Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkINTRODUCCIÓN
Fusarium oxysporum Schltdl. 1824, es una de las especies morfológicamente más diversas y ampliamente distribuida del género Fusarium (Baayen et al., 2000). Esta especie, se constituye en un complejo, con numerosas formas especiales las cuales son morfológicamente idénticas, solo pueden ser diferenciadas por el tipo de planta(s) atacada(s) (Leslie y Summerell, 2006), con alrededor de 100 formas especiales. Fusarium se caracteriza por producir colonias de rápido crecimiento y morfología muy variable, predominando dos tipos primordiales, una micelial y una pionotal (Garcés et al., 2001). En general, la mayoría de cepas patógenas pueden atacar las plantas de tomate a lo largo de todo su ciclo de desarrollo, produciendo síntomas de marchitamiento, acompañada del amarilla-miento parcial de las hojas y el doblamiento de los brotes de la planta enferma. Una característica que permite diagnosticar y diferenciar rápidamente esta enfermedad es la coloración blanquecina, amarillenta o marrón en los haces vasculares; adicionalmente, se puede presentar enanismo en los brotes y disminución del crecimiento general de la planta (Garcés et al., 1999).
Sin embargo, los síntomas exteriores en la planta hospedera no siempre se detectan rápidamente, o esta puede permanecer asintomática durante su ciclo de desarrollo; esto implica que una planta infectada sea en ocasiones difícil de detectar y pueda ser una hospedera asintomática que actúe como reservorio del patógeno, convirtiéndose en una fuente de contaminación para el resto del cultivo (Kolander et al., 2012). En la actualidad se utiliza una gran cantidad de técnicas de detección de enfermedades de plantas, pero éstas presentan varias limitaciones que disminuyen su eficiencia al usarlas para obtener resultados que conduzcan a decisiones de control. Por ejemplo, la detección basada en la observación de síntomas en enfermedades vasculares implica que cuando hay disturbios visibles el hongo u organismo patógeno haya invadido ya el tejido vascular. Los métodos de detección que se basan en técnicas moleculares, microscopía óptica, microscopía electrónica de transmisión, bioensayo para la detección de virus, y la serología, tienen buenos niveles de precisión, pero requieren la destrucción de las muestras, consumen tiempo, son costosos y requieren alta capacitación profesional, por lo que su uso es modesto, implicando imprecisión porque la enfermedad se diagnostica usando tejidos o plantas completas que se destruyen, no siendo óptimas en el sentido estricto de detección temprana y en tiempo de avance de enfermedades de plantas (Sankaran et al., 2011).
Es por estas razones, y a pesar de la disponibilidad de estas técnicas, un sistema de detección temprana de la enfermedad basado en técnicas de espectroscopía puede ayudar a disminuir las pérdidas causadas en los cultivos y evitar una mayor propagación de las enfermedades, evitando por ejemplo la siembra de unidades de plantas portadoras del patógeno, con más rapidez, sensibilidad, selectividad y sin demandar la destrucción de muestras requeridas para el análisis (Chaerle y Van der Straeten, 2000). En la última década se han realizado muchos estudios en los cuales fue aplicada la espectroscopía y particularmente, el uso de radiación VIS en la detección de enfermedades en plantas sintomáticas (Sankaran et al., 2011; Abu-Khalaf y Salman, 2014; Szuvandzsiev et al., 2014); sin embargo, aquellos enfocados en la detección de enfermedades en plantas asintomáticas o estudios enfocados a enfermedades que pueden "enmascarar" los síntomas usan la espectrometría de reflectancia en proporciones relativamente bajas. Por lo anterior, el objetivo de este estudio fue evaluar un método de detección y discriminación de la respuesta espectral debida a cambios fisiológicos en plantas de tomate asintomáticas infectadas con dos cepas de F. oxysporum usando es-pectroscopía de reflectancia en la región visible (VIS).
MATERIALES Y METODOS
Material vegetal
En este estudio se usó la variedad de tomate, tipo Chonto Santa Cruz®, resistente a F. oxysporum. Las semillas fueron plantadas en bandejas de germinación de 76 pozos con turba estéril como sustrato y se mantuvieron en una cámara de siembra con temperatura de 26±1°C, humedad relativa de 60±10% y fotoperiodo de 12 h con luz blanca fluorescente, con una intensidad entre 40 a 80 mol-2 s-1 durante el tiempo de los experimentos. Las plantas se regaron diariamente y se fertilizaron una vez por semana con una solución hidropónica que contenía (mili-equivalentes): NO3 (12,0), PO4 (1,0), K (1,7), Mg (1,5), Ca (2,8) y S (0,5), y en micro-equivalentes, Fe (70,0), Mn (18,0), Zn (7,7), Cu (1,5), B (27,5) y Mo (0,5). A las 4 semanas después de germinadas (sdg) se realizó el procedimiento de inoculación y las plantas se transfirieron a recipientes de 90 cm3 que contenían el mismo sustrato que el usado en plantulación. Las plantas inoculadas se mantuvieron en las mismas condiciones de cámara de siembra durante el resto del experimento, aumentando las fertilizaciones a dos veces por semana con las mismas concentraciones en macro y micronutrientes (Lafontaine y Benhamou, 2010).
Preparación de aislamientos de F. oxysporum
Se usaron dos cepas de F. oxysporum aisladas de hospederos diferentes; la cepa F05 fue aislada de Passiflora edulis (gulupa) (cepa A54; Ortiz y Hoyos-Carvajal, 2016) y la cepa F07 fue aislada de plantas de Solanum lycopersicum (tomate). Ambas fueron mantenidas en cajas de Petri con agar papa dextrosa (Merck®) PDA + 50 mg kg-1 de sulfato de estreptomicina (SE). Multiplicándose según las necesidades.
Inoculación de plantas de tomate antes del trasplante definitivo
Para este experimento se usaron 12 plantas de 4 sdg siguiendo la metodología de inoculación descrita por Ortiz y Hoyos-Carvajal (2016) y 12 plantas control. Se utilizó la concentración 1106 esporas/mL por cada aislamiento de Fusarium de 10 d de cultivo. Cuidadosamente las raíces de las plántulas fueron lavadas con agua de grifo hasta retirar los restos de turba. Luego de la limpieza se hacen heridas (cortes) en las raíces terciarias de todas las plantas que se infectaron con tijeras esterilizadas y se sumergen solo las raíces en 100 mL de la solución de esporas durante 10 min. Las plántulas inoculadas y controles fueron trasplantados a los vasos con turba estéril de 5 cm3. En las plantas control se realizó el mismo procedimiento de las plantas inoculadas, pero solo con agua destilada. Para llevar a cabo los postulados de Koch se realizaron muestreos destructivos a los 11 y 22 días pos-inoculación (dpi).
Prueba de infectividad
Para confirmar la infección de las plantas se realizaron dos muestreos destructivos, en los estados 1.10 (11 dpi) y el estado 2.20 (22 dpi), según la escala BBCH para las mono y dicotiledóneas (Feller et al., 1995). Se realizó la prueba de infectividad mediante la técnica de indexación en medio PDA + SE a 50 mg kg-1 (Ortiz y Hoyos-Carvajal, 2016). A los 5 d después de sembrar los segmentos indexados de tallo en el medio usado se observó la presencia y crecimiento del patógeno.
Espectroscopia
Para la adquisición de espectros de reflectancia VIS se utilizó un espectroscopio portátil USB2000+ (Ocean Optics, Largo, FL) con una fuente de luz halógena de tungsteno HL-2000-HP (rango de longitud de onda de 360-2.400 nm), un estándar de reflectancia difusa modelo WS-1 (reflectividad >98% en el rango de 2501.500 nm) y una sonda de reflectancia grado premium de 600 QR600-7-VID-125F (Ocean Optics, Largo, FL, USA). Las mediciones se realizaron en los estados fenológicos 1.10 y 2.20 de la planta en las hojas 2 y 3 con la fibra óptica en la cara adaxial obteniéndose cinco espectros por cada hoja. El tiempo de integración utilizado en esta investigación fue de 1 ms, un promedio de 10 lecturas por medición y "tiempo de intervalo" de 2000 μs.
Análisis de datos
La discriminación y comparación de los efectos causados por ambas cepas se llevó a cabo en hojas en el mismo estadio de crecimiento y desarrollo, sólo usándose hojas desarrolladas y de la misma edad (hoja dos de cada planta). Los análisis realizados en los datos de las plantas 22 dpi tienen un mayor muestreo. Intencionalmente, se usó mayor cantidad de plantas, tratando de prever una mortalidad natural que al fin del experimento fue muy baja, por lo que no todas las muestras contienen el mismo número de espectros.
Inicialmente, se realizó una selección de los espectros en la cual se retiraron algunos con ruido (deformados y/o con error de lectura). Estos espectros que mostraron patrones muy diferentes se confirmaron con análisis de los "valores atípicos" identificados en un Análisis de Componentes Principales (PCA) sin tratamiento previo de datos. Después de la eliminación de los espectros con ruido y tratamientos previos, se aplicó la combinación de las transformaciones Corrección Dispersión Multiplicativa (MSC) más la "segunda derivada" como mejor pre-tratamiento, lo que permite una buena agrupación de las plantas en los tratamientos realizados (Isaksson y Naes, 1998). Luego del pre-tratamiento de datos se realizaron los PCA y LDA enfocados a evaluar la posible separación entre plantas infectadas y las de control en diferentes tiempos pos-infección (Fig. 1).
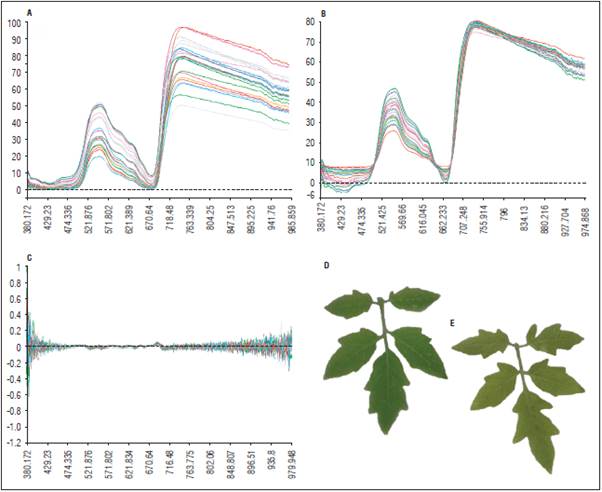
Figura 1 Ejemplo de datos espectrales usados para los análisis mostrando la diferencia entre los pre-tratamientos aplicados y hojas monitoreadas del día 22 después de inoculación: A. Perfiles espectrales sin transformación; B. Corrección de dispersión Multiplicativa (MSC); C. MSC + Segunda derivada; D. Hoja de planta control; E. Hoja inoculada.
Los espectros de las plantas infectadas con cada cepa de F. oxysporum (F05 y F07) se analizaron separados con los controles (no se realizó tratamiento combinando las cepas) y en los dos tiempos pos-infección. El conjunto total de datos tomados de las hojas 2 y 4 de las plantas infectadas y controles se compone de 156 espectros (promedios de cinco espectros tomados por hoja). En este análisis solo se tomaron los 78 espectros correspondientes a la hoja 2 para evitar ruido relacionado con la diferencia en el tiempo de desarrollo de ambas hojas, los cuales fueron medidos desde 10 plantas control, 10 infectadas con la cepa F05 y 10 infectadas con la cepa F07 en el día 11, y 13 controles, 20 infectadas con la cepa F05 y 17 infectadas con la cepa F07 en el día 22. Después de la eliminación de los espectros con ruido y "atípicos" se usaron 64 espectros en total.
RESULTADOS
Confirmación de la infección
Se evaluó la incidencia de la enfermedad en plantas de tomate inoculadas con F. oxysporum en plantas de tomate con base en el porcentaje de individuos infectados en relación con el total de las plantas inoculadas. La incidencia de la cepa F05, que fue aislada de una planta de gulupa, fue mayor a la observada en la cepa F07 (aislada de una planta de tomate), presentando valores del 60,0% a los 11 d y 81,8% a los 22 d. La cepa F07 presentó incidencias de 30,0% y 64,3% en los tiempos evaluados (11 y 22 d, respectivamente).
Análisis de datos espectrales
El Análisis de Componentes Principales (PCA) obtenidos de los espectros de las hojas de tomate infectadas con las cepas de F. oxysporum, F05 y F07, se agruparon según la cepa y tiempo pos-infección. La cepa F05 presentó mejor agrupamiento en los dos periodos de tiempo pos-inoculación con respecto a los controles (Fig. 2A y 2A') y la cepa F07 (Fig. 2C y 2C'). Sin embargo, la cepa F07 no presentó un agrupamiento claro a los 11 dpi y mejoró considerablemente para a los 22 dpi (Fig. 2B y 2B'). Es importante resaltar que estos resultados son coherentes con los obtenidos en los muestreos destructivos para confirmación de la infección, en la cual se dio mayor porcentaje de infección para la cepa F05 en ambos periodos de tiempo, y la cepa F07 tuvo un porcentaje muy bajo de infección en el día 11 (30%).
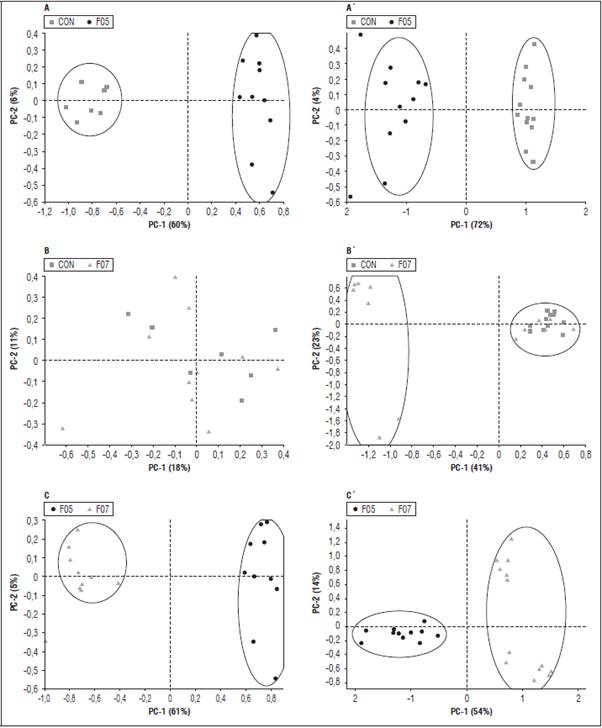
Figura 2 Análisis de Componentes Principales de espectros de tomate de mesa infectadas con dos cepas de F. oxysporum y dos tiempos después de la infección (dpi). A. Cepa F05 a los 11 dpi; A'. Cepa F05 a los 22 dpi; B. Cepa F07 a los 11 dpi; B'. Cepa F07 a los 22; C. Cepas F05 y F07 a los 11 dpi; C'. Cepas F05 y F07 a los 22 dpi.
El PCA realizado con la cepa F05 junto con sus controles (Fig. 2A y 2A') se emplearon 41 espectros, los componentes principales CP1 y CP2 explican el 66% (CP1 = 60%, CP2 = 6%) y 76% (CP1 = 70%, CP2 = 6%) de la variabilidad total de los datos en los 11 y 22 dpi, respectivamente. Adicionalmente, se usaron 43 espectros para los PCA realizados con la cepa F07 más los respectivos controles (Fig. 2B y 2B'), en los cuales los CP1 y CP2 solo explican el 29% (CP1 = 18%, CP2 = 11%) y 64% (CP1=41%, CP2 = 23%) en los 11 y 22 dpi, respectivamente. Finalmente, se usaron 22 espectros de hojas infectadas con la cepa FO5 y 22 de la cepa FO7 para evaluar si el método pude separar plantas infectadas con aislados diferentes de la misma especie (Fig. 2C y 2C'); en estos PCA, los CP1 y CP2 explicaron el 66% (CP1=61%, CP2=5%) y 68% (CP1=54%, CP2=14%) de la variabilidad total de los datos en los días 11 y 22 dpi, respectivamente.
El LDA permitió mejorar la separación de los respectivos tratamientos. Los LDA obtenidos a partir de los datos espectrales de la cepa F05 (Fig. 3A y 3A') alcanzaron el 100% de precisión obtenidos con la aplicación de cinco componentes principales que predicen las muestras de acuerdo con el tratamiento al cual pertenecen. Por el contrario, los LDA obtenidos de los espectros de las hojas infectadas con la cepa F07 obtuvieron una precisión de solo 58,82% a los 11 d (Fig. 3B) y un aumento al 80,77% en el día 22 (Fig. 3B') con la misma aplicación de cinco componentes principales para la predicción de muestras.
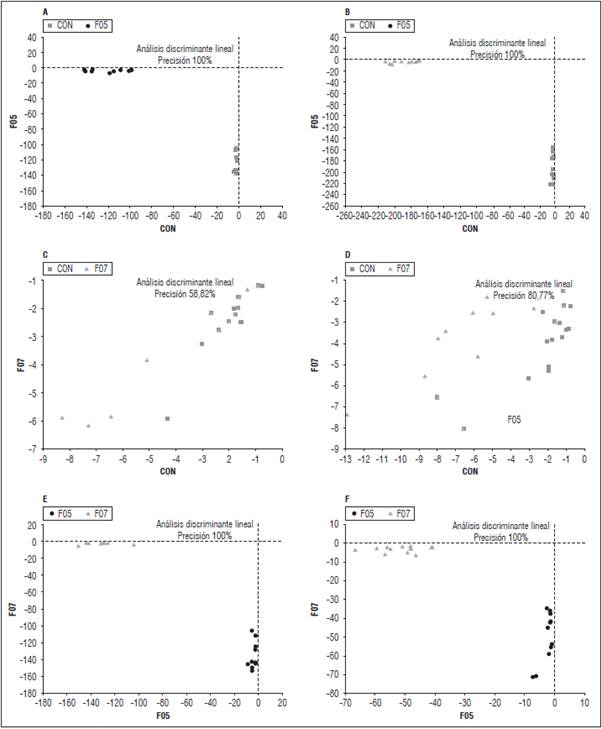
Figura 3 Análisis discriminantes lineales de espectros de tomate de mesa infectadas con dos cepas de F. oxysporum y dos tiempos después de la infección (dpi). A. Cepa F05 a los 11 dpi; A'. Cepa F05 a los 22 dpi; B. Cepa F07 a los 11 dpi; B'. Cepa F07 a los 22; C. Cepas F05 y F07 a los 11 dpi; C'. Cepas F05 y F07 a los 22 dpi.
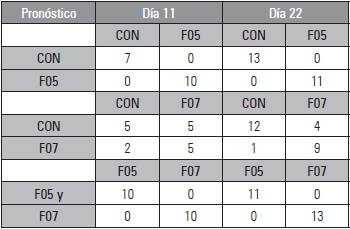
En las matrices de confusión se puede confirmar que los diferentes modelos clasificaron correctamente todas las plantas infectadas con la cepa F05 con respecto a las inoculadas con la cepa F07 y controles en ambos días de medición (precisión el 100%), mientras que para la cepa F07 el 50% se clasificó como control el día 11 y el 44 % el día 22 (Tab. 1), lo que denota una clara dificultad del modelo en la discriminación entre las plantas no inoculadas y plantas inoculadas con la cepa F07, las posibles razones serán abordadas en la discusión.
DISCUSIÓN
No se observaron los síntomas típicos del marchitamiento por F. oxysporum en el tiempo del ensayo, como epinastia, marchitez no uniforme de hojas o ramas y decoloración vascular extendida por encima de la línea del suelo. Aunque las raíces no fueron observadas en el trascurso del experimento, el muestreo destructivo permitió confirmar la presencia de ambas cepas (F05 y F07) en los periodos de tiempo evaluados (11 y 22 d después de inoculación) mediante identificación morfológica clásica. Se encontró mayor incidencia de la cepa F05 en las plantas de tomate en comparación con la cepa F07, ya que fueron las plantas con mayor incidencia en ambos muestreos destructivos, lo cual fue confirmado en los análisis PCA y LDA realizados con los datos espectrales. Se puede considerar que la cepa F05 tiene una alta capacidad para infectar las plantas de tomate, sin embargo, debido a la resistencia de las variedades de tomate (Jones y Crill, 1974; Rivard y Louws, 2011; Morid et al., 2012), se imposibilita la aparición de los síntomas.
Las hojas enfermas producen cambios fisiológicos significativos antes de la colonización de las hifas de F. oxysporum, por lo que es posible la clasificación de una planta como "enferma" con criterios basados en fluoresencia y/o espectroscopia una o dos semanas antes de que aparezcan los síntomas evidentes. Es importante destacar la concordancia de nuestros resultados con los obtenidos en el estudio de Lorenzini et al. (1997), en el cual encontraron que los parámetros de fluorescencia de la clorofila de las primeras hojas asintomáticas fueron alterados por la infección de F. oxysporum hasta el día 14 después de la infección, debido posiblemente a una disminución de la fluorescencia variable y confirmado por la actividad alterada del fotosistema II. También es importante destacar las diferencias obtenidas entre las dos cepas de F. oxysporum usadas para infectar las plantas de tomate, debido a que la cascada de eventos que ocurren en las células inoculadas con microconidias de aislados con diferente nivel de patogenicidad cambia la relación específica planta-patógeno; desafortunadamente, los mecanismos implicados son complejos y aún no están claramente definidos (Olivain et al., 2003).
Los resultados encontrados en los espectros de reflectancia concuerdan con los muestreos destructivos. Además, se observó una dinámica característica de los espectros de reflectancia medidos en el rango espectral para los días 11 y 22 dpi en las cepas estudiadas, F05 y F07. Esta dinámica se puede observar claramente cuando se comparan las mayores incidencias obtenidas de los muestreos destructivos en las plantas infectadas con los análisis de los datos espectrales (PCA y LDA) de la cepa F05, en los cuales las plantas se agrupan claramente y puede discriminarse de las plantas sin enfermedad y las infectadas en los dos días de medición. Por el contrario, la enfermedad tiene una baja incidencia en las plantas infectadas con la cepa F07 (particularmente en el día 11) y en los análisis de los espectros de reflectancia solo es posible agrupar y discriminar los individuos infectados al día 22, cuando la incidencia sube al 64,3%.
Las alteraciones fisiológicas o bioquímicas entre las plantas infectadas y sanas pueden ser causadas por la composición química del parásito o una respuesta desde la planta hospedera, implicando cambios en la ubicación de nutrientes y energía, hasta producción de metabolitos secundarios (Ramachandra y Ravishankarb, 2002), además de posibles respuestas causadas por estrés en las plantas sugerido por cambios en segmentos del infrarrojo cercano (Ferri et al, 2004; Gitelson et al., 2005; Ustina et al., 2009). En ese sentido, los resultados de este estudio son contrarios a los obtenidos por (Spinelli et al., 2006), los cuales usaron técnicas basadas en espectroscopia de reflectancia en el NIR para detectar el "fuego bacteriano" en plantas de peral asintomáticas, sin lograr discriminar las plantas infectadas de plantas sanas. Los autores discuten que sus resultados posiblemente fueron debido a la pequeña escala de medición en la hoja (2 mm2), lo cual pudo ser contrarrestado en nuestro estudio ya que se tomaron cinco mediciones por hoja en las cuales cada espectro medido es el resultado promedio de 100 mediciones.
Adicionalmente, es importante destacar que las respuestas fisiológicas e histológicas en plantas hospederas asintomáticas y en etapas tempranas de la infección fúngica que están relacionadas con cambios en las propiedades ópticas de las hojas en las plantas aún son desconocidas en su mayor parte. Además, la mayoría de estudios enfocados a predecir la química foliar son basados en análisis de tejido seco, lo que imposibilita el muestreo de la misma hoja en el tiempo (Fourty et al., 1996; Couture et al., 2013), lo cual ofrece grandes ventajas para la caracterización de respuestas de las plantas a patógenos usando muestras frescas y/o muestreo in situ como lo abordaron en Franke y Menz, (2007), Naidua et al. (2008), Mahlein et al. (2012), en los cuales se usaron técnicas de teledetección en cultivos de trigo para detectar infección con oídio y roya de la hoja en diferentes periodos de tiempo del cultivo. Estos resultados concuerdan con la base teórica propuesta por Zhang et al. (2003), en la cual se sugiere que la magnitud de onda variará típicamente en diferentes longitudes, aumentando la reflectancia en el VIS y disminuyendo en el NIR (7501100 nm) en las plantas infectadas con patógenos y sometidos a diferentes tipos de estrés.
CONCLUSIONES
Se probó un método para detectar y discriminar la respuesta espectral debida a cambios en la fisiología de las plantas de tomate asintomáticas infectadas con dos cepas de F. oxysporum usando espectroscopía de reflectancia. Los resultados sugieren que las plantas de tomate pueden mantener niveles relativamente altos de infección sin mostrar síntomas visibles y que es posible detectar y discriminar infecciones asintomáticas de F. oxysporum en plantas usando espectroscopia de reflectancia con radiación en el VIS/NIR. Las cepas de F. oxysporum usadas en este estudio causaron diferencias claras en los espectros de reflectancia en las plantas infectadas y las no infectadas, además de permitir una buena discriminación entre las plantas infectadas con ambas cepas.
Los resultados obtenidos indican que la espectroscopia VIS/NIR es promisoria para la detección de hongos vasculares, incluso en plantas asintomáticas. Adicionalmente, la aplicación de técnicas multivariadas como PCA y LDA a datos espectrales en el VIS/ NIR de muestras frescas inoculadas con diferentes cepas de F. oxysporum ha mostrado ser un método eficiente para la detección y discriminación de infecciones fúngicas.
Aunque son varias las técnicas espectroscópicas y de imágenes que se han usado para la detección de enfermedades en plantas sintomáticas, es muy baja la cantidad de estudios especializados en la detección de enfermedades en plantas asintomáticas usando técnicas no destructivas como la espectroscopía de reflectancia. En este sentido, los resultados de este estudio son valiosos como contribución en esta área del conocimiento. Adicionalmente, la correspondencia entre la medición de la incidencia de la enfermedad y los cambios encontrados por la técnica de espectroscopía de reflectancia en plantas asintomáticas sugiere que es un método sensible y confiable que puede ser adecuado para el diagnóstico temprano de enfermedades en plantas. Sin embargo, los mecanismos fisiológicos y anatómicos que influyen en la expresión de síntomas de la planta debido a las infecciones fúngicas aun parecen muy complejos y no están claramente definidos, particularmente aquellos mecanismos presentes en etapas tempranas de la infección, lo cual invita a desarrollar nuevos proyectos de investigación que profundicen y aclaren esta área del conocimiento de gran importancia en enfermedades tanto vasculares como sistémicas.