











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La gulupa (Passiflora edulis f. edulis Sims) es un frutal perenne de crecimiento indeterminado. En Colombia, es la segunda especie de importancia económica dentro del género Passiflora por su alta demanda en mercados internacionales. Puede consumirse en fresco y procesada, caracterizándose por su contenido nutricional, sabor y aroma (Ocampo et al., 2007; Jiménez et al., 2011).
El seguimiento al desarrollo fenológico en etapa reproductiva y productiva de frutos, permite construir curvas de crecimiento y desarrollo, las cuales son útiles para optimizar las prácticas y planes de manejo del cultivo. Recientemente el estudio de la fenología de especies frutales fue enfocado a la respuesta frente a variaciones climáticas (Yaacoubi et al., 2014); sin embargo, algunos autores consideran que el aumento en los recursos nutricionales también puede influir en el desarrollo fenológico de las especies (Chmielewski, 2013). Se ha reportado que la producción de algunos frutales se ve comprometida por la pérdida de flores y frutos durante el cuajamiento, las cuales están relacionadas con factores bióticos y abióticos, actividad hormonal y estado nutricional de la planta (Mengel et al., 2001; Lahav and Whiley, 2002). En algunas especies frutales se ha demostrado que la deficiencia de algunos micronutrientes, principalmente del boro (B), afectan la floración y fructificación (Muhammad et al., 2015).
El B desempeña un papel estructural como componente del complejo Rhamnogalacturonano II (RGII) que une polisacáridos de la pared celular y la lámina media (Kobayashi et al., 2011; Reid, 2004), ocasionando que la deficiencia de este elemento presente efectos anatómicos, fisiológicos y bioquímicos, como daños en la expansión celular e integridad de membranas, afectando el crecimiento y desarrollo de las plantas (Mei et al., 2016).
El B también se ha reportado en procesos como el desarrollo floral, en varios cultivos. La deficiencia de este elemento causa limitada producción de flores, baja producción y viabilidad de polen, infertilidad de flores y caída prematura de flores y frutos (Nymora y Brown, 1997; Perica et al., 2001). Se considera que esos efectos pueden ser observados en ausencia de síntomas vegetativos o reducción del crecimiento, lo que indica que el requerimiento de B en los tejidos reproductivos es mayor que en los tejidos vegetativos, o que la liberación del B a los tejidos reproductivos es limitada (Iwai, 2006).
Estudios recientes han reportado que el bajo porcentaje de cuajado de fruto de algunas especies frutales está asociado a pistilos subdesarrollados, producción de flores más pequeñas y poca viabilidad del polen. Estos fenómenos están relacionados con bajos contenidos de almidón y translocación de carbohidratos a estas estructuras, lo cual a su vez está relacionado con deficiencias nutricionales asociadas con B en la etapa de floración y fructificación. Lo anterior ha sido bien documentado para las especies Prunus armeniaca (Rodrigo et al., 2000), Persea americana (Boldingh et al., 2016) y Punicagranatum (Davarpanah et al., 2016). Así mismo, se ha demostrado que la aplicación foliar de B genera un efecto positivo en la germinación del polen y en el crecimiento del tubo polínico, aumentando así el porcentaje de cuajado de fruto (Sarkar et al., 2007).
Pocos reportes se encuentran acerca del efecto de la aplicación de B en especies de Passiflora en etapa productiva. Freitas et al. (2006, 2011) indican que en Pas-siflora edulis Sims f. flavicarpa bajo deficiencia de B se presentan flores secas, con deformaciones, número y tamaño anormal de estilos y estigmas, reducción en el número de frutos por planta y menor peso de los frutos.
Para el caso de P. edulis Sims f. edulis se ha estudiado la fenología reproductiva y el crecimiento del fruto en diferentes zonas bajo condiciones ambientales (Flórez et al., 2012); sin embargo, no se ha documentado el efecto de la fertilización foliar con B en el crecimiento y desarrollo de estructuras reproductivas y el cuajado de frutos.
Teniendo como base la importancia que tiene el B en especies frutales, sumado al hecho que existe poca información acerca del efecto de su aplicación foliar en plantas de P. edulis Sims f. edulis y de la relación del micronutriente con el desarrollo fenológico en etapa demandante, el objetivo de esta investigación fue evaluar el efecto de la aplicación foliar de B en el desarrollo fenológico durante la etapa productiva y cuajado de frutos de P. edulis Sims f. edulis bajo condiciones de campo.
MATERIALES Y MÉTODOS
Material vegetal y condiciones de crecimiento
Las plantas de gulupa (Passiflora edulis f. edulis Sims) se establecieron en el municipio de Pasca (Cundinamarca, Colombia), en la vereda San Pablo, en la finca Bellavista, ubicada a 4°18'45,52'' N y 74°19'58,13" W a una altitud de 2.006 msnm. Se sembraron plántulas de gulupa obtenidas a partir de semilla, a una densidad de siembra de 816 plantas/ha, a una distancia de siembra de 3,5 m entre surcos y 3,5 m entre plantas. Se empleó un sistema de tutorado de espaldera sencilla a doble alambre, uno ubicado a 1,5 m y el otro a 2 m del nivel del suelo. Para dar arquitectura a las plantas, se dejó una rama principal y posteriormente se dejaron dos ramas laterales que fueron guiadas sobre el alambre del tutorado, estas se podaron cuando alcanzaron una longitud aproximada de 1,75 m con el objetivo de estimular la formación de ramas secundarias. Para el segundo ciclo productivo, posterior a la cosecha de frutos de cada rama productiva, se realizó la poda sobre cada rama dejando dos yemas con el objetivo de dar formación a las nuevas ramas productivas.
El suelo presentó pH de 4,5, por lo que fue necesario antes de la siembra aplicar 100 g de cal dolomita por planta. Las características físico-químicas del suelo fueron determinadas en el laboratorio de la Universidad ECCI, la extracción de B en el suelo se realizó por el método de Beger y Troug y la determinación por absorción atómica (Tab. 1). Se realizó fertilización edáfica fraccionada, desde el momento de siembra de las plantas hasta producción aplicando en total 132,76 kg ha-1 de N, 71,16 kg ha-1 de P2O5, 142,56 kg ha-1 de K2O, 61,20 kg ha-1 de CaO y 27,64 kg ha-1 de MgO. El fraccionamiento consistió en cinco aplicaciones durante el período vegetativo y el primer ciclo productivo cada 45 d, y dos aplicaciones durante el segundo ciclo, la primera 15 d después de la poda de inducción de ramas y la segunda en el día 45.
Tabla 1 Características físico-químicas del suelo donde se estableció el cultivo de gulupa, vereda San Pablo, finca Bella Vista a 4°18'45,52" N y 79°19'58,13 W en Pasca, Cundinamarca.
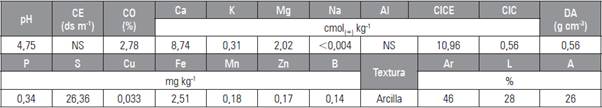
CE, conductividad eléctrica; CO, carbono orgánico; CICE, capacidad de intercambio catiónico efectiva; CIC, capacidad de intercambio catiónico; DA, densidad aparente; Ar, arcilla; L, limo; A, arena.
Se instaló una estación meteorológica (Coltein Ltda, Bogotá, Colombia) con dataloggers (Coltein Ltda., Bogotá y Hobo U12-006, Onset Computer Corporation, Bourne, MA) para el registro de temperatura, humedad relativa y radiación fotosintéticamente activa (PAR) y se registraron datos cada 15 min. Las características climáticas presentadas durante el primer ciclo productivo fueron: temperatura media del aire 18,4°C, humedad relativa del aire 85,0%, radiación fotosintéticamente activa de 365,2 /µmol de fotones m-2 s-1 y déficit de presión de vapor (DVP) de 0,60 kPa. Para el segundo ciclo productivo se presentó una temperatura media del aire 18,1°C, humedad relativa del aire 79,9%, radiación fotosintéticamente activa de 382,2µmol de fotones m-2 s-1 y DVP de 0,74 kPa.
Tratamientos foliares con boro
Los tratamientos de aplicación foliar con B fueron: T0, 0 kg ha-1; T50, 0,3 kg ha-1; T100, 0,6 kg ha-1 y T150, 0,9 kg ha-1 y se realizaron con base en el reporte de extracción de este nutriente por Haag et al. (1973) para P. edulis Sims f. flavicarpa. Para el presente trabajo dicho valor de extracción se tomó como el 50% debido a que por el método de aplicación foliar se considera que solo el 50% de la solución aplicada es absorbida por la planta y que depende también de la velocidad de absorción del elemento; para algunos elementos de baja movilidad como el B pueden tener una velocidad de absorción foliar entre 1 a 20 días (Meléndez y Molina, 2001).
En cada uno de los ciclos productivos (desde el momento de formación de botones florales) las aplicaciones de B se distribuyeron en cuatro momentos cada 7 d, asperjando a la planta la concentración del respectivo tratamiento en un volumen de solución final de 150 mL; adicionalmente se realizó una quinta aplicación inmediatamente se observó cuajado de fruto. En cada uno de los ciclos productivos, el inicio de la aplicación del tratamiento fue cuando la mayoría de las ramas terciaras presentaron emergencia de botones florales, con un tamaño de botón entre 0,5 cm de ancho y 1 cm de largo. Para el primer ciclo productivo las aplicaciones iniciaron a los 178 días después de trasplante (ddt) equivalente al estado de formación de botones florales. Para el segundo ciclo se realizó después de la poda de producción aproximadamente a 390 ddt). La aplicación de B se realizó vía foliar utilizando como fuente ácido bórico (H3BO3). Para la aspersión se usó un equipo manual de espalda Royal Condor® (Bogotá) ref. CO-004®, con una capacidad de 20 L, se usó una boquilla de tipo abanico sólido, con un caudal de aspersión de 600 cm3 min-1, adicionalmente se usó como coadyuvante y corrector de pH solución Agrotin® (Bayer Crop Science).
Desarrollo de ramas productivas
Las mediciones iniciaron desde el momento en que se aplicó el primer tratamiento foliar. Se seleccionaron cuatro plantas por tratamiento marcando cuatro ramas productivas de 4 cm. Se registró semanalmente la longitud de ramas y el número de entrenudos de cada rama hasta alcanzar una longitud de 115 cm para el primer ciclo productivo, y 130 cm para el segundo ciclo productivo; estas longitudes corresponden al momento en el cual la rama está lista para ser podada después del descargue de frutos. Para el segundo ciclo productivo, después de realizar la poda de ramas productivas con solo dos yemas, se marcaron la misma cantidad de ramas para cada tratamiento y se continuó con el registro.
Emisión de botones florales y frutos
Se seleccionaron cuatro ramas productivas por planta y tres plantas por tratamiento. Después de la primera aplicación de B, se realizó cada 7 d el conteo de cada botón floral emergido y los frutos cuajados sobre la rama productiva marcada.
Cuajado de fruto
En cada ciclo productivo se seleccionaron tres ramas productivas por planta, dentro de estas se marcaron cuatro botones florales de aproximadamente 0,5 cm de largo. Se tomaron tres plantas por tratamiento, para un total de 36 botones por tratamiento. Semanalmente se monitoreó la aparición de frutos cuajados; siguiendo la escala BBCH generada en el presente trabajo (resultados no mostrados) con base en la escala descrita para Passiflora ligularis Juss (Rodríguez-León et al., 2015). El código BBCH 69 corresponde al cuajado del fruto el cual ocurre a los 30 días después de la aparición del botón floral. El porcentaje de cuajado de fruto se determinó para cada tratamiento como la relación entre número de frutos formados y número de botones marcados.
Desarrollo del fruto
Para evaluar el desarrollo de fruto se marcaron por planta nueve frutos recién cuajados de 1,5 cm de longitud (calibrador digital Fisher Scientific, 0-150 mm), se tomaron tres plantas por tratamiento. En cada fruto marcado se registró el diámetro ecuatorial semanalmente (cada 7 d) hasta que los frutos llegaron a la madurez de cosecha (87 ddt) para el primer ciclo productivo. Para el segundo ciclo la evaluación se realizó hasta madurez comercial (143 ddt).
Análisis estadístico
Se estableció un diseño completamente aleatorio con submuestreo, se validaron los supuestos de normalidad y homogeneidad de varianzas para todas las variables, de acuerdo con las pruebas estadísticas de Kolmogorov-Smirnov (Normalidad) y Levene (Homogeneidad de varianzas), se realizaron análisis de varianza (ANOVA) de una vía y pruebas de comparación de medias de Tukey y Student (P≤0,05) para determinar diferencias entre tratamientos. Para las curvas de crecimiento se ajustó al modelo matemático de Gompertz de tres parámetros (a, b y c) con el software estadístico SPSS v.20.0 (IBM Corporation, Armonk, Nueva York). La parametrización de este modelo se realizó según lo indicado por Winsor (1932).
RESULTADOS
Desarrollo de ramas productivas
La elongación de ramas secundarias en el primer ciclo productivo (Fig. 1A) presentó el mismo comportamiento para todos los tratamientos sin diferencias significativas con la prueba Tukey (P>0,05); sin embargo, la longitud final de las ramas varió entre tratamientos, siendo mayor en T50 (115,72 cm) y menor en T150 (86,83 cm), indicando que las plantas de T50 requieren menos tiempo (94 ddt) para alcanzar la máxima longitud. En el segundo ciclo productivo (Fig. 1B) no se presentaron diferencias significativas con la prueba Tukey (P>0,05) entre tratamientos en los primeros 28 ddt; posteriormente el T50 mostró una elongación de ramas más rápida comparado con los demás tratamientos, alcanzando la máxima longitud (130,7 cm) a los 119 ddt, sin mostrar diferencias significativas con T100. Los tratamientos T0 y T150 presentaron una elongación de ramas más lenta, alcanzando la longitud máxima a los 143 ddt (130,5 y 129,87 cm, respectivamente) (Fig. 1B). De acuerdo con los resultados del modelo de crecimiento de Gompertz para esta variable, se encontró un mayor ajuste para el segundo ciclo (R 2>0,91) comparado con el primero (R 2>0,62) en todos los tratamientos. Las tasas de crecimiento obtenidas con el modelo para el primer ciclo fueron mayores en los tratamientos T50 y T100 (1,23, 1,19 cm d-1, respectivamente) y menores en T0 y T150 (0,96 y 0,92 cm d-1, respectivamente). Los resultados en el segundo ciclo muestran un comportamiento similar (0,91, 1,09, 1,08 y 0,90 cm d-1 para los tratamientos T0, T50, T100 y T150, respectivamente).
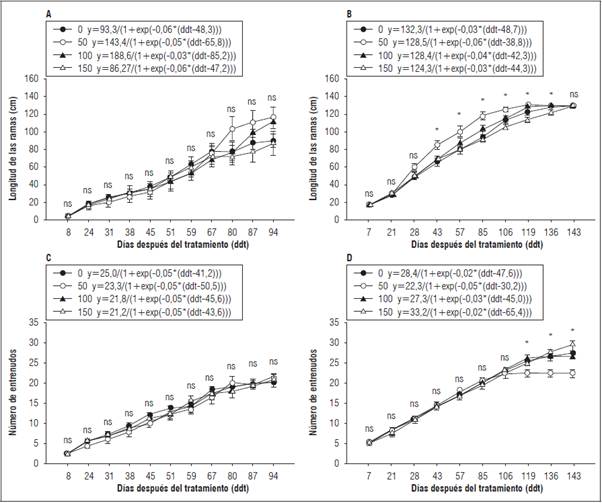
Figura 1 Desarrollo de ramas y entrenudos en plantas de gulupa (Passiflora edulis f. edulis Sims) fertilizadas con diferentes dosis de boro foliar aplicado en la formación del botón floral y cuajado de fruto en dos ciclos productivos (T0: 0 kg B ha1; T50: 0,3 kg B ha1; T100: 0,6 kg B ha-1 y T150: 0,9 kg B ha1). Longitud de ramas en el primer (A) y segundo ciclo (B); número de entrenudos en el primer (C) y segundo ciclo (D) de la producción. Las barras verticales corresponden al error estándar n=9. Se muestra la ecuación del modelo Gompertz para cada uno de los tratamientos. * indica diferencias significativas entre tratamientos de acuerdo con la prueba de Tukey (P≤0,05). ns: sin diferencias significativa.
En el primer ciclo productivo, el número de entrenudos no presentó diferencias significativas con la prueba de Tukey (P>0,05) entre tratamientos (Fig. 1C). En el segundo ciclo productivo no se encontraron diferencias significativas con la prueba Tukey (P>0,05) hasta los 106 ddt, tiempo después del cual, el T50 detuvo el desarrollo de entrenudos (22,3), mientras que los demás tratamientos desarrollaron un mayor número (29,44, 27,33 y 26,37 para T150, T0 y T100, respectivamente) (Fig. 1D). Esta variable mostró un ajuste adecuado al modelo Gompertz en ambos ciclos (R 2>0,83 primer ciclo y >0,90 segundo ciclo). Para el primer ciclo y segundo ciclo no se observaron diferencias estadísticas en la tasa de crecimiento de entrenudos.
Emisión de botones florales y formación de frutos
La emisión de botones florales mostró un comportamiento muy variable en el tiempo sin diferencias significativas entre tratamientos con la prueba de Tukey (P≤0,05). La mayor tasa de emisión de botones florales se observó en T50 a los 7 y 21 ddt (3,58 y 3,56 botones por rama y semana, respectivamente) (Fig. 2); mientras que la menor emisión se observó en T0 (0,95 botones por rama y semana). En general, la mayor emisión de botones florales se observó en los primeros 28 ddt, lo cual coincide con la aplicación de los tratamientos.
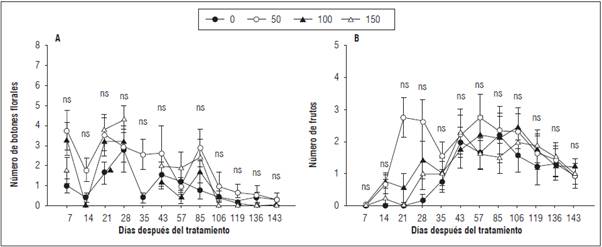
Figura 2 Emisión de órganos reproductivos y productivos en plantas de gulupa (Passiflora edulis f. edulis Sims) fertilizadas con diferentes dosis de boro foliar aplicado en la formación del botón floral y cuajado de fruto (T0: 0 kg B ha-1; T50: 0,3 kg B ha-1; T100: 0,6 kg B ha-1 y T150: 0,9 kg B ha-1). A, número de botones florales; B, número de frutos. Las barras verticales indican el error estándar n=9. * indica diferencias significativas entre tratamientos con la prueba de Tukey (P≤0,05). ns: sin diferencia estadísticamente.
Cuajado de fruto
El cuajado de frutos presentó diferencias significativas con la prueba Tukey (P<0,05) bajo los diferentes tratamientos foliares con B. En el primer ciclo, el mayor porcentaje se alcanzó en T100 (85,19%), seguido por T50 (81,48%) y luego T0 y T150 (62,96%) (Fig. 3A). Por su parte, en el segundo ciclo productivo, el tratamiento T50 presentó el mayor porcentaje de cuajado de fruto (90,0%), seguido por T100 (73,3%), T0 (63,3%) y T150 (50%) (Fig. 3B).
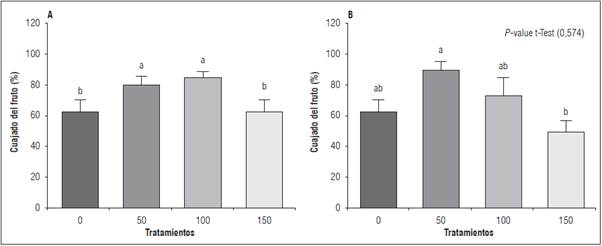
Figura 3 Cuajado de frutos en plantas de gulupa (Passiflora edulis f. edulis Sims) fertilizadas con diferentes dosis de boro foliar aplicado en la formación del botón floral y cuajado de fruto en dos ciclos productivos (T0: 0 kg B ha-1; T50: 0,3 kg B ha-1; T100: 0,6 kg B ha-1 y T150: 0,9 kg B ha-1). A, primer ciclo; B, segundo ciclo. Las barras verticales corresponden al error estándar n= 10. Promedios con letras distintas indican diferencia significativa entre tratamientos de acuerdo con la prueba de Tukey (P≤0,05). P-valor de la prueba de t de student ≤0,05 indica diferencias entre los dos ciclos de producción.
Desarrollo de frutos
El diámetro ecuatorial de frutos de gulupa no presentó diferencias significativas entre tratamientos en los primeros 51 ddt (P>0,05) para el primer ciclo del cultivo; posteriormente, los frutos de T100 mostraron valores más altos, alcanzando el mayor diámetro ecuatorial a los 87 ddt (55,63 mm) (Fig. 4A). A diferencia del primer ciclo, en el segundo se presentaron diferencias (P<0,05) desde el inicio del experimento. Los frutos de T50 desarrollaron un mayor diámetro ecuatorial desde los 21 ddt, alcanzando el máximo diámetro (55,68 mm) a los 136 ddt. Para los demás tratamientos, T0, T100 y T150 el diámetro máximo de fruto se alcanzó a los 143 ddt, con valores de 51,82, 53,75 y 52,92 mm, respectivamente (Fig. 4B). El modelo Gompertz para esta variable se ajustó adecuadamente en los dos ciclos del cultivo, siendo mayor en el segundo (R 2>0,90), comparado con el primero (R 2>0,56). Las mayores tasas de crecimiento del fruto se alcanzaron en T100 y T50 (0,63 y 0,38 mm d-1, respectivamente), con valores similares en el segundo ciclo.
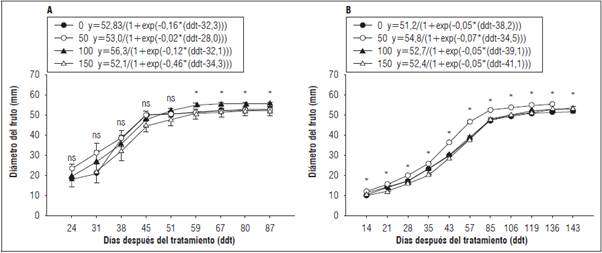
Figura 4 Diámetro de frutos en plantas de gulupa (Passiflora edulis f. edulis Sims) fertilizadas con diferentes dosis de boro foliar aplicado en la formación del botón floral y cuajado de fruto en dos ciclos productivos (T0: 0 kg B ha-1; T50: 0,3 kg B ha-1; T100: 0,6 kg B ha-1 y T150: 0,9 kg B ha-1). A, primer ciclo; B, segundo ciclo. Las barras verticales indican el error estándar n = 10. Se muestra la ecuación del modelo Gompertz para cada uno de los tratamientos. * indica diferencias significativas entre tratamientos de acuerdo con la prueba de Tukey (P≤0,05). ns: sin diferencia estadísticamente.
DISCUSIÓN
Se encontró que la aplicación foliar de B en plantas de gulupa en etapa de floración presentó un efecto positivo en el crecimiento de ramas secundarias con el tratamiento T50 para los dos ciclos evaluados; sin embargo, no se observó un patrón similar en el número de entrenudos, lo que sugiere que la emisión de nudos es una característica intrínseca de la planta, la cual no se ve afectada por los tratamientos con B. Resultados similares, pero en ramas primarias, han sido reportados en P. edulis f. flavicarpa, donde la aplicación de B genera incremento de la longitud de ramas en un 20,27%, así como no se presentó efecto en el número de nudos (Añez y España, 2011), pero se registró un incremento del 62% en la producción de materia seca de la parte aérea de las plantas (Prado et al., 2006). Probablemente las plantas bajo deficiencia de B presentan una reducción en el crecimiento de la pared celular (Kocábek et al., 2009; Tang et al., 2011; Marschner, 2012; Cong et al., 2015). Además, en condiciones de deficiencia o exceso de B se afectan las células de los vasos conductores debido a que las traqueidas se lignifican (Demiray y Dereboylu, 2013) y como consecuencia se observa una reducción en el crecimiento de las plantas, siendo así importante un adecuado suministro de B, debido a que más del 90% del B presente en las plantas está en las paredes celulares (Mei et al., 2016).
El incremento en el porcentaje de cuajado de fruto de gulupa, observado en este estudio, indica que la aplicación exógena de B favorece a la fructificación y que bajo deficiencia de B se reduce el número de frutos, lo cual concuerda con datos reportados para P. edulis f. flavicarpa (Freitas et al., 2006; Añez y España, 2011). Resultados similares también han sido reportados para otras especies frutales como Malus domestica (Wojcik et al., 2008), Pyrus communis (Shalan, 2013), Prunus persica (Yadav et al., 2013), Persea americana (Boldingh et al., 2016), entre otras.
En la presente investigación se observó que el mayor porcentaje de cuajado de frutos se obtuvo con la aplicación de B en etapa de formación de botón floral con los tratamientos T100 y T150 para los dos ciclos productivos. Resultados similares han sido reportados para Pyrus communis (Shalan, 2013), Solanum melongena (El-Gawad y Osman, 2014) y Ribes grossularia (Wójcik y Filipczak, 2015). Los resultados obtenidos para el desarrollo de botón floral y cuajado de fruto fueron similares a otros trabajos; se observó que la aplicación de B sobre los tejidos reproductivos mejora la producción de flores y cuajado del fruto, debido a que estos órganos son altamente demandantes de B por las altas tasa de crecimiento y las paredes celulares ricas en pectina (Marschner, 2012; El-Gawad y Osman, 2014). Por lo tanto, las aplicaciones foliares de B son especialmente valiosas en el período de desarrollo de las flores, y más aún en especies donde la movilidad del B se restringe al floema, por consiguiente, el desarrollo de las flores depende de un suministro continuo de B (Perica et al., 2001).
En cuanto al diámetro de los frutos de gulupa, los tratamientos T100 y T50 presentaron los mejores valores en los dos ciclos evaluados. Resultados similares han sido observados para otras especies trepadoras como Actinidia deliciosa, donde la aplicación de B aumentó en 8,27% el diámetro de los frutos (Sotomayor et al., 2010), y en otras especies frutales tropicales como Artocarpus heterophillus donde se registró incremento del 29,83% en la longitud y de 38,88% en el diámetro del fruto (Halder et al., 2008). La aplicación foliar de B mejora significativamente las características físicas de los frutos (diámetro, longitud y peso) lo cual puede atribuirse a que el B aumenta la tasa de transporte de azúcares a los vertederos y zonas de crecimiento y por tanto mejora el crecimiento de los frutos (Mengel et al., 2001). Adicionalmente, se ha considerado que el B es absorbido rápidamente por las flores (Sarrwy et al., 2012) y al ser aplicado de manera foliar, permite que sea disponible más rápidamente en etapas demandantes. En este sentido, las aplicaciones de B en estas etapas generan un efecto rápido sobre el desarrollo de botones florales y frutos.
CONCLUSIONES
Las plantas de gulupa, sometidas al tratamiento T0 mostraron para ambos ciclos productivos no tener la mejor respuesta de crecimiento y desarrollo con respecto a los otros tratamientos evaluados. Se observó que para los dos ciclos productivos el tratamiento T50 presentó la mayor longitud de ramas secundarias, el mejor porcentaje de cuajado de fruto y un adecuado diámetro de fruto para comercialización. Por otro lado, el tratamiento T100 mostró los mejores resultados para desarrollo de botón floral, emisión de botones florales y número de frutos. En general, se considera que las mayores limitaciones se presentaron bajo el tratamiento T0 y por el contrario la aplicación del tratamiento T150 no mostraron efecto negativo marcado sobre las variables evaluadas, sugiriendo que este tratamiento no representó una condición de exceso de este elemento.


