evidence for the significance of acorn-embryo excision. Journal of Mammalogy, 82, 32-42. [ Links ]
Van Horik, J. & Burns, K. (2007). Cache spacing patterns and reciprocal cache theft in New Zealand robins. Animal Behaviour, 73, 1043-1049. [ Links ]
Vander Wall, S. & Jenkins, S. (1987). Reciprocal pilferage and the evolution of food-hoarding behavior. Behavioral Ecology, 14, 656-667. [ Links ]
Vander Wall, S. (1990). Food Hoarding in Animals. Chicago: Chicago University Press. [ Links ]
Yoshizawa, Y., Wada, K., Shiomi, G., Kameyama, Y., Wakabayashi, Y., Fukuta, K., & Hashizume, R. (2015). A 1-bp deletion in Fgf5 causes male-dominant long hair in the Syrian hamster. Mammalian Genome, 26 (11-12), 630-637. [ Links ]
Yasuda, M., Miura, S. & Nor Azman, H. (2000). Evidence for food hoarding behaviour in terrestrial rodents in Pasoh Forest Reserve, a Malaysian lowland rain forest. Journal of Tropical Forest Science, 12, 164-173. [ Links ]
Zhang, H., Gao, H., Yang, Z., Wang, Z., Luo, Y., & Zhang, Z. (2014). Effects of interspecific competition on food hoarding and pilferage in two sympatric rodents. Behaviour, 151(11), 1579-1596. [ Links ]
6. ACKNOWLEDGEMENT
This research complies with Colombian laws and Animal Behavior Guidelines for animal experimentation. Authors thank Oscar Córdoba, Jonathan Buriticá, Nataly Yañez and Lina Velandia for help with data collection, anonymous referees for valuable suggestions on a previous version of the manuscript, and María Luisa Ramírez for a thorough edition and suggestions on the manuscript.

