



![Behavior of market prices and analysis of volatility for peach ( Prunus persica [L.] Batsch.) produced in Boyaca, Colombia](/img/en/prev.gif)







 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El tomate es una de las hortalizas más importantes en el mundo, debido al área sembrada, valor de la producción y nutricional, amplia distribución geográfica, elevado consumo, y sustento económico para pequeños y medianos productores. Esta hortaliza se ha establecido en todos los continentes del mundo. El área cosechada, de tomate, en el país durante el año 2014 alcanzó las 16.388 ha (FAOSTAT, 2014).
El tomate tipo “chonto”, es el cultivar de mayor producción y consumo en Colombia, se caracteriza por tener frutos bi o tri-loculares, peso promedio de fruto entre 80 y 180 g y formato de fruto ovalado. La totalidad de los cultivares nacionales poseen hábito de crecimiento indeterminado (Vallejo, 1999).
En Colombia, el tomate tipo “chonto” posee alta demanda para consumo directo y en menor proporción para uso industrial. A pesar de ser una de las especies vegetales más utilizadas y estudiadas, su cultivo está acompañado de múltiples problemas bióticos, abióticos y económicos. Dentro del aspecto económico se destacan el alto costo de producción generado, en gran parte, por el hábito de crecimiento indeterminado de la planta (Vallejo y Estrada, 2004), asociado a múltiples labores de tutorado, amarres, poda, manejo agronómico y aplicación de pesticidas, principalmente.
El crecimiento determinado en tomate está regulado por la expresión del gen recesivo self pruning (sp) en estado homocigoto (Vallejo, 1999; Krieger et al., 2010), y el crecimiento indeterminado por el mismo gen en estado dominante (Sp_) bien sea homocigoto y heterocigoto, otorgando un porte alto de planta, ciclos fenológicos prolongados y un crecimiento indefinido, debido a la terminación de la planta en un meristemo vegetativo.
En el crecimiento determinado (spsp), la planta posee porte bajo y crecimiento definido debido a la terminación de la planta en un meristemo apical reproductivo (Pnueli et al., 1998; Fridman et al., 2002). Se ha reportado un tercer tipo de crecimiento, gobernado por la expresión del gen SPD9, presente en la especie silvestre Solanum penellii, que atribuye a la planta crecimiento semideterminado y alto contenido de sólidos solubles (Carmel-Goren et al., 2003).
Para la introducción de características monogénicas en tomate, como el hábito de crecimiento determinado, se ha utilizado la metodología del retrocruzamiento debido a la efectividad para recuperar las características favorables del padre recurrente (Vallejo et al., 2010).
Las ventajas de los cultivares de tomate con hábito de crecimiento determinado han justificado una serie de estudios tendientes a la modificación de la arquitectura de la planta de los cultivares indeterminados actuales (Vallejo y Estrada, 2002). Por lo anterior, el objetivo del presente estudio fue producir, evaluar y seleccionar líneas de tomate tipo “chonto” con hábito de crecimiento determinado y características propias de los cultivares requeridos por el mercado nacional.
MATERIALES Y MÉTODOS
Ubicación
El estudio se realizó en el Centro Experimental de la Universidad Nacional de Colombia, sede Palmira (CEUNP), ubicado en el municipio de Candelaria, Valle del Cauca, corregimiento El Carmelo a 3º24’ N y 76º26’ W, a 980 msnm, se categoriza dentro de una zona cálida-moderada y semiárida, 24ºC de temperatura promedia, 69% humedad relativa y 1.009 mm de precipitación promedia anual (Pardey, 2008). La cuantificación del contenido de sólidos solubles se llevó a cabo en el Laboratorio de Fisiología Vegetal de la Universidad Nacional de Colombia, Palmira.
Material vegetal
Se utilizaron nueve líneas de tomate tipo “chonto”, procedentes del retrocruzamiento tres (RC3) entre el cultivar brasilero IPA 4 que fue el progenitor donante del gen sp y el cultivar colombiano (progenitor recurrente) UNAPAL-Maravilla:
1) L1-RC3 (IPA 4 x UNAPAL-Maravilla)
2) L2-RC3 (IPA 4 x UNAPAL-Maravilla)
3) L3-RC3 (IPA 4 x UNAPAL-Maravilla)
4) L4-RC3 (IPA 4 x UNAPAL-Maravilla)
5) L5-RC3 (IPA 4 x UNAPAL-Maravilla)
6) L6-RC3 (IPA 4 x UNAPAL-Maravilla)
7) L7-RC3 (IPA 4 x UNAPAL-Maravilla)
8) L8-RC3 (IPA 4 x UNAPAL-Maravilla)
9) L9-RC3 (IPA 4 x UNAPAL-Maravilla)
El parental recurrente utilizado para realizar el cuarto retrocruzamiento (RC4), correspondió al cultivar comercial de crecimiento indeterminado, UNAPAL-Maravilla.
Trabajo de campo
Se desarrolló en cuatro fases:
Fase 1: obtención de la semilla F1 del retrocruzamiento cuatro (F1RC4). Cada una de las nueve líneas procedentes del retrocruzamiento tres (RC3), usadas como progenitores maternos donantes, se cruzaron con el cultivar de crecimiento indeterminado UNAPAL- Maravilla que hizo la función de progenitor recurrente. Se utilizaron parcelas de 10 plantas por cada progenitor.
Fase 2: obtención de la semilla F2 del retrocruzamiento cuatro (F2RC4). La semilla F1 del retrocruzamiento cuatro (F1RC4) se sembró y se dejó a libre polinización con el fin de que se presentara la autofecundación natural, debida a su condición autógama. Se utilizaron parcelas de cinco plantas para cada F1RC4.
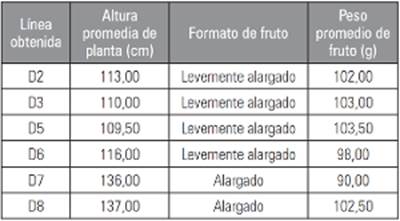
Fase 3: selección de plantas con crecimiento determinado (spsp). Se sembraron 40 plantas F2RC4 de cada uno de los retrocruzamientos con el fin de observar la segregación genética y proceder a la selección de plantas con crecimiento determinado, altura menor a 150 cm, peso de fruto mayor a 90 g y formato de fruto ovalado.
Fase 4: evaluación de líneas seleccionadas, con crecimiento determinado. Se usó el diseño de Bloques Completos al Azar (BCA), con cuatro repeticiones. Como testigo se utilizó el progenitor recurrente de crecimiento indeterminado UNAPAL-Maravilla. Se sembraron nueve plantas por parcela y se evaluaron las seis plantas centrales (parcela útil).
Evaluación de caracteres
Se evaluaron los siguientes caracteres relacionados con la fenología, morfología, producción y calidad de fruto:
Caracteres fenológicos
Días a inicio de floración (DAF) |
cuando el 60% de las plantas de la parcela útil presentaron la primera flor abierta. |
Días a inicio de cosecha (DAC) |
cuando el 60% de las plantas de la parcela útil iniciaron el periodo de cosecha. |
Duración de cosecha (DUC) |
diferencia entre el inicio y el final de la cosecha, para cada una de las plantas de la parcela útil. |
Ciclo productivo (CP) |
días trascurridos entre la siembra de plántulas a campo y el final de la cosecha. |
Caracteres morfológicos
Altura final de planta (AFP) |
promedio de altura de planta, expresado en centímetros, de las seis plantas de la parcela útil, al finalizar la cosecha. |
Distancia de entrenudos (DE) |
promedio, expresado en centímetros, medido en el tallo principal de las seis plantas de la parcela útil. |
Diámetro de tallo (DT) |
promedio del diámetro, en milímetros, del tallo principal de las seis plantas de la parcela útil. |
Caracteres productivos
Producción por planta (PP) |
producto del peso promedio de fruto por el número de frutos por planta. |
Número de frutos por planta (NFP) |
suma de todos los frutos producidos por planta dentro de la parcela útil. |
Peso promedio de fruto (PPF) |
relación entre el peso total de frutos y el número de frutos cosechados por planta, en la parcela útil. |
Número de racimos por planta (NRP) |
suma de todos los racimos producidos por planta dentro de la parcela útil. |
Rendimiento (R) |
proyección de la producción por planta a una hectárea (ha), teniendo en cuenta una densidad poblacional de 20.833 plantas por hectárea, se expresó en toneladas/ha. |
RESULTADOS Y DISCUSIÓN
Fases 1 y 2: retrocruzamiento y autofecundación
Se obtuvieron seis híbridos F1RC4, como producto del cuarto retrocruzamiento, lo cual permitió recuperar, teóricamente, el 96,87% del progenitor recurrente de acuerdo a lo mencionado por Vallejo y Estrada (2002). Estos híbridos se sometieron a autofecundación natural con el fin de obtener las poblaciones segregantes F2RC4. En esta fase, la presencia de plagas y altas temperaturas redujo la eficiencia de los cruzamientos dirigidos y la formación de frutos. Lo anterior fue reportado por Filgueira (1982) al encontrar aborto floral, como efecto negativo de temperaturas superiores a 35ºC e insectos plaga sobre el desarrollo normal de flores y frutos, en esta especie.
Fase 3: selección de líneas con crecimiento determinado a partir de poblaciones F2RC4
La segregación de la generación F2RC4 permitió identificar plantas con hábito de crecimiento determinado. La población total fue de 240 individuos sembrados, en la que 40 plantas fueron utilizadas por cada genotipo, de esta población total se seleccionaron nuevamente seis líneas tipo “chonto” con crecimiento determinado. Estas líneas se obtuvieron a través de selección fenotípica de plantas individuales o mezcla de dos individuos con características similares. Estas líneas se denominaron con la letra D (determinado) y el número de la línea que las generó o la secuencia numérica producto del orden de las selecciones; se escogieron por valores promedio de los criterios de selección, en los individuos analizados (Tab. 1).
Fase 4: evaluación de líneas seleccionadas
En las Tablas 2 y 3 se presentan los cuadrados medios de todos los caracteres evaluados. La fuente de variación genotipo, conformada por las seis líneas seleccionadas y el testigo UNAPAL-Maravilla, presentó diferencia estadística para la mayoría de las variables fenológicas, morfológicas, productivas, forma y calidad de frutos, a excepción de los caracteres diámetro de tallo (DT) y relación diámetro-longitud de fruto (DT). Lo anterior indicó que dentro de los genotipos evaluados, al menos un genotipo presentó diferencias para estos caracteres.
Tabla 2 Cuadrados medios del análisis de varianza para algunas variables fenológicas y morfológicas de plantas de tomate tipo “chonto”.
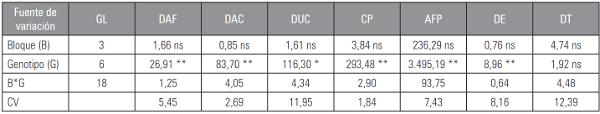
CV: coeficiente de variación: GL: grados de libertad; * significancia estadística (P≤0,05); ** significancia estadística (P≤0,01); ns: no significancia.
DAF: día a inicio de floración; DSC: días a inicio de cosecha; DUC: duración de cosecha; CP: ciclo productivo; AFP: altura final de planta; DE: distancia de entrenudos; DT: diámetro de tallo.
Tabla 3 Cuadrados medios del análisis de varianza para caracteres relacionados con producción, forma y calidad de fruto, en plantas de tomate tipo “chonto”.
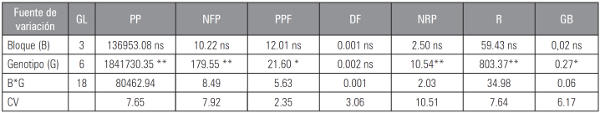
CV: coeficiente de variación: GL: grados de libertad; * significancia estadística (P≤0,05); ** significancia estadística (P≤0,01); ns: no significancia.
PP: producción por planta; NFP: número de frutos por planta; PPF: peso promedio de fruto; DF: relación longitud y diámetro de fruto; NRP: número de racimos por planta; R: rendimiento; GB: contenido de sólidos solubles (grados Brix).
En la figura 1 se presentan los promedios de los caracteres fenológicos, mientras que en la Tabla 4, se presentan los promedios de algunos caracteres morfológicos, productivos y de calidad de fruto.
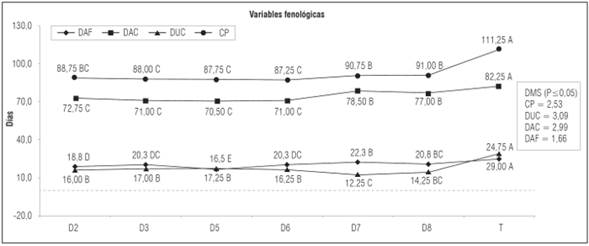
Figura 1 Promedios de algunas variables fenológicas de las seis líneas de tomate tipo “chonto” obtenidas en la generación F2. Promedios con letras distintas indican diferencia significativa según la prueba DMS (P≤0,05).
Tabla 4 Promedios de algunas variables morfológicas, productivas y de calidad de fruto de las seis líneas de tomate tipo “chonto” obtenidas en las poblaciones segregantes F2RC4.
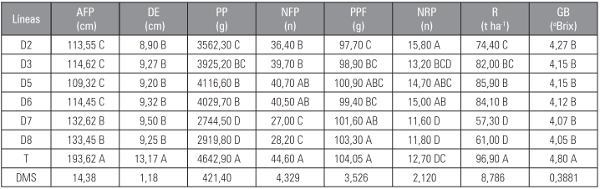
Promedios con letras distintas indican diferencia significativa según la prueba DMS (P≤0,05).
AFP: altura final de planta; DE: distancia de entrenudos; PP: producción por planta; NFP: número de frutos por planta; PPF: peso promedio de fruto; DF: relación longitud y diámetro de fruto; NRP: número de racimos por planta; R: rendimiento; GB: contenido de sólidos solubles (grados Brix).
Caracteres fenológicos
El comportamiento fenológico de las líneas con crecimiento determinado fue diferente al del testigo comercial de crecimiento indeterminado. Para el parámetro DAF se encontró diferencias significativas. La línea D5 mostró mayor precocidad, 16,50 d, para inicio de floración en comparación con el testigo que alcanzó 24,75 d. Las líneas D2, D3 y D6 presentaron una reducción en el tiempo a inicio de floración del 24,20; 18,18 y 18,18%, respectivamente.
En el estudio de la fenología de la floración de tomate, Piotto y Peres (2012) reportaron reducciones entre el 10,00 y 15,00% para plantas con hábito de crecimiento determinado; sin embargo, en esta investigación, al comparar el genotipo de mayor precocidad, D5, con el testigo se presentó una mayor reducción con el 33,30%; por otro lado, la línea de crecimiento determinado con el periodo más largo para iniciar floración, D7, presentó una reducción del 10.10% lo cual concuerda con lo reportado por dichos autores.
Lo anterior permite inferir que existe un importante potencial en cuanto a la precocidad para iniciar floración en las líneas evaluadas. El valor encontrado en el testigo se asemeja al reporte de Vallejo y Estrada (2004), en el que se menciona que los DAF para este tipo de cultivares es aproximadamente de 30 d.
Para el parámetro DIC, la línea D5 presentó la mayor precocidad con 70.50 días; sin embargo fue estadísticamente igual a las líneas D2, D3, D5 y D6.
El genotipo con crecimiento de tipo indeterminado presentó el periodo más tardío para inicio de la cosecha y alcanzó 82,25 d. El comportamiento de las líneas con crecimiento determinado presentó un valor inferior al encontrado por Santiago et al. (1998) para este parámetro, en el que la variedad determinada Río Grande, tipo “saladete”, inició la cosecha a los 75,00 d; a excepción de las líneas D7 y D8 que presentaron valores cercanos con 77,00 y 78,50, respectivamente.
Nuez (1995) reportó la gran diferencia en cuanto a precocidad para las fases fenológicas entre cultivares de crecimiento determinado e indeterminado en tomate, siendo las de plantas de porte bajo, las que presentan fases más cortas.
La DUC fue menor en las líneas con hábito de crecimiento determinado. Los resultados encontrados mostraron que el tomate tipo “chonto” con hábito de crecimiento indeterminado necesitó mayor tiempo para la maduración total de los frutos. El testigo alcanzó la mayor duración de cosecha con 29,00 d; sin embargo esta fase pudo haberse prolongado, por la susceptibilidad de UNAPAL-Maravilla a insectos plaga como Prodiplosis longifila Gagné. y Neoleucinodes elegantalis Guenée, que dificultó la continuación del proceso de floración y maduración de frutos. Esto lo menciona Vallejo (2004) en sus investigaciones realizadas para conocer la resistencia a las principales plagas de esta variedad. Estos resultados se ajustaron a los encontrados por Vallejo et al. (1994) para tomate de tipo “chonto” en los que el rango para duración de cosecha estuvo entre 18,00 y 29,00 d.
Por otro lado, las líneas con hábito de crecimiento determinado presentaron periodos de cosecha que oscilaron entre 12,20 y 17,20 d, siendo más precoces, para los estándares de tomate tipo “chonto” con crecimiento indeterminado, reportados por Vallejo et al. (1994). Lo anterior es corroborado también por estudios en los que se destaca, la concentración de la cosecha, como una de las principales ventajas del tipo de crecimiento determinado, ya que la maduración es más homogénea (Vallejo, 1999).
En la medición del parámetro CP de la planta, el testigo UNAPAL-Maravilla, presentó el ciclo de cultivo más largo con 111,25 d y las líneas con crecimiento determinado D6, D5, D3 y D2, terminaron su ciclo total en 87,20; 87,70; 88,00 y 88,70 d, respectivamente. Al comparar la línea D6, que presentó el menor valor para esta característica, con el testigo que alcanzó 111,20 d, se obtuvo una reducción del 21,47% en el tiempo total del ciclo.
La obtención de cultivares con ciclos cortos de producción permite reducir la exposición de las plantas a condiciones ambientales, que pueden ser adversas, como resultado de modificaciones provocadas por el cambio climático; además disminuir la cantidad de aplicaciones químicas para control fitosanitario, lo que favorece el factor ecológico y económico del cultivo. Al tener cultivares precoces y de porte bajo, la ejecución de algunas labores puede reducirse, tal como mencionan Vallejo y Estrada (2004); plantas de menor altura y menor ciclo productivo, requieren menor cantidad de insumos y materiales.
Por otra parte, Nuez (1995) reporta que el cultivo de variedades de tomate de crecimiento determinado promueve el uso eficiente e intensivo del suelo, al permitir mayor cantidad de siembras al año, lo que genera mayor productividad.
Caracteres morfológicos
Se encontraron diferencias significativas para los caracteres distancia de entrenudos (DE) y altura final de planta (AFP). La distancia de entrenudos presentó diferencias significativas entre los genotipos con hábito de crecimiento contrastantes, el testigo presentó plantas con mayor distancia para esta estructura, con 13,17 cm; las líneas de crecimiento determinado presentaron un comportamiento estadísticamente igual entre ellas, oscilando entre 8,90 y 9,50 cm. Lo anterior es corroborado con los resultados de Filgueira (1982) cuando mencionó que uno de los efectos de la expresión del gen sp en estado homocigoto, es la reducción de la distancia de entrenudos. Además, la reducción de la distancia de entrenudos resulta una característica importante por su relación con la altura de la planta y mayor concentración de la cosecha.
El testigo UNAPAL-Maravilla alcanzó la mayor altura final de planta (AFP) con 193,60 cm al terminar su ciclo productivo. En los genotipos de crecimiento determinado se encontró que la línea D5 presentó la menor altura con 109,32 cm. Al comparar esta línea con el testigo se presentó una reducción del 43,53% en la AFP, alcanzada al terminar el ciclo productivo. Lo anterior concuerda con lo reportado por Vallejo (1999), quien mencionó que los genotipos de tomate con hábito de crecimiento indeterminado pueden presentar alturas mayores a 2,00 m.
Caracteres de producción, forma y calidad de fruto
Los caracteres producción por planta (PP), número de frutos por planta (NFP), número de racimos por planta (NRP) y rendimiento (R) presentaron diferencias altamente significativas; mientras que los caracteres peso promedio de fruto (PPF) y contenido de sólidos solubles (GB) presentaron diferencias significativas.
El testigo de crecimiento indeterminado, presentó la mayor producción (PP) con 4.642,90 g por planta, diferenciándose estadísticamente de las líneas con crecimiento determinado. Dentro de las líneas evaluadas spsp, el valor más alto fue alcanzado por los genotipos D5, D6 y D3 con 4.116,60, 4.029,70 y 3.925,20 g por planta, respectivamente, sin diferencia estadística entre ellas. Estos resultados concuerdan con los reportados por Baena et al. (2003) quien encontró valores de rendimiento, en tomate “chonto”, que oscilaron entre 3.328,00 y 5.904,00 g por planta.
El NFP presentó un comportamiento estadísticamente similar, entre el testigo UNAPAL-Maravilla y las líneas con crecimiento determinado. Las líneas D5 y D6 produjeron 40,70 y 40,50 frutos por planta, respectivamente; mientras que el testigo alcanzó 44,62 frutos. El número de frutos del testigo concordó con lo reportado por Moya (2005), quien encontró valores entre 37,00 y 89,00, para la variable frutos por planta, en cultivares con crecimiento indeterminado.
Las líneas con crecimiento determinado presentaron mayor cantidad de racimos por planta (NRP), comparadas con el testigo que presentó 12,70 racimos. Las líneas D2, D6 y D5 fueron las de mejor comportamiento, con 15,80; 15,00 y 14,70 racimos, respectivamente y estadísticamente similares. La expresión fenotípica de estas líneas, en cuanto a esta variable, resultó promisoria.
El rendimiento proyectado por hectárea, permitió observar que el testigo, con 96,97 t ha-1, fue superior a las líneas de mejor comportamiento D5, D6 y D2, con 85,97; 84,10 y 74,40 t ha-1, respectivamente. Lo anterior concuerda con la investigación realizada por Carvalho et al. (2003), en la que se encontraron algunos rendimientos proyectados de 85,00; 95,10 y 96,50 t ha-1, para este tipo de cultivares de crecimiento indeterminado. Se evidenció el buen comportamiento de las líneas de crecimiento determinado, puesto que obtuvieron rendimientos cercanos a los producidos por cultivares de crecimiento indeterminado y superiores a los reportados por Ledo et al. (1994), para líneas de crecimiento determinado, con 18,40 y 53,30 t ha-1.
El carácter PPF presentó un comportamiento estadísticamente similar entre el testigo y las líneas D8, D7, D5. El mayor peso de fruto fue alcanzado por el testigo con 104,05 g y la línea D8 con 103,37 g. Estos resultados coincidieron con los encontrados por Baena et al. (2003), quien obtuvo pesos entre 95,00 y 124,00 g para frutos de tomate tipo “chonto”.
El contenido de sólidos solubles (GB), osciló entre 4,05 y 4,27 ºBrix, en las seis líneas con crecimiento determinado. El cultivar con crecimiento indeterminado presentó mayor valor con 4,80 ºBrix. Los valores encontrados para esta variable estuvieron dentro del rango reportado por Carvalho et al. (2003) con valores entre 3,70 y 5,01 ºBrix, para estos cultivares. Por otra parte, Yelle et al. (1991) reportaron que los frutos provenientes de plantas con crecimiento determinado presentaron un menor contenido de sólidos solubles que los frutos de plantas con hábito de crecimiento indeterminado.
Lo anterior puede deberse a que el contenido de sólidos solubles está relacionado, principalmente, con la acumulación de azúcares; según Yelle et al. (1991), este proceso está influenciado por factores ambientales y dentro de la planta por procesos fisiológicos, como la asimilación de CO2 a través del tejido foliar y la movilización de azúcares por medio del floema hasta su almacenamiento en el fruto. De esta manera, el hábito de crecimiento determinado podría afectar el contenido de sólidos solubles debido a la reducción en el número de hojas o área foliar; sin embargo es necesario realizar más investigaciones en este tema y su relación con el hábito de crecimiento. Esta inferencia tiene en cuenta los reportes de Nuez (1995), quien evidencia la reducción del número de hojas en plantas de porte bajo.
CONCLUSIONES
El método de retrocruzamiento permitió obtener híbridos F1RC4 con la presencia del gen sp, responsable del crecimiento determinado, y con una recuperación teórica del 96,875%, de las características del progenitor recurrente.
En la generación segregante F2 de retrocruzamiento cuatro (F2RC4) se obtuvieron seis líneas promisorias con crecimiento determinado, peso y formato de fruto propios de tomate tipo “chonto”.
Se seleccionaron las líneas D5 y D6, tipo “chonto,” por presentar crecimiento determinado; buen potencial productivo; peso promedio de fruto mayor a 100,00 g, precocidad a cosecha, ciclo de cultivo corto, frutos con formato ovalado y grados Brix superiores a 4,00.

