











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
El tomate de árbol [Cyphomandra betacea (Cav.) Sendt.], es un frutal representativo y originario de los Andes americanos de buena aceptación y alta demanda por su contenido de vitaminas, agradable sabor, color y con posibilidades de consumo fresco y potencial agroindustrial (Lobo, 2006; Lagos et al., 2012). Es un frutal de climas templados y fríos, se desarrolla bien en temperaturas entre 13 y 24°C, siendo la óptima entre 16 y 19°C. Esta planta se desarrolla en altitudes que varían de 1.000 a 3.000 msnm (Lagos et al., 2012).
En Colombia, la superficie cultivada de tomate de árbol alcanza aproximadamente las 8.107 ha y un rendimiento promedio de 13 t ha-1 (Agronet, 2014), siendo los departamentos de Antioquia, Huila, Nariño y Boyacá los principales productores.
En Nariño se reporta una área sembrada de 487 ha con una producción de 2.844 t y un rendimiento de 10 t ha-1 con una diferencia de 3 t ha-1 con respecto al promedio nacional (Agronet, 2014). Sin embargo, este incremento se ha visto afectado por la escasa oferta de híbridos o variedades mejoradas que suplan las necesidades del productor y ayuden a resolver los problemas fitosanitarios causados por patógenos de importancia como Phytophthora infestans sensu lato, Colletotrichum acutatum, virus, entre otras, los cuales afectan el rendimiento y la calidad del producto, reduciendo significativamente los ingresos del agricultor (Arturo et al., 2003).
Dentro de las enfermedades más limitantes para el cultivo de las Solanaceas se encuentra la gota o tizón tardío causado por el oomycete Phytophthora infestans sensu lato. Este patógeno es de una gran agresividad no solo en el cultivo del tomate de árbol (Cyphomandra betacea), sino también tomate de mesa (Lycopersicon esculentum) y en la papa (Solanum tuberosum), destruyendo el follaje, los tallos y el fruto o el tubérculo (en el caso de la papa), en cualquier momento de desarrollo de la planta y hasta el almacenamiento (Pérez y Forbes, 2008). En Colombia la presencia de este patógeno reporta que las pérdidas pueden alcanzar hasta un 100% en la producción y su incidencia se atribuye a la disponibilidad de fuentes constantes de inóculo en campo y a la ubicación de sus hospederos en regiones con condiciones ambientales favorables para el establecimiento de poblaciones del patógeno (García et al., 2008).
Teniendo en cuenta lo anterior, el objetivo de este estudio fue evaluar la reacción de 29 genotipos de tomate de árbol frente a la enfermedad Phytophthora infestans sensu lato, bajo condiciones de laboratorio.
Materiales y métodos
Localización
El presente trabajo se desarrolló en el laboratorio de microbiología y genética a cargo del Grupo de Investigación Genética y Evolución de Organismos Tropicales (GENPAT), en la Universidad de Nariño, sede Torobajo, ubicado a 2.450 msnm con una temperatura promedio de 18°C.
Material vegetal
Los genotipos de tomate de árbol (Cyphomandra betacea) hacen parte de la colección de trabajo del Grupo de Investigación en Producción de Frutales Andinos de la Universidad de Nariño. Se utilizaron un total de 29 genotipos provenientes de las zonas productoras de Nariño (Tab. 1), con diferentes características como variabilidad en tipo, calidad de fruto, respuesta a enfermedades y capacidad de carga (Valencia et al., 2011).
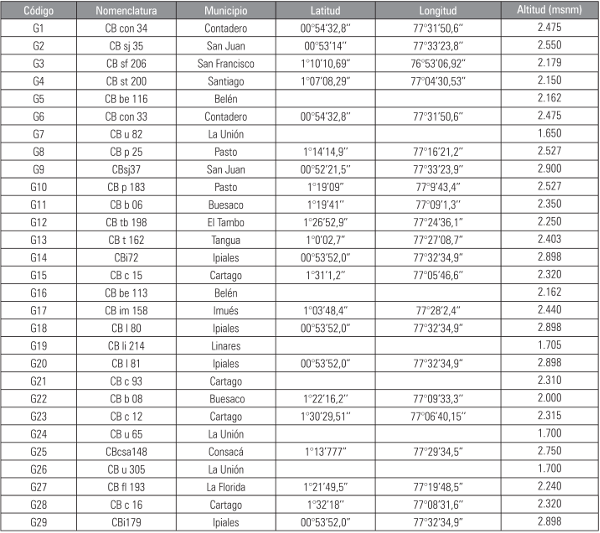
Aislamientos de Phytophthora infestans sensu lato
Las cepas utilizadas en esta investigación hacen parte de la colección de P. infestans sensu lato a cargo del Grupo de Investigación Genética y Evolución de Organismos Tropicales (GENPAT) de la Universidad de Nariño; estas cepas fueron colectadas en el año 2009 en cultivos de C. betacea y son mantenidas en agar tomate-arveja a 18°C (Tab. 2).
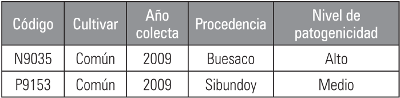
Estas cepas fueron seleccionadas teniendo en cuenta el nivel de patogenicidad reportado por GENPAT (2011) en el informe “Caracterización genética y fenotípica de las poblaciones de las poblaciones de Phytophtora infestans sensu lato procedentes de tomate de árbol (Cyphomandra betacea) en las zonas productoras de Nariño”.
Inoculación en hoja desprendida (in leaf)
La metodología consistió en utilizar hojas de planta de tomate de árbol de 5 a 7 meses de edad, provenientes de semilla sexual. Los cuales fueron evaluados frente a dos cepas de Phytophthora infestans sensu lato. La inoculación de las hojas se realizó siguiendo la metodología propuesta por Carlisle et al. (2002). Se preparó una solución de 4.5×105 esporangios mL-1 a partir de cultivos de 15 d de crecimiento (agar arveja), la suspensión de esporangios se dejó 3 h en nevera a una temperatura de 4°C antes de inocular con 25 μL el material vegetal en cuatro puntos. Las hojas se mantuvieron en cámaras húmedas por separado con el fin de mantener las condiciones apropiadas para que el proceso de infección fuera exitoso.
Se realizó un registro fotográfico durante 10 d a partir del tercer día de inoculación, el área de infección se calculó empleando el programa Image J 1.45s (Waine Rasband, National Institutes of Health, Bethesda, MD).
Diseño experimental
El diseño experimental empleado en esta investigación fue un Diseño de Bloques Completos al Azar (BCA). Los genotipos se evaluaron por separado frente a dos cepas P. infestans sensu lato, identificadas como N9035 y P9153, siendo 58 tratamientos en triplicado con tres repeticiones en el tiempo.
Variables evaluadas
Tamaño de la lesión (TL). El tamaño de la lesión se calculó a partir de los registros fotográficos para cada punto de inoculación. El valor de la lesión utilizado para el análisis estadístico fue la medida al último día de evaluación siguiendo el principio propuesto por Chacón et al. (2007).
Área bajo la curva de progreso de la enfermedad (AUDPC). Para cada genotipo se calculó el Área Bajo la Curva de Progreso de la enfermedad - AUDPC (Shaner y Finney, 1977), expresada como:
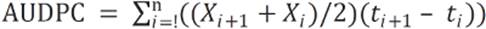 (1)
(1)
Donde
Xi |
área de tejido afectado en la observación i-ésima |
ti |
tiempo en días en la observación i-ésima |
n |
número de observaciones |
Eficiencia de infección (EI). Se define como la proporción de sitios inoculados que desarrollaron la lesión en todas las réplicas por aislamiento (Lebreton et al., 1999). Teniendo en cuenta la escala propuesta por López et al. (1997) los genotipos presentan tres tipos de respuestas diferentes: reacción positiva (necrosis múltiple acompañada por la esporulación del patógeno), reacción negativa (no presenta lesión ni necrosis) y reacción de hipersensibilidad (formación de pequeñas manchas o puntos negros con un halo clorótico) (Fig. 1).

Figura 1 Reacciones de Cyphomandra betacea frente al ataque de Phytophthora infestans sensu lato. Reacción positiva (A), reacción negativa (B) y reacción de hipersensibilidad (C).
Tasa de esporulación (TS). Definida como el total de esporangios por el área de la lesión al último día de evaluación, se calculó realizando el conteo de esporangios en cámara de Neubauer después de cortar las lesiones, pasarlas por vortex y preparar la solución (Carlisle et al., 2002; Lebreton et al., 1999).
Periodo de incubación (PI). Se calculó como el número promedio de días que han transcurrido desde la inoculación hasta el primer día que aparecen los síntomas de la enfermedad en cada sitio inoculado (Chacón et al., 2007).
RESULTADOS Y DISCUSIÓN
Se evaluaron 29 genotipos frente a las cepas N9035 y P9153. El análisis estadístico no mostró diferencias significativas para los genotipos inoculados con la cepa P9153, por el contrario, genotipos que fueron inoculados con la cepa N9035 presentaron diferencias significativas para las variables área bajo la curva de progreso de la enfermedad, eficiencia de infección y tamaño de lesión.
Área bajo la curva de progreso de la enfermedad (AUDPC)
Se encontró que existen diferencias significativas (P≤0,05) en el análisis del crecimiento del patógeno a través del tiempo en los diferentes genotipos (Tab. 3). La prueba de comparación de medias LSD de Fisher mostró que el genotipo G1 difiere significativamente de los demás (Tab. 4) presentando el valor más bajo de AUDPC (15,554), al ser el genotipo menos afectado por la enfermedad y siendo identificado posiblemente como tolerante, esto se debe probablemente a que las plantas poseen mecanismos inducidos de defensa también conocidos como resistencia inducida que se activan solamente como una respuesta al ataque de un patógeno. La resistencia inducida es un mecanismo activo de defensa que involucra cambios claros en el metabolismo provocados por la expresión de genes. Por lo tanto, para que ocurra la inducción de la defensa es necesaria la mediación de sistemas de reconocimiento específico, mediante los cuales la planta reconoce la presencia del patógeno (Madriz, 2002).
Tabla 3 ANDEVA combinado para las variables AUDPC, EI y TL (datos transformados según la fórmula y = log X).
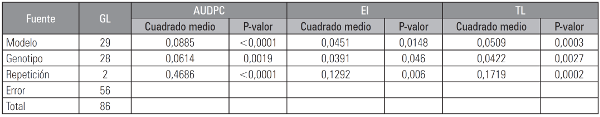
Área bajo la curva del progreso de la enfermedad (AUDPC); eficiencia de infección (EI); tamaño de lesión (TL).
Tabla 4 Prueba de significancia de medias LSD de Fisher para AUDPC (datos transformados según la formula y = log X).
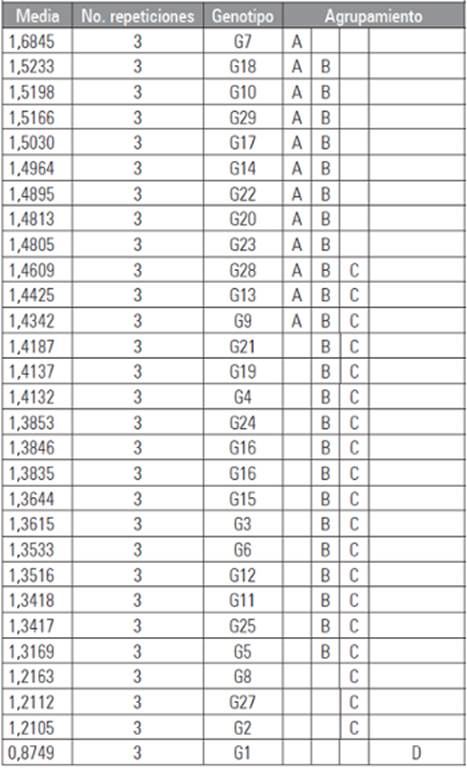
Promedios con letras distintas indican diferencia significativa según la pruebaLSD de Fisher (P≤0,005).
Por otra parte, los genotipos G7, G10 y G18 presentaron los valores más altos (47,817; 48,277 y 58,624, respectivamente) los cuales reflejan un elevado crecimiento del patógeno en un corto periodo de tiempo (Fig. 2). Este comportamiento se debe a la capacidad que tiene un planta de servir como hospedante y de su reacción frente al patógeno lo cual depende de su constitución genética y la interacción entre el patógeno y los tejidos de la planta (Niks y Linfhout, 2004). Además de que se presentó una interacción planta-patógeno compatible (hospedante susceptible, patógeno virulento) donde el reconocimiento por parte de la planta no se lleva a cabo, la respuesta de defensa no es activada y la enfermedad se establece (De Wit, 1997).
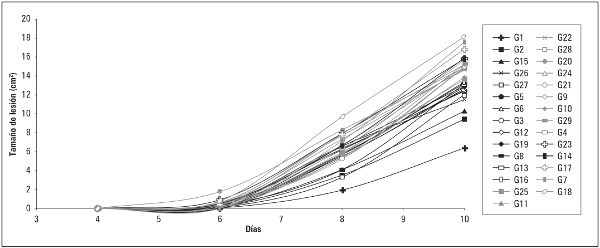
Figura 2 Área bajo la curva del progreso de la enfermedad de P. infestans sensu lato en los genotipos de C. betacea (Cav.) Sendt.
El Centro Internacional de la Papa (CIP) recomienda el uso de AUDPC para sintetizar las lecturas de tamaño de lesión de la gota de una manera más simple y clara (Fry, 1978), además autores como Jaramillo (2003) resaltan que los valores de AUDPC son de gran utilidad para estimar el nivel de susceptibilidad en las plantas.
El uso de este parámetro para diferenciar la resistencia en diferentes accesiones ha sido muy común en papa (Carlisle et al., 2002; Grünwald y Flier, 2005; Chacón et al., 2007) y otros hospederos como tomate de mesa (Suassuna et al., 2004) considerándose en todos los casos una buena aproximación para determinar el tipo de reacción. Esto hace de la técnica de infección sobre tejido vivo un medio confiable para el estudio del comportamiento tanto del oomiceto como de la planta (Díaz et al., 2003). Por esta razón, este tipo de investigaciones se constituyen como una importante fuente de información para la obtención de material vegetal con potencial productivo y cierto grado de tolerancia a enfermedades (Portilla y Salas, 2007).
Eficiencia de Infección (EI)
Para la variable eficiencia de infección, se encontraron diferencias significativas entre los genotipos evaluados (P≤0,05) (Tab. 3). En la prueba de comparación de medias LSD se destacan los genotipos G1, G4, G6 y G15 (Fig. 3) los cuales presentan los valores más bajos para esta variable con un valor promedio de 59% de infección positiva (Fig. 3), dentro de estos se destaca el genotipo G1 que presentó el valor más bajo con un 39% de respuestas positivas situación que coincide con los valores bajos reportados para las variables AUDPC y tamaño de lesión. Por el contrario, los 25 genotipos restantes presentaron un promedio para eficiencia de infección de 89%, del grupo se destacan los genotipos G9, G11, G20, G23 y G26 los cuales mostraron los valores más altos con un 100% de respuestas positivas. Estos porcentajes de infección indican posiblemente una alta susceptibilidad de la planta frente al ataque del patógeno, lo que se debe posiblemente a que existe una interacción compatible (genotipo susceptible, patógeno virulento), en la cual la planta no posee unos genes de resistencia para contrarrestar a cierto patógeno virulento, por lo cual el reconocimiento no se lleva a cabo, la planta no activa una eficiente respuesta de defensa permitiendo la penetración, invasión y multiplicación del patógeno en sus tejidos, produciendo la sintomatología del patógeno (De Wit, 1997; Stange et al, 2007). Las respuestas de los genotipos G1, G4, G6 y G15 con promedios de infección bajos pueden estar asociados a factores como la calidad nutricional, elementos estructurales, cargas enzimáticas, entre otras (Sañudo et. al, 2003). Es probable que manifestaran sistemas de defensa preexistentes o que interactuaran con el sistema de ataque del patógeno, controlando la extensión del desarrollo de la enfermedad, manteniéndola a niveles moderadamente bajos (Collinge et al., 2001).
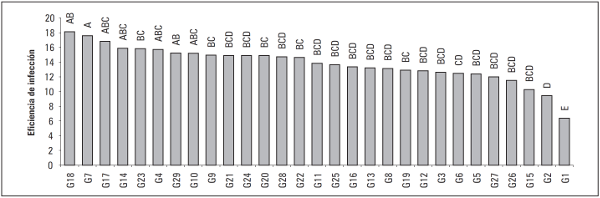
Figura 3 Prueba de significancia de medias LSD de Fisher (P≤0,005) para la variable eficiencia de infección.
La importancia del parámetro eficiencia de infección es destacada por Lebreton et al. (1999) y Carlisle et al. (2002) para estimar el nivel de susceptibilidad de algunas especies frente a P. infestans sensu lato, estos reportes son consistentes con los resultados obtenidos en este estudio, siendo este uno de los parámetros determinantes para diferenciar los genotipos entre sí.
Tamaño de lesión (TL)
Para esta variable, el análisis estadístico no encontró diferencias significativas entre los genotipos con excepción del genotipo G1 en el cual la prueba de comparación de medias LSD mostró que este genotipo difiere de los demás (P≤0,05) (Fig. 4) presentando el área más pequeña afectada al décimo día de evaluación con un área en promedio de 6,380 cm2, lo que se debe posiblemente a la presencia de un gen R específico produciendo una reacción incompatible, la planta activa sus mecanismos de defensa efectivos llevando a cabo el reconocimiento del patógeno, impidiendo su penetración, invasión y multiplicación, lo que se manifiesta como resistencia (Stange et al., 2007). Este valor coincide con los reportados para la variable AUDPC y EI. Los demás genotipos no presentaron diferencias estadísticas en esta variable con excepción de aquellos en que el tamaño de lesión fue significativamente superior (P≤0,05) al promedio, el cual fue 14,037 cm2 siendo el genotipo G18 con el tamaño de lesión más grande con 18,139 cm2 (Fig. 4), donde fue evidente el gran tamaño de las lesiones lo que se debe a una interacción compatible como reportan Vlesshouwers et al. (1999), quien afirma que la necrosis desarrollada por una interacción compatible es más rápida y de mayor tamaño.
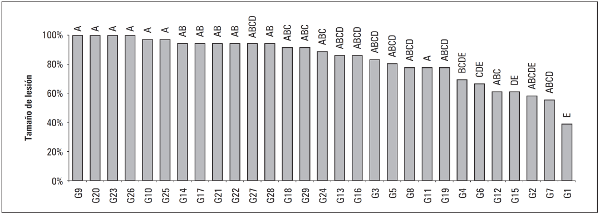
Figura 4 Prueba de significancia de medias LSD de Fisher (P≤0,005) para la variable tamaño de la lesión.
Resultados similares encontraron Reis et al. (2002) quienes estudiaron el estado y factores epidemiológicos sobre poblaciones de P. infestans sensu lato en Brasil, determinando que los componentes epidemiológicos son específicos de los huéspedes, dado que observaron valores significativos para los parámetros tamaño de lesión y eficiencia de infección. Además, Gabriel et al. (2011), al igual que Vlesshouwers et al. (1999), mencionan que se ha demostrado que los parámetros de resistencia como el tamaño de lesión pueden dar información valiosa sobre la resistencia genética de los genotipos evaluados.
Tasa de esporulación (TS)
El ANDEVA no mostró diferencias significativas para la variable tasa de esporulación entre los tratamientos (P>0,05), presentando desviaciones estándar bajas entre tratamientos (±0,1468) y entre repeticiones (±0,385), indicando que no existió variación entre los datos. Los datos de esta variable oscilaron entre 16.271,62 esporangios/cm2 y 61.448,87 esporangios/ cm2 para los genotipos G14 y G15, respectivamente. En general se presentó una tasa de esporulación promedio de 29.027,18 esporangios/cm2. El hecho de no encontrar diferencias entre las tasas de esporulación, se debe a que el ensayo estuvo sometido a condiciones de luz, temperatura, humedad, etc. semejantes y posiblemente no fueron las condiciones óptimas para la esporulación como las describe Castaño (1996) afirmando que las condiciones ideales para la producción de esporangios son humedad relativa alrededor del 80%, temperaturas que oscilan entre 16 y 22°C. Erwin y Ribeiro (1996) encontraron que el desarrollo y producción de esporangios se ve afectado por factores como son la humedad, la tensión de oxígeno, la luz, la temperatura del día tanto como de la noche y la nutrición. Sin embargo el micelio es capaz de vivir de forma saprofita sobre materia orgánica asegurando la invasión del tejido vegetal. Resultados similares encontraron Reis et al. (2002) en papa quienes afirman que los factores como la temperatura afecta los componentes epidemiológicos como la producción de esporangios.
Además, no se encontró una relación directa entre esporulación y el área de tejido afectado, los aislamientos que presentaron poca esporulación causaron un mayor daño al tejido, mientras que en áreas pequeñas de tejido afectado se registró una alta tasa de esporulación, resultados similares observaron Marín y Mira (1998) en estudios realizados en papa y pepino. Lo que se justifica con lo que proponen Agrios (2004), Niks y Lindhout (2004) y Sañudo y Betancourth (2005), quienes afirman que el contacto del patógeno con el hospedante no se limita solo a la reproducción de P. infestans sensu lato después de su establecimiento, sino a la cantidad de infección que produce con la presencia de daños o síntomas por unidad de patógeno presente.
Periodo de incubación (PI)
El periodo de incubación estuvo comprendido desde la hoja sin evidencia de lesiones seguido de la presencia de manchas necróticas de aproximadamente 4 cm2 al quinto día, hasta la necrosis casi total de las hojas en el octavo y décimo día (Fig. 5). Resultados similares reportan investigadores del Grupo Genética y Evolución de Organismos Tropicales (GENPAT) (2011) quienes observaron la presencia de síntomas visibles al cuarto y quinto día después de la infección en genotipos de tomate de árbol. Estos resultados muestran que la cepa más agresiva infecta la hoja rápidamente, causando necrosis en corto tiempo después del periodo de incubación e invaden casi completamente la hoja para el caso de los genotipos más susceptibles.
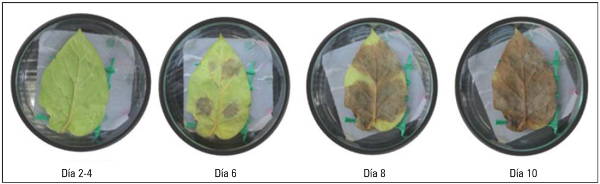
Figura 5 Desarrollo de la infección provocada por la cepa N9035 de Phytophthora infestans sensu lato sobre el genotipo G18 de C. betacea (Cav.) Sendt.
Además del buen comportamiento que reflejó el genotipo G1 este se destacó en otros estudios como el realizado por Benavides (2012) quien reporta un buen comportamiento agronómico y productivo como la variable número de frutos por racimo. Por otra parte, Valencia et al. (2011), afirman que este genotipo se destaca por su alto su porcentaje de ácido cítrico (3,64%) y buen tamaño de mesocarpo importante para el sector agroindustrial, ya que facilita la obtención de productos elaborados con valor agregado como mermeladas y conservas a costos reducidos.
CONCLUSIONES
Los genotipos presentaron diferentes comportamientos frente al ataque de la cepa N9035 de P. infestans sensu lato, a diferencia de la cepa P9153 con la cual los genotipos no difirieron entre sí. El genotipo G1 se identificó como un genotipo potencial para usarse en el mejoramiento de la especie por su tolerancia a P. infestans.

