










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Hortícolas
Print version ISSN 2011-2173
rev.colomb.cienc.hortic. vol.10 no.2 Bogotá July/Dec. 2016
https://doi.org/10.17584/rcch.2016v10i2.4702
Doi: http://dx.doi.org/10.17584/rcch.2016v10i2.4702
1 Facultad de Ciencias Agropecuarias, Universidad de La Salle, Yopal-Casanare (Colombia).
2 Facultad de Ciencias, Departamento de Biología, Universidad Nacional de Colombia, Bogotá (Colombia).
3 Autor para correspondencia. jofernandez@unisalle.edu.co
Fecha de recepción: 31-05-2016. Aprobado para publicación: 24-11-2016.
RESUMEN
El éxito de las estrategias de expansión de la producción cacaotera en Colombia por cuenta de las nuevas oportunidades del mercado requiere entre otros, de producción de plantas de alta calidad. A pesar de que uno de los factores más determinantes para la calidad del material vegetal en vivero es la nutrición, la información existente no considera características genéticas y fisiológicas inherentes a las plantas ni el balance de nutrientes a partir de las condiciones locales para recomendaciones de fertilización en el país. Con el objetivo de analizar la dinámica nutricional de plantas de cacao (IMC67) cuando son sometidas a diferentes dosis de fertilizantes con N, P y K en etapa de vivero, se establecieron tres ensayos con dosis variables de N, P y K a partir de su relación porcentual, con una dosis de referencia. A los 124 días después de siembra, se determinó la biomasa seca y el contenido nutricional de N, P, K, Ca, Mg, Cu, Fe, Mn, Zn y B de las plantas y se estimaron parámetros de dinámica nutricional, de correlación entre nutrientes y de eficiencia agronómica. Los resultados permiten concluir consistentemente que la dinámica nutricional de N, P y K no solo cambió en relación a los tratamientos implementados, sino que afectó la dinámica de prácticamente todos los otros nutrientes evaluados, lo que se confirmó al analizar la eficiencia agronómica de los nutrientes aplicados, por cuanto las diferencias en las magnitudes de este parámetro correspondieron a desbalances nutricionales.
Palabras clave adicionales: absorción, extracción, concentración, eficiencia agronómica.
ABSTRACT
The success of the expansion strategies of cocoa production in Colombia due to the new opportunities of the market requires, among others, the production of high quality plants. Although one of the most determinant factors for the quality of plant material in nursery is nutrition, the existing information does not consider genetic and physiological characteristics inherent to plants or nutrient balance from local conditions used for fertilization recommendations in the country. In order to analyze the nutritional dynamics of cocoa plants (IMC67) when submitted to different doses of fertilizers with N, P and K in the nursery stage, three experiments with variable doses of N, P and K from the percentage relation were established, with a reference dose. At 124 days after sowing, the dry biomass and the N, P, K, Ca, Mg, Cu, Fe, Mn, Zn as well as B content of plants were determined and also parameters of nutritional dynamics, correlation between nutrients and agronomic efficiency. The results allow to conclude consistently that the nutritional dynamics of N, P and K not only changed in relation to the implemented treatments, but also affected the dynamics of almost all other nutrients evaluated, which was confirmed by analyzing the agronomic efficiency of the nutrients applied as the differences in the magnitudes of this parameters corresponded to the nutritional imbalances.
Additional key words: absorption, extraction, concentration, agronomic efficiency.
INTRODUCCIÓN
El cacao (Theobroma cacao L.) se cultiva en países del trópico ubicados entre los 20° latitud norte y 20° latitud sur (Clough et al., 2009). El precio del cacao ha venido presentado una tendencia al alza en tanto que se espera un crecimiento del consumo de cacao en el mundo (World Cocoa Foundation, 2014), lo que se traduce en nuevas oportunidades para el mercado de cacao para países como Colombia.
Colombia cuenta con alrededor de 165.000 ha sembradas de cacao pero tiene un potencial de 693.669 ha sin restricciones para su siembra (García et al., 2005). Recientemente, el Plan Decenal Cacaotero 2012- 2021, proyectó expandir la producción cacaotera con diversas estrategias dentro de las que se cuentan el establecimiento de 150.000 ha nuevas, la renovación de otras 40.000 ha y el incremento en la productividad de 400 a 1.200 kg ha-1 por año (MADR, 2012). Para el éxito de estas estrategias se requiere entre otros, la producción de plantas de alta calidad desde la etapa de vivero para ser utilizadas como patrones en las nuevas plantaciones.
A pesar de que uno de los factores más determinantes para la calidad del material vegetal en vivero es la nutrición (Hartmann et al., 2011), en el país son pocas las publicaciones científicas en torno a la nutrición de cacao en etapa de vivero. Al igual que otros países, en Colombia los servicios de extensión aún recomiendan el uso de fórmulas únicas que no toman en cuenta las variaciones en el suelo, las condiciones ambientales, ni el balance de nutrientes a partir de las condiciones locales (Snoeck et al., 2016). Más escasos son los reportes científicos que consideran las características genéticas y fisiológicas inherentes a las plantas y que determinan cómo absorben, traslocan o usan los nutrientes, lo cual puede ser un factor limitante, aunque la disponibilidad de nutrientes sea adecuada (Van Vliet et al., 2015).
En este contexto, la producción de material vegetal de propagación de calidad desde el punto de vista nutricional, implica la realización de estudios nutricionales en los patrones más utilizados en Colombia. Recientemente, Santiago et al. (2016) describieron el efecto de diferentes tratamientos de fertilización con N, P y K sobre algunos parámetros biométricos indicadores de calidad de material vegetal de un clon ampliamente utilizado en Colombia como patrón (IMC67), sin embargo no se suministraron datos acerca de su dinámica nutricional. Consecuentemente, el objetivo de esta investigación fue analizar la dinámica nutricional de plantas de cacao (IMC67) cuando son sometidas a diferentes dosis de fertilizantes con N, P y K en etapa de vivero.
MATERIALES Y MÉTODOS
Se utilizaron semillas de cacao (IMC67), procedentes de un jardín clonal ubicado en el municipio de Muzo (Boyacá, Colombia), las cuales fueron sembradas de forma individual en bolsas de 3 kg que contenían un sustrato homogéneo compuesto por suelo y arena en proporción 1:1. Para esto se utilizó un suelo inceptisol proveniente de una capa ubicada en los primeros 0 a 20 cm desde la superficie. De este sustrato homogéneo se tomó una muestra compuesta para análisis físicoquímico (Tab. 1). El experimento se desarrolló en condiciones de vivero por 124 d. Durante el crecimiento, las plantas estuvieron bajo polisombra a 12 h de luz natural al 60%, a una temperatura promedio de 25°C y a una humedad relativa promedio de 83%. No se presentaron problemas sanitarios importantes, así que su manejo fue estrictamente cultural. La metodología de establecimiento previamente descrita, se basó en experimentos similares de nutrición en cacao (Oyewole et al., 2012; Sodré et al., 2012; Oliveira et al., 2015).
Bajo un diseño completamente aleatorizado, se implementaron tres ensayos de fertilización para N, P y K. En cada ensayo se priorizó un nutriente de interés, en el primer ensayo se priorizó el N (ensayo A, Nx- P-K), en el segundo el P (ensayo B, N-Px-K) y en el tercero el K (ensayo C, N-P-Kx). En los tres ensayos se incluyó una dosis variable del nutriente de interés (0, 50, 100 y 150% de la dosis de referencia) y una dosis fija de los otros dos nutrientes (100% de la dosis de referencia); adicionalmente todos los ensayos tuvieron el mismo control absoluto (sin fertilización, N0- P0-K0) (Tab. 2). Los fertilizantes utilizados y las dosis de referencia fueron: urea, 2,6 g/planta; superfosfato triple (SPT), 1,52 g/planta; cloruro de potasio (KCl), 2,41 g/planta. Estas cantidades se aplicaron teniendo en cuenta las necesidades de nitrógeno (N), fósforo (P2O5) y potasio (K2O) reportadas por Thong y Ng (1978) para plantas de cacao en etapa de vivero. Las dosis respectivas se fraccionaron en dos aplicaciones iguales a los 56 y 64 d después de siembra, para evitar quemazón en las plantas, debido al poco espacio dentro de la bolsa para ubicar el fertilizante.
Cada planta constituyó una unidad experimental y para los análisis se utilizaron 3-6 unidades experimentales por tratamiento. Las plantas se cosecharon 124 días después de la siembra (dds) y se separaron la raíz, el tallo y las hojas. Luego, los órganos se llevaron a 104°C por 48 h para la posterior determinación de su biomasa seca y de contenido nutricional en el Laboratorio de Aguas y Suelos de la Facultad de Ciencias Agrarias de la Universidad Nacional de Colombia, sede Bogotá. El N se analizó con el método Kjeldahl, el P por calcinación a 600°C y valoración colorimétrica con molibdato y vanadato de amonio, el B por el método de Azometina (colorimétrico) y el Ca, Mg, K, Cu, Fe, Mn, Zn por calcinación a 600°C, digestión ácida y valoración por absorción atómica. Tanto la biomasa como el contenido nutricional se determinaron de forma separada por órganos en una cantidad de tres a cinco réplicas por tratamiento. No obstante, algunos resultados se presentan relacionando la raíz con la parte aérea debido a que refleja más claramente la dinámica de los nutrientes a nivel de toda la planta.
Los parámetros asociados con la dinámica nutricional se evaluaron, para todos los nutrientes analizados en el laboratorio, de forma separada para los ensayos A, B y C. A continuación, se describe su cálculo y unidades en que se presentan:
Eficiencia en la absorción del nutriente (Wang et al., 2007):
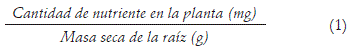
Extracción del nutriente (Ribeiro et al., 2008):
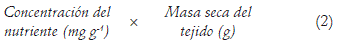
Concentración del nutriente (Ribeiro et al., 2008):
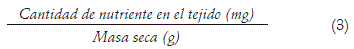
Eficiencia en el uso del nutriente (Ribeiro et al., 2008):
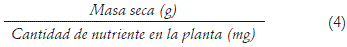
Se realizó un análisis de correlación de los parámetros mencionados para N, P y K con respecto a los mismos parámetros para Ca, Mg, K, Cu, Fe, Mn, B y Zn, de forma independiente en los ensayos A, B y C. Por último, se estimó la eficiencia agronómica (EAgr) de las diferentes dosis utilizadas en los ensayos A, B y C para N, P y K en cada ensayo. Esto se realizó a partir de la fórmula sugerida por Ghulam et al. (1996) pero modificando el rendimiento comercial por el rendimiento biológico:

Donde Rb(x) es el rendimiento biológico promedio de las plantas de cada tratamiento y Rb (N0-P0-K0) es el rendimiento biológico promedio de las plantas que no recibieron fertilización. Para facilitar la comparación, se estandarizaron los valores de EArg con respecto al mayor valor obtenido en cada ensayo.
El análisis estadístico se realizó inicialmente con un ANOVA y en los casos en que resultaron diferencias estadísticamente significativas se realizó una prueba de comparación de medias con la prueba t a un nivel de significancia de 5% utilizando el software estadístico JMP® versión 9.0.
RESULTADOS Y DISCUSIÓN
Dinámica de extracción y uso de N, P y K
Dinámica del nitrógeno
La eficiencia en la absorción de N (EAN), que indica la eficiencia en el transporte del N (Soon, 1992), aumentó en casi un 100% en las plantas que recibieron fertilización nitrogenada con respecto a las plantas que no fueron fertilizadas con este elemento (Tab. 3). Ribeiro et al. (2008) encontraron, en clones TSH-565 y ICS-9, que el aumento en la dosis de N incrementó la eficiencia en la absorción de N. La extracción N en las plantas fertilizadas con este elemento también se incrementó de forma significativa (en un 100%) y esto sucedió a nivel de raíces y de la parte aérea (Tab. 3).
La concentración natural de N en el sustrato en el que fueron sembradas las plántulas de cacao (Tab. 1) es cercana al valor utilizado por Snoeck et al. (2010) como la concentración de N en suelo mínima adecuada para el cacao (0,06%). La concentración de N en el sustrato equivale a 750 mg/bolsa lo que explica la absorción de N en las plantas sin fertilizar.
La concentración de N en las plantas fertilizadas con este elemento varió tanto por su presencia o ausencia, como por la dosis aplicada. En presencia de la dosis de referencia de P y K, las plantas fertilizadas, con la dosis de referencia de N, tuvieron una mayor concentración del elemento, que las plantas fertilizadas con la mitad de la dosis, específicamente a nivel de la parte aérea. En contraste, no se presentaron diferencias significativas en la concentración de N entre plantas que recibieron la dosis de referencia y las que recibieron mayor cantidad (N150%) lo cual sugiere que el nivel de 100% N fue suficiente. Las plantas que no recibieron fertilización nitrogenada tuvieron menores concentraciones de N con respecto a las que sí lo recibieron, lo cual, es más evidente en la parte aérea de la planta (Tab. 3).
La eficiencia en el uso del N (EUN) fue mayor en las plantas que no recibieron fertilización nitrogenada (Tab. 3). Independientemente de los tratamientos, la EUN fue superior en la raíz con respecto a la parte aérea lo cual es debido a que este órgano acumula las menores cantidades de nitrógeno que oscilan entre el 10 y 15% del total de N absorbido en comparación con el 15-27% en el tallo y con el 58-75% en las hojas (Fig. 1a). De forma similar a lo obtenido por Ribeiro et al. (2008), el aumento de la eficiencia de la absorción de N disminuyó la eficiencia en el uso del N tanto a nivel de raíz como de parte aérea.
A nivel de la parte aérea, las plantas que no recibieron ningún tipo de fertilización crecieron menos que las plantas fertilizadas. No se presentaron diferencias significativas a nivel de la biomasa total entre los tratamientos; sin embargo, la relación entre la concentración foliar de N y la biomasa seca total, describe una curva de tipo cuadrático positivo que muestra una clara separación entre las plantas fertilizadas con N y las que no lo estaban (Fig. 1b). Esto puede obedecer a que las plantas que no fueron fertilizadas con N, tuvieron una concentración foliar ligeramente menor (17,8 mg g-1) a la establecida por Wessel (1971) (18 mg g-1) como límite inferior de suficiencia para la concentración de N en hojas juveniles de cacao. La curva cuadrática positiva entre concentración foliar de nitrógeno y biomasa en cacao también fue descrita por Souza Júnior y Carmello (2008).
Se estimó que la concentración foliar de N a la que se produce mayor biomasa total es 23,89 mg g-1 de N, la cual se acerca a la obtenida con el tratamiento N50-P-K (Fig. 1). En experimentos previos con plántulas de cacao bajo sombrío artificial, también se ha encontrado que la adición de N estimula el crecimiento de una forma más evidente que en plantas adultas con sombrío natural (Wessel, 1971; Appiah et al., 2000).
Ribeiro et al. (2008) estudiaron cinco dosis de N (0, 120, 240, 360, y 480 mg N) que en su forma elemental presentaban una concentración por matera, menor a la implementada en el estudio actual. Así, es razonable que la concentración N50-P-K esté asociada a la mayor producción de biomasa.
La relación parte aérea:raíz no tuvo alteraciones significativas entre los tratamientos relacionados con la fertilización nitrogenada. Esto puede ser consecuencia de que las plantas presentaron EUN alta entre los tratamientos evaluados (Tab. 3).
Fósforo
La eficiencia en la absorción de P (EAP) varió según la fertilización con N y K y según la dosis de fertilizante fosfatado aplicado. En presencia de N y K, la EAP fue significativamente mayor en plantas fertilizadas con la dosis más alta de P (P150) con respecto a las dosis P50 y P0. De otra parte, las plantas que no fueron fertilizadas (N0-P0-K0) tuvieron una EAP similar a la estimada con plantas fertilizadas con P150 (Tab. 4).
A nivel de toda la planta, tanto la ausencia de fertilización (N0-P0-K0) como la fertilización con dosis completa y alta de P (N-P100-K) (N-P150-K), respectivamente, mostraron una extracción similar de P, mientras que la dosis baja (P50) y la ausencia de fertilización fosfatada causaron menor extracción de fósforo cuando las plantas se fertilizaron con N y K. A nivel de la raíz y la parte aérea, esto fue más evidente con respecto a la dosis baja de P.
Las concentraciones foliar de P en los tratamientos N-P0-K, N-P50-K y N-P100-K se encontraron por debajo de los niveles de suficiencia de P para hojas jóvenes establecido por Wessel (1971) (2,0 mg g-1), especialmente las dos primeras dosis.
El tratamiento que no tuvo ningún tipo de fertilización tuvo la mayor concentración, tanto para la raíz, como para el tallo y las hojas (Fig. 2a). El aumento en la extracción y concentración de P en el tratamiento N0-P0-K0 con respecto a los demás tratamientos, pudo deberse a un desbalance entre la relación N/P a nivel del suelo. De forma particular, se encontró que a medida que la relación N/P2O5 (disponibles), disminuía, aumentaba la eficiencia en la absorción de P (Tab. 4). Lotodé y Jadin (1981) propusieron que la medida apropiada de esta relación N/P a nivel de suelo sería entre 1,5 y 2 de N/P2O5 (totales). Dentro de este rango, pero considerando N y P2O5 disponibles solo se ubica el tratamiento N100-P100-K100. Hidalgo (1989) también realizó esta modificación para P2O5 en suelos de Costa Rica.
Las plantas fertilizadas (N100-Px-K100) fueron más eficientes en el uso del P que las no fertilizadas (N0- P0-K0). De otra parte, no se presentaron diferencias significativas en la producción de biomasa de las plantas sometidas a los diferentes tratamientos. Esto fue debido a que la eficiencia en el uso del P disminuyó cuando su concentración aumentó. Consecuentemente, se observó que la relación entre biomasa total y concentración foliar de P correspondió a una tendencia lineal negativa que indica la relación inversa entre concentración de P y el crecimiento dentro del rango evaluado (Fig. 2b).
Las plantas que tuvieron la mayor concentración de P (N0-P0-K0) y la menor producción de biomasa, también mostraron una relación de concentración foliar N/P significativamente menor (4,09±0,46) a la misma relación en los otros tratamientos (12,19±4,69, datos no mostrados). Puentes et al. (2016) encontraron que los rendimientos de cacao disminuían de manera importante con una relación N/P menor a cinco, mientras que los mayores rendimientos se obtuvieron a una relación de nueve.
Finalmente, las plantas sometidas a la dosis alta de P presentaron una disminución en la biomasa de raíz que modificó significativamente la relación parte aérea: raíz (Tab. 4), lo que podría confirmar la relación inversa descrita previamente.
Potasio
La eficiencia de la absorción de K (EAK) no cambió significativamente en los tratamientos del ensayo C. A nivel de toda la planta y de la raíz principalmente en las que fueron fertilizadas con N y P (N100-P100-Kx), la extracción de K fue mayor en la dosis K100, que en las no fertilizadas con este elemento. Sin embargo, no hubo diferencias significativas a nivel de la parte aérea a pesar que es allí en donde hay mayor acumulación de K (Tab. 5). El contenido natural de K en el sustrato utilizado correspondió a 225 mg, lo que explica la extracción de K en los tratamientos con K0.
A excepción de las plantas sin fertilizar (N0-P0-K0), la concentración de K aumentó con la dosis creciente de fertilizante potásico. La mayor concentración de potasio en tejidos se presentó cuando las plantas fueron fertilizadas con la dosis alta de K (K150), especialmente a nivel del tallo y la menor en plantas sin fertilización potásica, pero fertilizadas con N y P (N100-P100-K0) especialmente a nivel de tallo y raíz (Fig. 3a). En cacao, los altos niveles de N podrían afectan la absorción de K (Puentes et al., 2016).
Las plantas sin fertilizar (N0-P0-K0) presentaron una concentración mayor de K que las plantas fertilizadas con N100-P100-K0, pero similar a las de los demás tratamientos. Esto correspondió a una inadecuada relación entre las bases totales intercambiables (BTI) y el N. Snoeck et al. (2016) reportaron que la relación óptima entre las BTI y el N obedece a una línea de regresión BTI=8,9*N(â°) - 6,15. La BTI calculada con la concentración de N (0,8â°) es de 0,97, lo que indica que el tratamiento sin fertilizar presentaba una deficiencia de N a expensas de las bases intercambiables, incluido el K (datos no mostrados); situación que se corrigió con la aplicación de N en los demás tratamientos.
El tratamiento sin fertilización nitrogenada, mostró una relación de concentración foliar N/K significativamente menor (0,7±0,17) a la misma relación en los otros tratamientos (1,13±0,15, datos no mostrados) y que se acerca a la sugerida por Puentes et al. (2016) para cacao. Sin embargo, no se observó una relación clara entre la magnitud de esta relación y los resultados de biomasa.
La eficiencia en el uso de K (EUK) fue menor en la dosis alta de K que en las otras dosis, especialmente en la parte aérea. Por su parte, la mayor EUK se presentó en N-P-K0. La EUK de las plantas sin fertilizar (N0-P0-K0), fue similar a la de plantas fertilizadas con K en presencia de N y P. Esto obedece básicamente a la disminución de la EUK al incrementar la concentración del K.
El peso seco de las plantas fertilizadas con dosis K50 fue mayor que en los demás tratamientos. Se determinó una función cúbica a partir de la relación de la concentración foliar de K y la biomasa seca total (Fig. 3b). Según esta función, la concentración foliar de K que corresponde al mayor valor de biomasa es 19,0 mg g-1, que es muy cercana a la determinada en el tratamiento (N-P-K50). Según Snoeck et al. (2007), los porcentajes de K, Ca y Mg deben ser 8, 68 y 24%, respectivamente. En el sustrato utilizado el K tiene el 4,3% de las bases intercambiables en tanto que con K50 sube a 14,2%, lo que por una parte, explica la mejor absorción de K y, por otra, abre la posibilidad de un efecto negativo por encima de esa dosis, coherente con la función cúbica descrita en la figura 3b. Previamente han sido reportados los efectos negativos del potasio en exceso sobre el crecimiento de plantas de cacao en etapa de vivero (Frederick, 1961; Sodré et al., 2012).
De otra parte, la variación en la fertilización no tuvo efecto sobre la relación parte aérea:raíz en las plantas de cacao lo que obedece al aumento en la EUK de las plantas que tenían una menor concentración de K. Es probable que los requerimientos por K no sean altos para este clon en etapa de vivero. Ahenkorah et al. (1987) sugirieron que los requerimientos de K en el cacao tienden a incrementarse con la edad y en especial, bajo condiciones de poco sombrío.
Correlación de N, P y K con otros nutrientes
En general, el N se correlacionó negativamente con P, Mg, Cu, Zn y B (concentración y eficiencia en el uso del nutriente) y positivamente con K y Fe (eficiencia en su absorción y extracción), y con Ca y Mn (eficiencia en su absorción, extracción, concentración y eficiencia en el uso) (Tab. 6).
Los patrones de correlación negativa y positiva de N con los otros nutrientes son similares a los obtenidos por Souza y de Camargo (2008) en plantas de cacao en etapa de vivero, a excepción del K. Estos autores atribuyen a un efecto de dilución de los nutrientes correlacionados negativamente lo cual es posible que haya sucedido en este estudio por cuanto se presentaron a nivel de concentración y eficiencia en el uso del nutriente, pero no a nivel de absorción. Sin embargo, solo se lograron detectar correlaciones negativas significativas entre biomasa total y la concentración total de Zn (-0,65p=0,0022) y B (-0,46p=0,0451).
Las correlaciones de N con Mg y Zn fueron positivas en cuanto a la eficiencia en la absorción y en la extracción, pero fueron negativas en la concentración y en la eficiencia en el uso del nutriente, principalmente en el ensayo C. Esto se debe a que las dosis de K150% parecieron limitar la absorción tanto de N como de Mg y Zn (datos no mostrados), en tanto que a K50% y K100% la absorción de N, Mg y Zn aumentaron, indicando así que las altas concentraciones de K podrían deprimir la absorción de otros nutrientes (Fageria, 2001). En cacao este fenómeno había sido reportado para Mg, Ca y Fe (Frederick, 1961; Sodré et al., 2012). No obstante, la correlación de N con respecto a Mg y Zn fue primordialmente negativa debido a efecto de dilución causado en los tratamientos con fertilización nitrogenada.
Además del efecto de dilución y de forma similar al desbalance entre N y P descrito anteriormente, en el sustrato, el porcentaje de Mg (39,3%) con respecto a las otras bases intercambiables (K y Ca), fue superior a la sugerida por Snoeck et al. (2007) (24%), así que, cuando hubo fertilización con N la absorción de Mg disminuyó y las de K y Ca, aumentaron; por el contrario, cuando no hubo fertilización con N, la absorción de Mg aumentó.
La correlación positiva observada entre N y K, a diferencia de lo reportado por Souza y Carmello (2008), no correspondió a un efecto de dilución porque aun con la menor dosis de K (K50), se suministró la cantidad suficiente de este elemento en el sustrato. Esto mismo pudo suceder con Ca, Fe y Mn resultando en una interacción positiva. Sumado a esto, la interacción positiva de estos nutrientes con el N en cuanto a la absorción sugiere una sinergia a nivel de su transporte a larga distancia. Por una parte, tanto el K como el Ca se han asociado con el transporte de N-NO3 por el xilema (Mengel y Kirkby, 2001) y por otra parte, Ca, Fe y Mn pueden ser transportados en el xilema como cationes acompañantes de ácidos orgánicos (Marschner, 2012), cuya producción puede ser mayor en plantas fertilizadas con N dado su efecto positivo sobre la fotosíntesis.
En conjunto, estos resultados obedecen a que el incremento en el suministro de N, aumenta el crecimiento de las plantas, incrementando la demanda por otros nutrientes y en algunos casos causando posibles deficiencias (Fageria, 2001).
El P solamente se correlacionó negativamente con Ca y Mn, lo que puede obedecer a un efecto indirecto dada la estrecha correlación positiva entre estos dos nutrientes y el N, y la correlación negativa entre N y P, por las razones expuestas previamente (Tab. 6). Finalmente, el K se correlacionó positivamente con todos los nutrientes evaluados en este estudio, especialmente a nivel de la eficiencia en la absorción y en la extracción. Sodré et al. (2012) encontraron que la aplicación de K aumentó la concentración de Ca, Mg, Zn y Mn disponibles en el sustrato, en tanto que la disponibilidad de Cu y Zn no se alteraron y las de Al y Fe disminuyeron. Las diferencias radican en que Sodré et al. (2012) utilizaron un sustrato con mayor cantidad de materia orgánica lo que facilitó la complejización de los cationes con ácidos orgánicos. En el presente estudio, la cantidad de materia orgánica fue baja (1,05%) lo cual explica la mayor disponibilidad de cationes.
Eficiencia agronómica
En el ensayo A, la eficiencia agronómica del N fue mayor en N50-P-K, así por cada gramo de N aplicado, se obtuvo un incremento de 4,22 g en la biomasa seca, en tanto que la eficiencia agronómica del P y del K fueron más alta en N150-P-K (8,53 g g-1 y 2,19 g g-1, respectivamente). Sin embargo, en N50-P-K esta eficiencia solo se disminuyó un 4% (8,19 g g-1 y 2,11 g g-1, respectivamente) (Fig. 4a). Dado que la dosis N150-P-K requiere tres veces más cantidad de N que la dosis N50-P-K, se sugiere que la dosis más apropiada de N es N50-P-K.
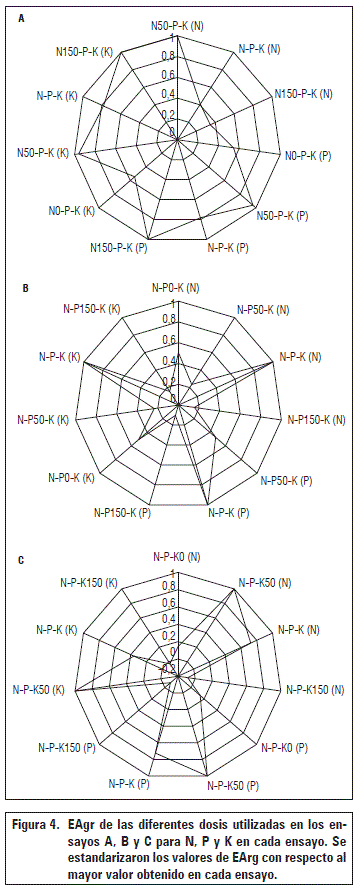
En el ensayo B, la eficiencia agronómica de los tres elementos aplicados fue mayor en la dosis N-P-K . Así, N tuvo una eficiencia de 1,72 g g-1, P de 6,68 g g-1 y K de 1,72 g g-1 (Fig. 4b). En el ensayo C, la eficiencia agronómica de los tres elementos aplicados fue mayor en la dosis N-P-K50, así N tuvo una eficiencia de 2,37 g g-1, P de 9,23 g g-1 y K de 4,75 g g-1 (Fig. 4c).
Los resultados de la EAgr en los tres ensayos siguen un patrón que es coherente con la dinámica nutricional descrita a lo largo de este documento. Consecuentemente, diferencias en las magnitudes probablemente corresponden a desbalances nutricionales. Puentes et al. (2014b) encontraron notorias diferencias en varios tratamientos de fertilización de cacao en campo y lo explicaron a partir de desbalances nutricionales del cultivo asociados a cambios en la composición de la solución del suelo.
En las dosis superiores a N50 y especialmente a N150 se presentaron efectos de dilución de otros nutrientes y de consumo de lujo en tanto que en las dosis superiores a K50 se presentaron desbalances con respecto al TBI (en K150), lo que explicaría porque N50 y K50 tuvieran la mayor EAgr. De hecho, Wessel (1971) encontró, a partir de una revisión de investigaciones desarrolladas en Ghana, que las plantas jóvenes de cacao son sensibles al N y al K.
La mayor EAgr del P se obtuvo en el 100% de la dosis debido a que, cuanto estaba en presencia de N100 y K100, parece haberse obtenido un balance apropiado en la solución del suelo que permitió, no solo una alta extracción de P, sino también una alta eficiencia en su uso, a diferencia del tratamiento N0-P0-K0.
Tomado en conjunto, los resultados con los tres grupos de tratamientos evaluados sugieren que los futuros estudios en nutrición de plantas de cacao en vivero bajo las condiciones evaluadas, deberían incluir el estudio de relaciones entre los nutrientes a nivel del suelo y de la planta utilizando dosis de Urea (1,3 g/ planta), SPT (1,52 g/planta) y KCl (1,205 g/planta). Esto sería un avance importante para la obtención de material vegetal de cacao de alta calidad desde el punto de vista nutricional.
CONCLUSIONES
La dinámica nutricional de N, P y K cambió en relación a los tratamientos implementados. Así, la dinámica del N en cacao se relacionó con su concentración en el sustrato y mostró un valor crítico en términos de tejido foliar (23,89 mg g-1) por encima del cual el consumo no fue eficiente. La dinámica del P en cacao se modificó con la relación N/P2O5 (disponibles) en el suelo que fue más favorable en un valor de 1,58. La dinámica del K en cacao se modificó con la relación entre las BTI y entre estas y el N en el suelo.
La dinámica nutricional del N, P y K afectó la dinámica prácticamente de todos los otros nutrientes evaluados, principalmente debido a los desbalances a nivel del sustrato y al efecto de dilución cuando aumenta el tamaño de las plantas al ser fertilizadas con N. Por último, es importante resaltar que la dinámica e interacciones entre los nutrientes fueron consistentes en los tres ensayos evaluados y que la eficiencia agronómica de todos los tratamientos de fertilización implementados es coherente con la dinámica de los nutrientes analizados.
La importancia de estos resultados radica en que se contribuye al entendimiento del manejo de los balances nutricionales más que de las dosis de fertilizantes para la obtención de material vegetal de calidad para trasplante en cultivo de cacao en Colombia.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos a Colciencias por la financiación de esta investigación a través del proyecto "Análisis de tecnologías novedosas para el cultivo del cacao en el departamento de Casanare- Colombia" código 1101-569-33833.
REFERENCIAS BIBLIOGRÁFICAS
Agronet. 2014. Estadísticas cacao Colombia. En: http://www.agronet.gov.co; consulta: febrero de 2015. [ Links ]
Ahenkorah, Y., B. Halm, M. Appiah, G. Akrofi y J. Yirenkyi. 1987. Twenty years’ results from a shade and fertilizer trial on Amazon cocoa (Theobroma cacao) in Ghana. Exp. Agric. 23, 31-39.
Appiah, M., K. Ofori-Frimpong y A. Afrifa. 2000. Evaluation of fertilizer application on some peasant cocoa farms in Ghana. Ghana J. Agric. Sci. 33, 183-190. [ Links ]
Clough, Y., H. Faust y T. Tscharntke. 2009. Cacao boom and bust: sustainability of agroforests and opportunities for biodiversity conservation. Conserv. Lett. 2(5), 197- 205. Doi: 10.1111/j.1755-263X.2009.00072.x [ Links ]
Fageria, V. D. 2001. Nutrient Interactions in crop plants. J. Plant Nutr. 24(8), 1269-1290. Doi: 10.1081/PLN-100106981 [ Links ]
Frederick, H. 1961. Manual de cacao. Antonio Lehmann, Turrialba, Costa Rica. [ Links ]
García L., J., M. Romero C. y L.A. Ortiz. 2005. Evaluación edafoclimática de las tierras del trópico bajo colombiano para el cultivo de cacao. Corpoica, Produmedios, Mosquera, Colombia. [ Links ]
Ghulam, H., A. Karimulla y S. Al-Jaloud. 1996. Effect of treat de fluent irrigation and nitrogen on yield and nitrogen use efficiency of wheat. Agric. Water Manage. 30, 175-184. [ Links ]
Hartmann, H.T., D.E. Kester, F.T. Davies y R.L. Geneve. 2011. Plant propagation: principles and practices. 6th ed. Prentice-Hall, Englewood Cliffs, NJ. [ Links ]
Hidalgo, E. 1989. Propiedades fisicoquímicas de los suelos y respuestas a la fertilización en microparcelas en áreas para cacao. CATIE, Turrialba, Costa Rica. [ Links ]
Lotodé, R. y P. Jadin. 1981. Calcul des besoins en engrais des cacaoyers. Café Cacao Thé. 25(1), 4-24. [ Links ]
MADR. 2012. Plan de Desarrollo Cacaotero 2012-2021. Ministerio de Agricultura y Desarrollo Rural, Bogotá [ Links ].
Marschner, P. (ed.). 2012. Marschner’s mineral nutrition of higher plants. 3rd ed. Elsevier, Amsterdam, The Netherlands.
Mengel, K., E. Kirkby, H. Kosegarten y T. Appel (eds.). 2001. Principles of plant nutrition. 5th ed. Kluwer Academic Publishers, Dordrecht, The Netherlands. [ Links ]
Oliveira, R., J. Olimpio, G. Sodré y B. Chanabasappa. 2015. Growth and nutrition of cacao seedlings influenced by zinc aplication in soil. Rev. Bras. Frutic. 37(4), 1053-1064. [ Links ]
Oyewole, O.S., O.J. Ajayi e I. Rotimi. 2012. Growth of cocoa (Theobroma cacao L .) seedlings on old cocoa soils amended with organic and inorganic fertilizers. Afr. J. Agric. Res. 7(24), 3604-3608. Doi: 10.5897/AJAR11.2045 [ Links ]
Puentes P., Y.J., A.G. Carabalí y J.C. Menjivar F. 2016. Influence of the relationship among nutrients on yield of cocoa (Theobroma cacao L.) clones. Acta Agron. 65(2), 176-182. [ Links ]
Ribeiro, M.Q., J. O. da Silva, W.M. Aitken, R.C.R. Machado y V.C. Baligar. 2008. nitrogen use efficiency in cacao genotypes. J. Plant Nutr. 31(2), 239-249. Doi: 10.1080/01904160701853720 [ Links ]
Santiago, M.A., I.R. Canacuan, J.J. Franco, W. Bohórquez y J.C. Fernández. 2016. Determinación de los estados críticos de nutrientes de cacao en etapa de vivero. pp. 191-204. En: Construyendo investigación: semilleros generadores de ideas. Ediciones Unisalle, Bogotá [ Links ].
Snoeck, D., M.K. Abekoe, A.A. Afrifa y M.R. Appiah. 2007. The Soil diagnostic method to compute fertilizer requirements in cocoa plantations. pp. 7-11. En: Proc. 15th International Cocoa Research Conference, Lagos. [ Links ]
Snoeck, D., A. A. Afrifa, E. Boateng y M. K. Abekoe. 2010. Mapping Fertilizer Recommendations for Cocoa Production in Ghana Using Soil Diagnostic and GIS Tools. West Afr. J. Appl. Ecol. 17, 97-107. [ Links ]
Snoeck, D., L. Koko, J. Joffre, P. Bastide y P. Jagoret. 2016. Cacao nutrition and fertilization. pp. 155-202. En: Lichtfouse, E. (ed.). Sustainable Agriculture Reviews 19, Springer International Publishing, Suiza. [ Links ]
Sodré, G.A., M.T. Venturini, D. Ribeiro y C. Lima. 2012. Extrato da casca do fruto do cacaueiro como fertilizante potássico no crescimento de mudas de cacaueiro. Rev. Bras. Frutic. 34(3), 881-887. [ Links ]
Soon, Y. 1992. Differential response of wheat genotypes to phosphorus in acid soils. J. Plant Nutr. 15, 513-526. [ Links ]
Souza, J.O. y Q.A. de Camargo. 2008. Formas de adubação e doses de uréia para mudas clonais de cacau cultivadas em substrato. Rev. Bras. Ciênc. Solo 32, 2367-2374. Doi: 10.1590/S0100-06832008000600015 [ Links ]
Souza, J.O. y Q.A. de C. Carmello. 2008. Formas de adubação e doses de uréia para mudas clonais de cacau cultivadas em substrato. Rev. Bras. Ciênc. Solo 32(6), 2367-2374. [ Links ]
Thong, K.C. y W.L. Ng. 1978. Growth and nutrient composition of monocrop cocoa plants on inland Malaysian soils. pp. 262-286. Proc. Int. Conf. Cocoa coconuts, Kuala Lumpur 1978. [ Links ]
Van Vliet, J., M. Slingerland y K.E. Giller. 2015. Mineral nutrition of cocoa : a review. Wageningen University and Research Centre, Wageningen, The Netherlands. [ Links ]
Wang, F.Y., X.G. Lin y R. Yin. 2007. Inoculation with arbuscular mycorrhizal fungus Acaulospora mellea decreases Cu phytoextraction by maize from Cu-contaminated soil. Pedobiol. 51(2), 99-109. Doi: 10.1016/j.pedobi.2007.02.003 [ Links ]
Wessel, M. 1971. Fertilizer requirements of cacao (Theobroma Cacao L.) in South-Western Nigeria. Koninklijk Institut voor de Tropen, Amsterdam, The Netherlands. [ Links ]
World Cocoa Foundation. 2014. Cocoa market update. 1-7 p. En: http://www.worldcocoafoundation.org/wp-content/uploads/Cocoa-Market-Update-as-of-4-1-2014.pdf; consulta: febrero de 2016. [ Links ]

