










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Ciencias Hortícolas
versão impressa ISSN 2011-2173
rev.colomb.cienc.hortic. vol.10 no.1 Bogotá jan./jun. 2016
https://doi.org/10.17584/rcch.2016v10i1.4457
Doi: http://dx.doi.org/10.17584/rcch.2016v10i1.4457
Índices de crecimiento en plantas de mora (Rubus alpinus Macfad) bajo diferentes sistemas de poda
Growth rates in blackberry (Rubus alpinusMacfad) plants under different pruning systems
BRIGITTE LILIANA MORENO-MEDINA1, FÁNOR CASIERRA-POSADA2, 4, MICHAEL BLANKE3
2 Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia).
3 INRES-Horticultural Science, University of Bonn, Bonn (Alemania).
4 Autor para correspondencia: fanor.casierra@uptc.edu.co
Fecha de recepción: 08-02-2016 Aprobado para publicación: 14-05-2016
RESUMEN
El cultivo de mora en Colombia es un renglón importante para el desarrollo económico de medianos y pequeños productores. Las actividades de poda son labores indispensables en este cultivo y tienen el propósito de renovar las ramas productivas y mejorar la relación entre la oferta y demanda de fotoasimilados. En el presente ensayo se utilizaron plantas de dos años de edad, tutoradas en T; se marcaron 48 tallos bajo diferentes sistemas de poda (testigo: sin podar; poda corta: tallo a 1,6 m de longitud; poda larga: tallo a 2,6 m de longitud), se midió la longitud de los tallos, y se registró el número de hojas y de panículas aparecidas a lo largo del tiempo, se graficó el comportamiento de las variables respecto a los días después de brotación (ddb). Se establecieron las curvas y las tasas de crecimiento mediante análisis funcional. Como resultado se obtuvo que los modelos logísticos fueron de mejor ajuste para la descripción del crecimiento de esta especie, la máxima longitud del tallo se presentó a los 231 ddb con 535 cm, la tasa relativa de crecimiento (TRC) del tallo disminuyó gradualmente, la tasa absoluta de crecimiento (TAC) del tallo así como el número de hojas y panículas disminuyeron con el tiempo. El número máximo de hojas fue de 55 a los 231 ddb; la floración en todos los sistemas de poda inició a los 126 ddb aproximadamente; el número de inflorescencias en promedio fue de 18 en plantas sin poda, mientras que con poda corta tuvo un valor de 10, y de 21 con poda larga. Los resultados mostraron que la poda es un factor determinante en el desarrollo fisiológico y productivo en esta especie de mora.
Palabras clave adicionales: curva de crecimiento, tasa de crecimiento, panículas, modelo logístico, brotación.
ABSTRACT
In Colombia, the blackberry is an important perennial crop for the economic sustainable development of medium and smallholder farmers. Pruning is essential for this crop and aims to renew the productive, fruit bearing branches and normalize the sink-source relationships of photoassimilates. The establishment of physiological indices and growth curves for blackberry crops with different pruning practices will develop proposals for the agricultural handling of this species. In this study, two-year-old blackberry crops were used with a T-trellis; 48 canes were grouped into three pruning treatments; a) control: without pruning; b) short pruning: canes with a 1.6 m length; and c) long pruning; canes with a 2.6 m length. The length of the canes, and the number of leaves and panicles were registered over time, in terms of days after sprouting. The curves and rates of growth were established with a functional analysis. The logistic model was ideal for expressing the growth of the plants. The maximum length of the canes was seen at 231 days after sprouting: 535 cm. The relative growth rate of the canes decreased gradually over time. The values of the absolute growth rate of the canes as well as the number of leaves and panicles decreased over time too. The maximum number of leaves was seen from 55 to 231 days after sprouting; the flowering in the plants in the pruned treatments started at 126 days after sprouting. The average number of inflorescences was 18 in the shrubs without pruning, 10 with the short pruning and 21 with the long pruning. The results showed that pruning is a determinant factor in the physiological and productive development of blackberry shrubs and that the long pruning, with a 2.6 m length, was the best pruning alternative.
Additional keywords: Growth curve, growth rate, panicles, logistic model, sprouting.
INTRODUCCIÓN
La mora se considera un fruto delicatessen por su característico sabor acidulado, cualidades nutricionales, antioxidantes y gastronómicas, entre otras (Farinango, 2010; Souza et al., 2014). Colombia presenta alto potencial en el área de producción de este tipo frutales, y en este sentido, el género Rubus es importante para generar desarrollo económico para medianos y pequeños productores (MADR, 2014). Adicionalmente, se reportan algunas especies de mora silvestres que se pueden encontrar a lo largo de las cercas, caminos, fincas, bosques marginales y en los mismos cultivos comerciales (Cancino et al., 2011) y son consideradas en muchas regiones del país como sustento económico para los pobladores.
En el departamento de Boyacá los municipios que reportan mayor producción de mora son: Saboyá con 842 t año-1, Gachantivá con 735 t año-1 y Arcabuco con 562 t año-1, evidenciando que la provincia del Alto Ricarte muestra gran diversidad y potencial para este cultivo (MADR, 2014).
Es importante resaltar que en las zonas productoras de mora existe alta dependencia por la variedad Castilla (R. glaucus), sin embargo, esto limita la producción teniendo en cuenta la susceptibilidad de esta especie a problemas fitosanitarios, la baja calidad en parámetros poscosecha, deficientes prácticas de manejo agronómico, entre otros (Franco y Giraldo, 2000). En plantas de R. glaucus, García y García (2001) reportan que la primera cosecha inicia entre los 10 y 12 meses después del trasplante y luego se realizan cosechas semanales en forma continua con algunas épocas de concentración de la producción. Además, debido a su hábito de crecimiento y tipo de ramas, esta especie exige constantes labores de poda y tutorado (Mejía, 2011).
Casierra-Posada et al. (2012) mencionan que los objetivos de las actividades de poda están orientados a formar plantas para facilitar actividades agronómicas, mejorar la entrada de luz, regular los periodos de producción, mejorar la calidad poscosecha de los frutos, establecer el equilibrio fisiológico entre el crecimiento vegetativo y generativo, disminuir el número de ramas enfermas o no deseables, y dejar un número adecuado de frutos en cada rama, con el fin de evitar fracturas por exceso de peso. Las plantas de R. alpinusson arbustos semierectos y poseen ramas de seis metros o más de longitud en las cuales pueden distribuir cerca de 20 panículas. Esto pone de manifiesto que las actividades de poda son labores indispensables en este cultivo.
Las curvas de crecimiento en los vegetales permiten conocer la forma como evoluciona y se comporta una planta en función del tiempo y su entorno, por tal razón, la obtención de este tipo de información es necesaria para el diseño racional de estrategias de manejo agronómico en los cultivos, con el fin de garantizar respuestas que satisfagan las necesidades del productor y cumplan con las exigencias del consumidor final (Casierra-Posada et al., 2004). Graber (1997) manifiesta que los estudios fenológicos permiten entender en forma clara el comportamiento de un vegetal en relación con el tiempo, y a partir de ello obtener mayor conocimiento sobre el crecimiento y desarrollo de las plantas y sus diferentes etapas. En los vegetales, los procesos de crecimiento y desarrollo son eventos irreversibles, se evidencian cambios en tamaño, número, masa y forma. Estos parámetros y sus respectivas modificaciones son el resultado de la interacción del genotipo y el ambiente. Esto tiene como consecuencia el aumento cuantitativo en tamaño y peso en los vegetales, debido principalmente a la división y elongación celular, facilitadas por los procesos fotosintéticos, respiratorios y por la dinámica que genera la planta con la absorción, transporte y transpiración de agua y nutrientes (Taiz y Zeiger, 2010; Tadeu et al., 2015; Hussaina et al., 2016).
Pese a las dificultades de manejo técnico de R. alpinus(Gómez et al., 2010; Moreno-Medina y Deaquiz, 2016), en Arcabuco y la provincia del Alto Ricaurte en Boyacá, la producción y explotación de esta especie como cultivo se ha venido incrementando en términos de área sembrada. Sin embargo, no se conoce manejo agronómico que permita efectuar un direccionamiento adecuado para este sistema de producción. Por tal razón, mediante este trabajo se pretendió establecer índices fisiológicos de las plantas de mora (R. alpinus) bajo diferentes sistemas de poda, lo cual permitirá construir las curvas de crecimiento (tallos, hojas y panículas) y plantear una propuesta que mejore el manejo agronómico de esta especie en la zona.
MATERIALES Y MÉTODOS
Ubicación y material vegetal
Se utilizaron plantas de mora (R. alpinus) de dos años de edad, establecidas en Arcabuco (Boyacá), ubicadas a 5°4413.05 N y 73°2914.33 a una altitud de 2.495 msnm. El cultivo se estableció con una densidad de 1.100 plantas/ha, con una distancia entre surcos y plantas de 3 m, tutoradas con sistema de conducción en T. Se realizaron labores agronómicas adecuadas como fertilización, manejo de plagas, enfermedades y malezas.
Prácticas de poda y seguimiento del ensayo
En las plantas de mora se marcaron 16 tallos por cada tratamiento, a los anteriores se les dejó libre crecimiento y una vez alcanzaron la longitud requerida en cada uno de los tratamientos se efectuó la poda (testigo: sin podar; poda corta: tallo a 1,6 m de longitud; poda larga: tallo a 2,6 m de longitud), se registró la longitud de los tallos, tomando la medida desde la base del tallo hasta la yema terminal, a la par se contó el número de hojas y número de panículas a través del tiempo. Esta información se registró semanalmente entre los 14 y 231 días después de brotación (ddb). Las mediciones del tallo iniciaron una vez el brote se hizo visible. Las hojas se contaron una vez se inició la apertura de los foliolos y el número de panículas se contó una vez iniciada su formación.
Con la información obtenida, se graficó el comportamiento de cada una de las variables respecto al tiempo (ddb) y se establecieron los modelos de análisis funcional. Las curvas y tasas de crecimiento (expresadas en número de estructuras e incremento de la longitud) se ajustaron al modelo no lineal de tipo logístico, usando como parámetro de elección el ajuste o coeficiente de regresión (R2) más cercano a la unidad (Carranza et al., 2009). La información fue procesada con la aplicación SAS versión 8.1.
RESULTADOS Y DISCUSIÓN
Modelos de regresión
En la tabla 1 se muestran las ecuaciones de regresión obtenidas en la presente investigación, donde se puede estimar la longitud de los tallos, el número de hojas y el número de panículas en esta especie de mora, en respuesta a los diferentes sistemas de poda implementados respecto a los ddb. Según Barceló et al. (1987) el crecimiento de las plantas se puede cuantificar a través de la construcción de ecuaciones de regresión, y a la par se puede evaluar el comportamiento de una variedad bajo determinadas condiciones.
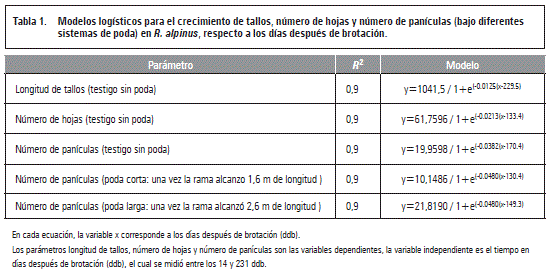
Crecimiento de tallos
La longitud del tallo en R. alpinussin podar presentó un crecimiento sigmoide simple generando un incremento progresivo a través del tiempo (ajustado al modelo logístico), el tallo alcanzó su longitud máxima (535 cm) a los 231 ddb en promedio (figura 1). Sin embargo, la ralentización en el crecimiento de los tallos entre los 100 y 150 ddb, posiblemente se debió a que al mismo tiempo inició la floración y se presentó la máxima tasa de crecimiento en hojas, es decir que este descenso parcial en la velocidad del crecimiento estuvo posiblemente afectado por la redistribución de los fotoasimilados en la planta. Lo anterior se puede corroborar con lo mencionado por Monasterio (1995), quien describe que, en presencia de estructuras foliares, el flujo de carbohidratos se dirige hacia las zonas de crecimiento, no obstante en el caso del género Rubus, los fotoasimilados tienen como vertedero principal las raíces. Por su parte, Parra-Quezada et al. (2007) indican que el sistema radicular en el cultivar Malling autum Blissalmacena carbohidratos y nutrientes para rebrotar después de las labores de poda y despuntes.
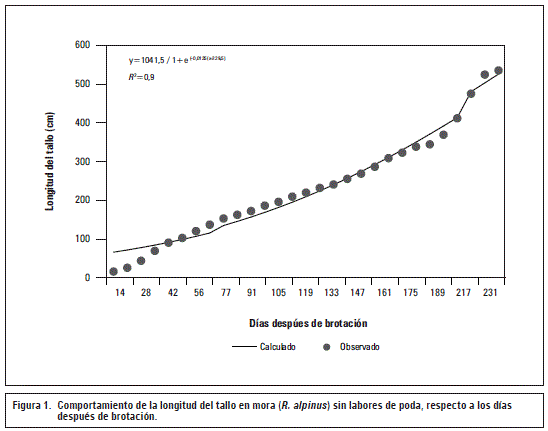
La tasa absoluta de crecimiento (TAC) en los tallos de la especie R. alpinusaumentó lentamente antes de los 50 ddb, seguidamente aumentó hasta alcanzar su punto máximo de crecimiento a los 217 ddb con 3,23 cm/semana, luego de esta etapa la TAC no continuo su ascenso, evidenciando que la máxima TAC se exhibió en la segunda etapa del periodo evaluado, según el modelo logístico (figura 2A).
Con relación a la tasa relativa de crecimiento (TRC), se observó que disminuyó de manera constante, sin embargo, hasta los 119 ddb la disminución fue lenta (0,0098 cm cm-1 por semana) y, a continuación, la TRC disminuyó notablemente (figura 2B). Lo anterior coincidió con el inicio en la formación de estructuras reproductivas en el presente ensayo, que tuvo lugar alrededor de los 126 ddb.
La TAC y TRC se muestran como indicadores de la relación fuente vertedero (Hunt, 1982), lo cual es importante para establecer actividades agronómicas tendientes a incrementar la cantidad de biomasa o estructura vegetal en un periodo determinado, con el fin de mejorar los procesos productivos en cultivos de R. alpinus.
Estos resultados confirman lo mencionado por Hunt (1982) y Villar et al. (2004), quienes describen que el estudio del crecimiento de las plantas y su posterior análisis permite relacionar la ecofisiología y la agronomía con el fin de mejorar las actividades de manejo en los cultivos.
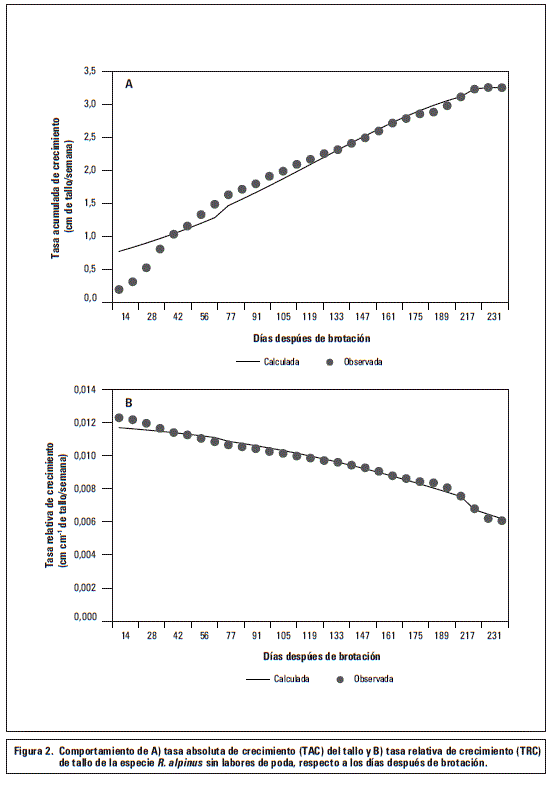
Número de hojas
En las plantas de R. alpinusel número de hojas emitidas aumentó a través del tiempo, produciéndose, en promedio, 55 hojas en los tallos sin podar (figura 3). En los tallos que se podaron a 1,6 m de longitud se desarrollaron en promedio 19 hojas y en los tallos podados a 2,6 m de longitud se encontraron hasta 36 hojas para el periodo comprendido entre los 14 y 231 ddb.
La aparición de hojas en plantas de R. alpinussin podar presentó un crecimiento sigmoide simple evidenciando un incremento progresivo a través del tiempo, ajustado al modelo logístico y alcanzó el número máximo de hojas (55) a los 231 ddb. Estos resultados ponen en evidencia el potencial productivo de la especie, dado que con un alto número de hojas se pueden mejorar los aspectos productivos en el cultivo, teniendo en cuenta que la actividad fotosintética foliar y el crecimiento en los vegetales está íntimamente relacionado. Al respecto se ha demostrado en cultivos de tomate que el proceso fotosintético es el responsable de la producción de fotoasimilados y con ello del incremento en el rendimiento (Barraza et al., 2004).
La tasa absoluta de crecimiento (TAC) para el número de hojas en R. alpinusaumentó paulatinamente antes de los 160 ddb, es aquí justamente donde la aparición de hojas tiene su crecimiento máximo (0,3 hojas/semana), luego, disminuyó drásticamente hasta alcanzar su menor punto de crecimiento a los 231 ddb con un valor de 0,1 hojas/semana (figura 4). Con lo anterior y al igual que en el crecimiento de los tallos, se evidenció que la máxima tasa de crecimiento se exhibió en la segunda etapa del periodo evaluado según el modelo logístico. A pesar de que la TAC en hojas disminuyó con el tiempo, la aparición de estas es continua, como se presentó en la figura 3, si los meristemos apicales permanecen intactos, es decir, si no se efectúan labores de poda o se evidencian daños por agentes bióticos o abióticos.
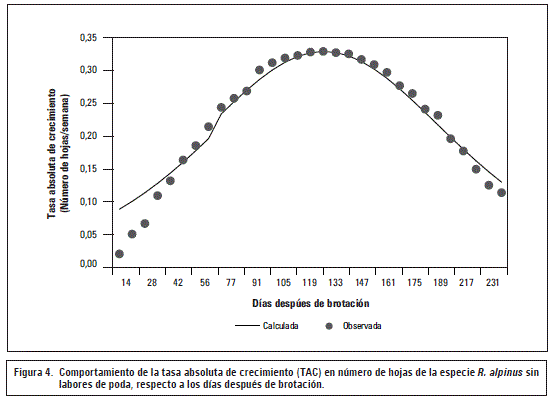
Al igual que en el crecimiento del tallo, la aparición de hojas también puede verse afectada por la formación de estructuras reproductivas, sin embargo, es importante aclarar que la disminución en la TAC puede verse influenciada por la relación fuente-vertedero, teniendo en cuenta que para este caso las hojas que se encuentran en formación y desarrollo actúan como órganos vertedero y causan competencia por fotoasimilados (Casierra-Posada et al., 2004).
Estos resultados son importantes debido a que en la mayoría de las plantas las hojas son los órganos directamente implicados en el proceso fotosintético, o el lugar donde se efectúa la captura de la energía luminosa necesaria para impulsar las diferentes reacciones químicas vitales para el crecimiento y desarrollo de los vegetales (Taiz y Zeiger, 2010).
En el caso de la especie R. ulmifolius, existen reportes que describen la producción de flores y frutos hasta por cuatro años en un mismo tallo (Salter, 1845), con lo anterior se puede mencionar que la especie R. alpinustiene excelentes posibilidades frente a una producción permanente, además Casierra-Posada et al. (2007) mencionan que el mayor tamaño y cantidad de sólidos solubles totales (°Brix) en los frutos de duraznero (Prunus persicaL. Batsch, cv. Rubidoux) se puede atribuir a la relación entre el número de hojas y el fruto, es decir que de manera análoga, podría plantearse que en R. alpinus, con la continua formación de hojas se incrementa y mejora la calidad de las bayas que se encuentran en cada tallo.
Número de panículas
El número de panículas en plantas de R. alpinuspresentó una tendencia sigmoide simple, ajustado a un modelo logístico, e independientemente del tipo de poda, evidenció un incremento progresivo a través del tiempo. Los tallos sin podar alcanzaron el número máximo de 18 panículas a los 231 ddb, los tallos que se podaron a 1,6 m de longitud presentaron su máximo número de inflorescencias a los 224 ddb con 10 panículas y los tallos podados a 2,6 m de longitud tenían 21 panículas como crecimiento máximo a los 217 ddb.
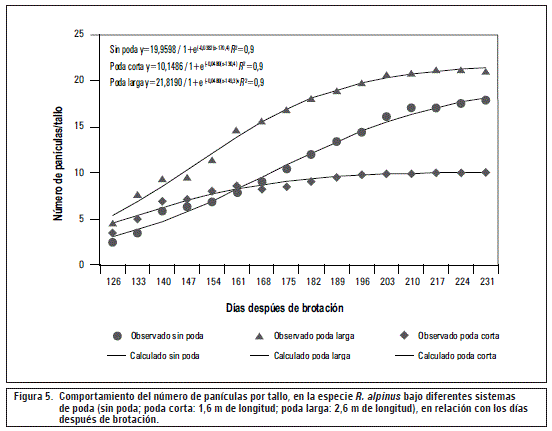
El número de panículas producidas aumentó con el tiempo, hasta completar 21 en promedio para el caso de los tallos que se podaron a 2,6 m de longitud. Los tallos que no se podaron desarrollaron en promedio 18 panículas, y en aquellos podados a 1,6 m de longitud se reportan 10 panículas. Es importante resaltar que las podas realizadas no tuvieron efecto significativo sobre el adelanto en la formación de panículas, ya que en los sistemas de poda evaluados la formación de este tipo de inflorescencias se originó a partir de los 126 ddbaproximadamente, es decir cerca de los 4,5 meses después de la emergencia del tallo, lo que muestra el posible tiempo o intervalos en los cuales se genera la producción. No obstante, Giraldo y Franco (2002) reportan para R. glaucusintervalos de producción cada 5-6 meses, es así como R. alpinus expresa posibles adelantos en el periodo de cosecha en relación con R. glaucus, la variedad de mora más cultivada en Colombia. Con lo anterior se evidencia un potencial para el establecimiento de la especie R. alpinuscomo cultivo.
Con base en estos resultados se puede mencionar que las actividades de poda para este cultivo se deben realizar con propósitos fitosanitarios y para retirar el exceso de ramas, hojas y estructuras que ya produjeron; inclusive, se debe tener en cuenta que con las labores de poda, se disminuye la competencia por fotoasimilados y con ello aumenta la calidad y adecuada formación de los frutos en desarrollo, además se puede establecer como estrategia de manejo agronómico, realizar podas sin dejar que las ramas sobrepasen los 3 m de longitud, lo anterior teniendo en cuenta que cerca del 70% de la producción (entre 18 y 20 panículas) se forman en esta porción de tallo.
La TAC en número de panículas en R. alpinusaumentó paulatinamente hasta los 170 ddb para todas las plantas independientemente de la poda (figura 6). En los tallos sin podar la máxima TAC se dio a los 168 ddb con 5,01 panículas/semana, mientras que en los tallos podados a 1,6 m de longitud se evidenció una TAC de 0,83 panículas/ semana y en los tallos con poda a los 2,6 m de longitud se alcanzó la máxima TCA a los 161 ddb con 5,94 panículas/semana. La TAC en número de panículas disminuyó gradualmente, al parecer debido a la formación continua de este tipo de estructuras, además, los fotoasimilados deben ser redistribuidos en las panículas antes formadas pero que se encuentran en continuo crecimiento hasta llegar a su maduración.
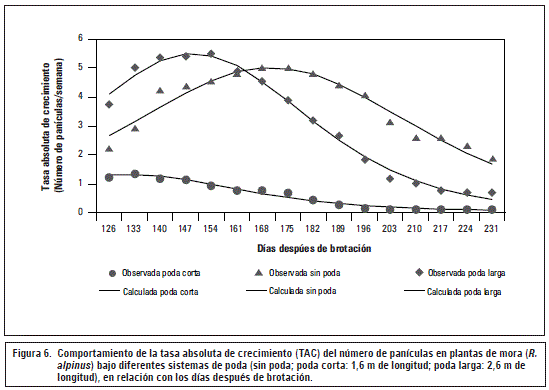
Tomando como referencia el trabajo de Aliquóet al. (2010), se describen para el cultivo de vid diferentes tipos de poda teniendo en cuenta la longitud del tallo al momento del corte (poda corta y poda larga) y se menciona que esta longitud se puede relacionar con el número de yemas que dan origen a los racimos y con ello a la obtención de cosechas de calidad. A la par, Reynier (1989) menciona que la poda corta implica menor número de yemas, menor producción y posiblemente mayor vigor en el vegetal, sin embargo, con la poda larga se incrementaría el número de yemas, racimos y área foliar, no obstante, se disminuye la calidad de la fruta por la maduración desigual con la respectiva reducción de color y del contenido de sólidos solubles totales, además de ocasionar una baja lignificación en la estructura principal de la planta.
El estudio del número de panículas muestra alternativas para la recomendación técnica en la implementación de podas para el cultivo de R. alpinus. Con base en esta información se puede proponer que las actividades de poda se realicen a 2,6 m de longitud con el objetivo de aumentar el número de inflorescencias en dicho cultivo, sin dejar de lado una fertilización adecuada, la cual permitirá la formación de estructuras propicias para el llenado de frutos y el aumento de su calidad.
CONCLUSIONES
Una vez se construyeron los modelos logísticos para la caracterización de los parámetros de crecimiento en R. alpinus, se puede afirmar que las plantas mostraron disminución en las tasa de crecimiento con el paso del tiempo para los parámetros de longitud del tallo, número de hojas y panículas, no obstante, las labores de poda no tuvieron efecto sobre el adelanto de la floración, aun cuando se encontraron algunas diferencias en el número de inflorescencias formadas por cada tallo. Los resultados obtenidos permitieron elegir la poda larga, realizada a 2,6 m de longitud, como una alternativa que, acompañada de otras labores agronómicas adecuadas, permitirá expresar el potencial productivo de esta especie en la zona en estudio.
REFERENCIAS BIBLIOGRÁFICAS
Aliquó, G., A. Catana y G. Aguado. 2010. La poda de la vid. Secretaría de Agricultura, Ganadería, Pesca y Alimentación; Instituto Nacional de Tecnología Agropecuaria (INTA), Buenos Aires. [ Links ]
Barceló, C.J., G. Nicolás, B. Sabater y R. Sánchez. 1987. Fisiología vegetal. Ediciones Pirámide, Madrid. pp. 429-441. [ Links ]
Barraza, F., G. Fischer y C. Cardona. 2004. Estudio del proceso de crecimiento del cultivo del tomate (Lycopersicon esculentum Mill.) en el Valle del Sinú medio, Colombia. Agron. Colomb. 22(1), 81-90. [ Links ]
Cancino, O., L. Sanchez, E. Quevedo y C. Díaz. 2011. Caracterización fenotípica de accesiones de especies de Rubus L. de los municipios de Pamplona y Chitagá, región Nororiental de Colombia. Uni. Sci. 16(3), 219-233. Doi: 10.11144/javeriana.SC16-3.pcor [ Links ]
Carranza, C., O. Lanchero, D. Miranda y B. Chaves. 2009. Análisis del crecimiento de lechuga (Lactuca sativa L.) Batavia cultivada en un suelo salino de la Sabana de Bogotá. Agron. Colomb. 27(1), 41-48. [ Links ]
Casierra-Posada, F., F. Barreto y O. Fonseca. 2004. Crecimiento de frutos y ramas de duraznero (Prunus pérsica L. Batsch, cv. Conservero) en los altiplanos colombianos. Agron. Colomb. 22(1): 40-45. [ Links ]
Casierra-Posada, F. y G. Fischer. 2012. Poda de árboles frutales. pp. 169-185. En: Fischer, G. (ed.). Manual para el cultivo de frutales en el trópico. Produmedios, Bogotá [ Links ].
Casierra-Posada, F., J. Rodríguez y J. Cárdenas-Hernández. 2007. La relación hoja:fruto afecta la producción, el crecimiento y la calidad del fruto en duraznero (Prunus persicaL. Batsch, cv. Rubidoux). Rev. Fac. Nal. Agr. Medellín 60(1), 3657-3669. [ Links ]
Farinango, M. 2010. Estudio de la fisiología postcosecha de la mora de castilla (Rubus glaucusBent) y mora variedad brazos (Rubus sp). Trabajo de grado. Escuela Politécnica Nacional, Facultad de Ingeniería Química y Agroindustria, Quito. [ Links ]
Franco, G. y M. Giraldo. 2000. El cultivo de la mora. 3ª ed. Instituto Colombiano Agropecuario (ICA), Manizales, Colombia. [ Links ]
García, M y H. García. 2001. Manejo cosecha y poscosecha de mora, lulo y tomate de árbol. Corpoica, Bogotá [ Links ].
Giraldo, M. y G. Franco. 2002. El cultivo de la mora. Corpoica-Pronatta, Manizales, Colombia. [ Links ]
Gómez, R. y H. Pinilla. 2010. Desarrollo en armonía ambiental: El caso de la vereda Peñas Blancas del municipio de Arcabuco, Boyacá. Cultura Científica 8, 65-74. [ Links ]
Graber, U. 1997. Fenología de los cultivos: mora de Castilla (Rubus glaucus B.). Granja Experimental, Píllaro, Ecuador. [ Links ]
Hunt, R. 1982. Plant growth curves. The functional approach to plant growth analysis. Edward Arnold, Londres, U.K. [ Links ]
Hussaina, I., S. Roberto, I. Batista, A. Marinho de Assis, R. Koyama y L.Correa. 2016. Phenology of Tupy and Xavante blackberries grown in a subtropical área. Sci. Hortic. 201, 78-83. Doi: 10.1016/j.scienta.2016.01.036 [ Links ]
Mejía, B. 2011. Caracterización morfoagronómica de genotipos de mora (Rubus glaucusBenth.) en la granja experimental. Informe técnico del proyecto de investigación. INIAP, Tumbaco, Ecuador. [ Links ]
MADR (Ministerio de Agricultura y Desarrollo Rural). 2014. Base de datos de la producción de frutas y hortalizas 2007-2014. En: http://www.agronet.gov.co/Paginas/estadisticas.aspx; consulta: noviembre de 2014. [ Links ]
Monasterio-Huelin, E. 1995. Biología de reproducción en Rubus L. (Rosaceae). Propagación vegetativa. Anales Jard. Bot. Madrid 52(2), 145-149. [ Links ]
Moreno, B.L. y Y.A. Deaquiz O. 2016. Caracterización de parámetros fisicoquímicos en frutos de mora (Rubus alpinusMacfad). Acta Agron. 65(2), 130-136. Doi: 10.15446/acag.v65n2.45587 [ Links ]
Parra-Quezada, R. Á., V. Guerrero-Prieto y J. ArreolaAvila. 2007. Efecto de fecha y tipo de poda en frambuesa roja Malling autumn Bliss. Rev. Chapingo Ser. Hort. 13(2), 201-206. [ Links ]
Reynier, A. 1989. Manual de viticultura. Cuarta edición. Ed. Mundi-Prensa, Madrid. [ Links ]
Salter, T.B. 1845. Observations on the genus Rubus. Phytologist 2, p. 107. [ Links ]
Souza, V. R., P. Pereira, T. Silva, L. Oliveira Lima, R. Pio y F. Queiroz. 2014. Determination of the bioactive compounds, antioxidant activity and chemical composition of Brazilian blackberry red raspberry, strawberry, blueberry and sweet cherry fruits. Food Chem. 156, 362-368. Doi: 10.1016/j.foodchem.2014.01.125 [ Links ]
Tadeu, M., F. Souza, R. de Pio, M. Valle, G. do Locatelli, G. Guimarães y B. Silva. 2015. Drastic summer pruning and production of blackberry cultivars in subtropical areas. Pesqui. Agropecu. Bras. 50, 132-140. Doi: 10.1590/S0100-204X2015000200005 [ Links ]
Taiz, L. y E. Zeiger. 2010. Plant physiology. 5a ed. Sinauer Associates, Sunderland, MA. [ Links ]
Villar, R., J. Ruiz. J. Robleto, L. Quero, P. Hendrik, F. Valladares y T. Marañón. 2004. Tasas de crecimiento en especies leñosas: aspectos funcionales e implicaciones ecológicas. Ecología del Bosque Mediterráneo en un Mundo Cambiante. Ministerio de Medio Ambiente, EGRAF S.A., Madrid. [ Links ]

