











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Quinoa (Chenopodium quinoa Willd.), a novel crop to the world, has been continuously selected in the Andes of South America, characterizing its gradual expansion from around Titicaca Lake, between Bolivia and Peru, the probable center of origin (Maughan et al., 2004). The expansion of quinoa growth occurred to the North (Ecuador, Colombia, and Venezuela) and to the South (Argentina and Chile), from the Andean Altiplano to the valleys and coastal regions. The expansion of quinoa cultivation occurred slowly, dispersing into environments of great climatic differences (Bertero et al., 2004).
Peru and Bolivia have been the major quinoa producers, followed by Ecuador and Argentina (Perez-Rea and Antezana-Gomez, 2018). In Brazil, the interest in quinoa cropping started in the 1990’s, as an option for diversification of cropping systems its excellent food source. In addition, with the no-till system evolution, it contributes biomass to protect the soil in the dry season and uses low quantities of seeds in sowing, a favorable factor to expand cultivation (Spehar et al., 2015a).
Thermal units, also known as growing degree days and heat units, are a way of incorporating both temperature and time into one measurement to quantify the rate of plant growth in response to temperature. Calculation of thermal units allows defining of the accumulated thermal units (ATU) for each phase of plant growth and development (Renato et al., 2013). ATU has been successfully employed in agriculture, particularly in phenological studies. The concept of thermal time, first introduced in 1730 by R. A. F. de Reaumur, to predict phenological events more affected by location and sowing date, as an alternative for the number of days, affected by the temperature of location (McMaster and Wilhelm, 1997).
Employing or calculation of thermal units can be useful in predicting the phases of plant growth and development, such as seedling emergence, early growth, flower initiation, reproductive period, and its subdivisions. Thermal units can be a tool to determine the sowing and harvest time of crops affected by climate change. The rise in temperature speeds up the phases; therefore, using the number of days alone has been no longer valid to assess growth and development phases (Sharma et al., 2021). Variations in air temperature can anticipate the phenological phases, turning the events of crop growth less unpredictable when measured in the number of days (Souza et al., 2017).
In the calculation of thermal units, one must consider the base temperature (BT), which is specific for every plant crop species. In quinoa it has been reported that BT seems to vary according to phenological phase (Garcia-Parra et al., 2020a). Quinoa has shown a high potential to adapt to different environments, being exposed to variable temperatures, affecting the length of plant phenology (Jojoa et al., 2021).
Clearly defined phenological stages are of great importance for phenotype reproducibility. Several studies have investigated and described the phenological stages of quinoa. These studies have provided valuable information on crop characterization (Stanschewski et al., 2021). Currently, the phenological stages of quinoa are described using the Biologische Bundesanstalt Bundessortenamt und Chemische Industrie (BBCH) scale, presenting as main phases germination (0), leaf development (1), formation of lateral buds (secondary stems) (2), inflorescence emergence (5), flowering (6), fruit development (7), ripening (8), senescence (9) (Sosa-Zuniga et al., 2017). Secondary phases are adapted to the changing behavior of different cultivars, which are influenced by environmental effects and their generic character (Garcia-Parra et al., 2020b).
Quinoa genotypes have shown variability in phenological phases as response to temperature (Anchico et al., 2020). Studies using (ATU) to describe quinoa growth and development are scarce and do not provide comprehensive information to predict the phenological phases of the crop. The information is essential to guide research on crop breeding and management for efficient production. In view of this, the study had the objective to calculate the thermal units for 13 codes of the BBCH scale of quinoa from the evaluation of 12 selected progenies grown in two sowing dates in the Brazilian Savannah.
MATERIALS AND METHODS
Physiographic characteristics of location and soil management
Two experiments (Season 1 - March / June 2018 and Season 2 -May / August 2019) were conducted in Água Limpa Farm, University of Brasilia (UnB), Federal District, Brazil, coordinates of 15° 56’ S and 47° 55’ W, at an altitude of 1,100 m. It is located in the core of the Cerrado Region (Brazilian Savannah physiognomy). The climate has been described according to Köeppen, as Aw, characterized by rainy period, October to April, and dry period, May to September (Kottek et al., 2006). Mean temperature during the experiments was 19.0 and 19.45ºC, with maximum mean temperature of 27.6 and 28,8ºC, and minimum mean temperature of 12.5 and 11.5ºC. Rainfall was 451.7 mm and 141.5 mm, respectively for the two sowing dates (Fig. 1).
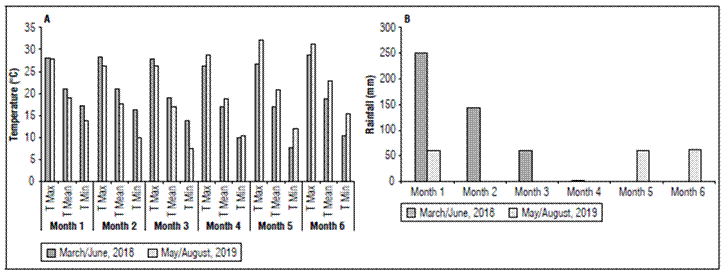
Figure 1. Temperatures and rainfall two growing period in 2018 and 2019 (March/June, 2018) and (May/August, 2019).
Prior to setting experiments, the area was fertilized according to the requirements of quinoa cropping with N, P2O5, and K2O sources in the proportion of 12:90:48 kg ha-1, harrowed, leveled off, and rows equally spaced by 0.5 m were opened (Spehar et al., 2015b). Thirty days after emergence, the plots received N in the dose of 40 kg ha-1, band applied, and were kept weed-free by hand hoeing. The experiments were sprinkler irrigated when water tension reached 40 kPa based on tensiometers installed in the area. For the quinoa cycle, 350 mm of irrigation water was required.
Quinoa genotypes and selected progenies
Progenies were obtained in Brazil and in Colombia at altitudes of 1,100 e 1,800 m by individual plant selection in existing cultivars (Anchico et al., 2020): a) Progenies BRQ1, BRQ 2, BRQ 3, BRQ 4, BRQ 5, BRQ 6, BRQ 8, and BRQ 10, from cultivar BRS Syetetuba (Brazil), of plant maturity cycle between 95 to 121 d and yield in the range of 1,500 and 2,200 kg ha-1; b) progeny from cultivar Aurora (AUR) (Colombia) of 97 d to maturity and 2,121 kg ha-1 yield; c) progeny from cultivar Blanca Dulce de Jerico (BLA) (Colombia) 150 d to maturity and 1,500 kg ha-1 yield; d) progeny of the cultivar Tunkahuan (TUN) originally from Ecuador and selected in Colombia, 118 d to maturity and 1,300 kg ha-1 yield e) cultivar Piartal (PRI) originally from Ecuador and selected in Colombia, 109 d to maturity and 1,500 kg ha-1 yield.
Phenological evaluation
The first evaluations were made between March and June 2018 and the second between May and August 2019. The phenological stages of quinoa in this research were classified according to the BBCH scale, which are described below: BBCH-08 (Hypocotyl with cotyledons growing towards soil surface, BBCH-10 (Cotyledons fully emerged), BBCH-11 (First pair of leaves visible) , BBCH-12 (Second pair of leaves visible), BBCH-13 (Third pair of leaves visible), BBCH-20 (Visible lateral buds or expanded leaves without lateral stems), BBCH-50 (Inflorescence present but still enclosed by leaves), BBCH-51 (Leaves surrounding inflorescence separated, inflorescence is visible from above), BBCH-59 (Inflorescence visible, but all the flowers are still closed), BBCH-69 (Complete anthesis: main inflorescence flowers with senesced anthers), BBCH-81 (Milky grain, easily crushed with fingernails, liquid content and green pericarp), BBCH-85 (Thick grain, easily crushed with fingernails, white pasty content, green, beige, red or black pericarp), BBCH-89 (Ripe grain, difficult to crush with fingernails, dry content, the grain has a beige, red or black colour on its outside. Ready to harvest) (Sosa‐Zuniga et al., 2017).
Determination of thermal units
Thermal units were determined for the thirteen phenological phases of quinoa growth and development of 12 progenies, on the basis of maximum and minimum daily temperatures obtained in the meteorological station of Água Limpa Farm, UnB.
Calculation of accumulated thermal units (ATU) for quinoa considered the base temperature of 3.1ºC according to (Bertero et al., 1999). This temperature is the minimum at which the quinoa plant paralyzes growth.
Accumulated thermal units (ATU) was estimated according to the method proposed by Arnold (1959). Two equations were used (1)
where, TU was thermal units, MT the maximum daily temperature, mt the minimum daily temperature and BT the base temperature.
The thermal summation or (ATU), was calculated by equation 2.
where, n was number of days to reach each plant growth and development phase.
Experimental design and statistical analysis
The experiments were conducted based on the randomized complete block design. Each block was consisted of eight progenies of selected individual plants from cultivar BRS Syetetuba, recommended for cultivation in the Brazilian Savannah (Spehar et al., 2011), in the period 2016-2018 and four progenies, two from Colombian cultivars Blanca Dulce de Jerico (BLA) and Aurora; and two from Ecuadorian cultivars Tunkahuan and Piartal. The four progenies were selected in 2017-2018. Altogether, each block contained 12 plots corresponding to progenies on six repetitions. The plot had four rows 2 m long, equally spaced by 0.5 m, with a distance between plots of 1 m. The plant density on the row was 30 plants/m-1.
An analysis of variance of the values of (ATU) for phenological phases between genotypes and sowing dates was carried out and compared by Tukey's test using SPSS software. Additionally, a hierarchical analysis was performed to evaluate the relationship between progenies and (ATU) values for phenological phases and a dendogram was obtained, using the Euclidean distance, for similarity grouping (Ward, 1963).
RESULTS AND DISCUSSION
Quinoa progenies had statistically significant F test differences at P≤0.01and P≤0.05 of (ATU) for BBCH-11 to BBCH-89. (Tab. 1). Comparisons were made by ATU, standardizing the values independently of the growing environment (Zapata et al., 2015). Therefore, it is expected in this experiment that the genotype grown on the two sowing dates present similar ATU values. This holds to other crops of different base temperatures as soybeans (Spehar et al., 2015a).
Table 1. Accumulated thermal units (ATU) of quinoa progenies for the two planting dates (March / June, 2018) and (May / August, 2019) at BBCH-08 (hypocotyl with cotyledons growing towards soil surface), BBCH-10 (cotyledons fully emerged), BBCH-11 (first pair of leaves visible), BBCH-12 (second pair of leaves visible), BBCH-13 (third pair of leaves visible), BBCH-20 (visible lateral buds), BBCH-50 (inflorescence present but still enclosed by leaves), BBCH-51 (leaves surrounding inflorescence separated,), BBCH-59 (inflorescence visible), BBCH-69 (complete anthesis), BBCH-81 (milky grain), BBCH-85 (thick grain), and BBCH-89 (ripe grain).
| P/E.F. | BBCH-08 | BBCH-10 | BBCH-11 | BBCH-12 | BBCH-13 | BBCH-20 | BBCH-50 | BBCH-51 | BBCH-59 | BBCH-69 | BBCH-81 | BBCH-85 | BBCH-89 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SEASON 1 - MARCH / JUNE 2018 | |||||||||||||
| AUR | 56.98 a | 112.55 a | 191.95 c | 305.75 c | 434.24 c | 533.13 d | 933.34 f | 1057.30 h | 968.14 j | 1093.28 i | 1382.28 h | 1608.81 k | 1817.32 h |
| PRI | 56.98 a | 112.55 a | 191.95 c | 394.83 a | 454.37 b | 572.26 b | 1035.58 bcd | 1185.72 d | 1110.9 f | 1220.81 f | 1600.02 d | 1819.83 e | 1981.49 e |
| BRQ1 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 536.38 cd | 910.24 g | 1032.42 i | 987.29 i | 1051.11 j | 1307.55 i | 1605.88 k | 1804.81 i |
| BRQ4 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 539.70 cd | 968.14 e | 1075.96 g | 1016.56 h | 1110.90 h | 1376.13 h | 1600.02 k | 1788.43 j |
| BRQ5 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 533.13 d | 1019.59 d | 1204.04 c | 1125.99 de | 1237.52 d | 1435.65 g | 1638.50 j | 1885.21 g |
| BRQ10 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 533.13 d | 1047.84 abc | 1204.04 c | 1141.61 bc | 1250.76 c | 1466.01 f | 1714.83 h | 2008.15 d |
| TUN | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 533.14 d | 1038.74 bcd | 1185.72 kd | 1135.31 bcd | 1224.17 ef | 1468.78 f | 1684.84 i | 1918.39 f |
| BRQ6 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 533.13 d | 1032.42 cd | 1244.20 a | 1160.58 a | 1292.09 a | 1600.02 d | 1770.44 f | 1918.39 f |
| BRQ2 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 539.63 cd | 1048.02 abc | 1224.17 b | 1160.58 a | 1270.19 b | 1617.6 c | 1734.97 g | 2008.15 d |
| BRQ8 | 56.98 a | 112.55 a | 232.53 b | 354.07 b | 434.24 c | 533.13 d | 1022.94 d | 1129.01 f | 1087.50 g | 1179.45 g | 1644.29 b | 1908.39 b | 2031.27 c |
| BRQ3 | 56.98 a | 112.55 a | 232.53 b | 394.83 c | 454.37 b | 552.67 c | 1063.52 a | 1224.17 b | 1147.91 ab | 1270.19 b | 1667.84 a | 1983.83 a | 2099.23 b |
| BLA | 56.98 a | 112.55 a | 232.53 b | 373.48 ab | 434.24 c | 533.33 d | 1044.92 abc | 1204. 04 c | 1132.16 cde | 1234.18 de | 1670.67 a | 1898.96 c | 2096.65 b |
| SEASON 2 -MAY / AUGUST 2019 | |||||||||||||
| AUR* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 613.97 l | 689.54 n | 661.34 o | 761.65 q | 1066.83 k | 1346.81 p | 1565.25 n |
| BRQ1* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 613.97 l | 704.16 m | 676.48 n | 777.24 p | 1066.83 k | 1346.81 p | 1565.25 n |
| BRQ4* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 704.16 k | 777.24 l | 747.11 m | 841.35 o | 1066.83 k | 1346.81 p | 1565.25 n |
| BRQ5* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 761.65 j | 841.35 k | 816.43 l | 912.11 m | 1234.21 j | 1391.76 n | 1612.88 k |
| BRQ10* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 761.65 j | 841.35 k | 816.43 l | 912.11 m | 1234.21 j | 1565.25 l | 1759.04 k |
| BRQ6* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 777.24 ij | 841.35 k | 816.43 l | 912.11 m | 1234.21 j | 1377.30 o | 1565.25 n |
| BRQ8* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 777.24 ij | 841.35 k | 816.43 l | 912.11 m | 1234.21 j | 1565.25 l | 1759.04 k |
| BRQ2* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 777.24 ij | 841.35 k | 829.61 k | 897.63 n | 1234.21 j | 1461.42 m | 1630.12 l |
| BRQ3* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 777.24 ij | 841.35 k | 829.61 k | 912.11 m | 1234.21 j | 1565.25 l | 1759.04 k |
| PRI* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 792.23 hi | 868.14 j | 841.35 k | 943.14 l | 1234.21 j | 1565.25 l | 1759.04 k |
| TUN* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 364.17 d | 451.14 e | 803.89 h | 868.14 j | 841.35 k | 958.26 k | 1234.21 j | 1565.25 l | 1759.04 k |
| BLA* | 50.66 a | 100.11 a | 202.17 c | 305.75 c | 481.93 a | 704.16 a | 1052.98 ab | 1169.31 e | 1121.17 ef | 1234.21 de | 1565.25 e | 1924.92 a | 2381.00 a |
| S.D. | 3.18 | 6.24 | 16.00 | 32.35 | 39.90 | 61.16 | 148.50 | 186.53 | 167.14 | 173.63 | 195.88 | 189.10 | 203.7 |
| C.V. % | 5.89 | 5.87 | 7.45 | 9.65 | 9. 83 | 12.09 | 16.67 | 18.58 | 17.45 | 16.40 | 14. 30 | 11.64 | 11.10 |
| MEAN | 53.83 | 106.33 | 213.97 | 335.20 | 405.79 | 505.81 | 890.78 | 1003.93 | 957.85 | 1058.78 | 1369. 85 | 1624.64 | 1834.9 |
In each column means followed by same letters in column do not differ significantly based on the Tukey test (P≤0.05).
The progenies studied here had variable ATU, turning into a reference to compare adaptability to cropping systems in environments different from quinoa’s Andean origin (Anchico et al., 2020). All progenies had seedling emergence (BBCH-08) in 3 d in 2018 and 2019, with 56,98 and 50,66 ATU (Tab. 1), expressing sensitivity to the temperature at this phase, as already described experimentally in Argentina (González et al., 2017). This early phase of quinoa has shown more sensitivity to temperature. For the remaining phases, there were different values of ATU as related to days, turning evident the effect of temperature in plant development (Salazar-Gutiérrez et al., 2013). In Cotyledons fully emerged (BBCH-10), there were no statistical differences among progenies, with ATU of 112.55 in 2018 and 100.11 in 2019. In the BBCH-11 (first pair of leaves visible), there were statistical differences in the 2018 experiment; progenies AUR and PRI had less accumulated thermal units, 191.95, than the other progenies of 232.53. In 2019 all progenies had 202.17 ATU at BBCH-11. When progenies for the two experiments were compared, three groups of ATU were formed (Tab. 1).
At BBCH-12 progenies, PRI and BLA had different accumulated thermal units (ATU) at first sowing date of 394.83 and 373.48, respectively. In the second experiment, all progenies had an ATU of 305.75 for the same phase (Tab. 1). At the BBCH-13, mean values were ATU 405, with a smaller mean value in the second year with a mean ATU of 364,17. Progeny with higher ATU was BLA with 481.93. At BBCH-20, the mean ATU was 505.81, while progenies in the second year had smaller ATU, although BLA had 481.93. These differences, although statistically significant, are still acceptable to compare accumulated thermal units (ATU) for progenies at each growth and development phase in quinoa. Other uncontrolled environmental factors such as rainfall, relative air moisture, evapotranspiration, solar radiation could have influence in the plant vegetative and early reproductive phases (Parra-Coronado et al., 2015).
The statistical differences were more predominant from BBCH-50, separating the progenies into early, mid-cycle and late maturity. In the first experiment, progenies with smaller ATU were AUR and BRQ 1, with 933,34 and 910.24 respectively, while the largest were BRQ 3 and BRQ 2 with 1063.52 and 1048.02 ATU (Tab 1). Differences are accentuated from the beginning of the reproductive phase. A similar finding was reported for Cucurbita moschata (Souza et al., 2017). At BBCH-51, progenies had an ATU larger mean value of 1,164.23; for the same phase mean value was 843.72 in the second experiment, AUR 613.97 was the least thermal unit accumulator progeny (Tab. 1). BBCH-50 and BBCH-51showed a similar trend, with AUR (661.34), BRQ1 (676.48), and BRQ4 (747.11), early progenies having the least ATU, while BRQ2 and BRQ6 were late (1,160.58). At BBCH-69, significant differences were found among progenies of early, mid-cycle, and late maturity grouping. Early AUR (761.65) and BRQ 1 (777.24) contrasted with late BRQ 6 (1292.09) and BRQ 2 (1270.19G) (Tab. 1).
In the second experiment, the progenies reduced the number of days until the BBCH-59, which was reflected in the accumulated thermal units. Temperature fluctuations could have influenced these variations at flowering reflecting in the following phases (Reguera et al., 2018). The mean ATU between BBCH-69 and BBCH-81 was 304.5 with the least values for AUR*, BRQ1*, and BRQ4*, accumulating 1,066.83 heat units (Tab. 1). Progenies BRQ3 and BLA accumulated 1,667.84 and 1,670.67 heat units. AUR had the least ATU, confirming results obtained in Colombia (Anchico et al., 2020; Montes-Rojas et al., 2018). ATU at BBCH-85 showed the same trend for AUR, BRQ1, and BRQ4 accumulating 1.346.81 thermal units, while BRQ3 (1.983.83) and BLA (1.924.92) were the latest. At BBCH-89, there were statistical differences for progenies and sowing dates. The phenological plant cycle, in days, for late maturity progenies extended to 145 d, while early maturity AUR*, BRQ1* and BRQ4* with respective ATU of 2,096.65, 2,099.23, and 2,381.00. The mean ATU difference was 223.11, indicating a reduction in phenological phases (Fig. 2).
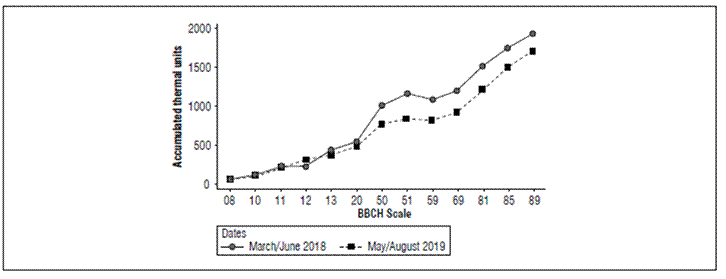
Figure 2. Accumulated thermal units (ATU) for the two planting dates at BBCH-08 (hypocotyl with cotyledons growing towards soil surface), BBCH-10 (cotyledons fully emerged), BBCH-11 (first pair of leaves visible), BBCH-12 (second pair of leaves visible), BBCH-13 (third pair of leaves visible), BBCH-20 (visible lateral buds), BBCH-50 (inflorescence present but still enclosed by leaves), BBCH-51 (leaves surrounding inflorescence separated,), BBCH-59 (inflorescence visible), BBCH-69 (complete anthesis), BBCH-81 (milky grain), BBCH-85 (thick grain), and BBCH-89 (ripe grain).
Progenies selected from BRS Syetetuba had statistical differences for BBCH-89, demonstrating ample variability, attained by possible segregation for the lateness in the cultivar originated from natural crosses (Spehar et al., 2015b). The spread of progenies into early, mid-cycle and late maturity has kept the relationship among them when tested in Colombia at different altitudes and temperatures, althoughs kept the relationship when tested in Colombia at different altitudes and temperatures. However, the ATU at BBCH-89 were not statistically different (Anchico et al., 2020).
In this experiment, progenies were classified up to BBCH-89, expressing potential adaptability relating to the environment they were selected (Bois et al., 2006). Progeny BLA of cultivar Blanca Dulce de Jerico had the highest ATU (2.239) (Tab. 2), extending the plant maturity cycle to180-214 d at environments above 2,000 m and low temperature (Montes-Rojas et al., 2018). This would be predicted by the use of accumulated heath units in quinoa cultivation instead of the number of days to maturity.
Table 2. Accumulated thermal units (ATU) of 12 quinoa progenies in BBCH-08 (hypocotyl with cotyledons growing towards soil surface), BBCH-10 (cotyledons fully emerged), BBCH-11 (first pair of leaves visible), BBCH-12 (second pair of leaves visible), BBCH-13 (third pair of leaves visible), BBCH-20 (visible lateral buds), BBCH-50 (inflorescence present but still enclosed by leaves), BBCH-51 (leaves surrounding inflorescence separated,), BBCH-59 (inflorescence visible), BBCH-69 (complete anthesis), BBCH-81 (milky grain), BBCH-85 (thick grain), and BBCH-89 (ripe grain).
| Progenies | BBCH-08 | BBCH-10 | BBCH-11 | BBCH-12 | BBCH-13 | BBCH-20 | BBCH-50 | BBCH-51 | BBCH-59 | BBCH-69 | BBCH-81 | BBCH-85 | BBCH-89 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BRQ1 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 493.8b | 762.1b | 868.3b | 831.9b | 914.2b | 1187.2d | 1476.3e | 1685d |
| BRQ2 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 495.4b | 912.6ab | 1032.8ab | 995.1ab | 1083.9ab | 1425.9abc | 1598.2bcde | 1819.1bcd |
| BRQ3 | 53.822a | 106.33a | 217.35a | 350.3a | 409.3ab | 501.9b | 920.4ab | 1032.8ab | 988.8ab | 1091.2ab | 1451ab | 1774.5ab | 1929.1b |
| BRQ4 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 495.4b | 836.1b | 926.6b | 881.8b | 976.1b | 1221.5cd | 1473.4e | 1676.8d |
| BRQ5 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 492.1b | 890.6ab | 1022.7ab | 971.2ab | 1074.8ab | 1334.9bcd | 1515.1de | 1749bcd |
| BRQ6 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 492.1b | 904.8ab | 1042.8ab | 988.5ab | 1102.1ab | 1417.1abc | 1573.9cde | 1741.8bcd |
| BRQ8 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 492.1b | 900.1ab | 985.2ab | 952ab | 1045.8ab | 1439.3abc | 1736.8abc | 1895.2b |
| BRQ10 | 53.822a | 106.33a | 217.35a | 329.91a | 399.2b | 492.1b | 904.7ab | 1022.7ab | 979ab | 1081.4ab | 1350.1bcd | 1640bcde | 1883.6bc |
| AUR | 53.822a | 106.33a | 197.06b | 329.91a | 399.2b | 492.1b | 773.7b | 873.4b | 814.7b | 927.5b | 1224.6cd | 1477.8e | 1691.3cd |
| PRI | 53.822a | 106.33a | 197.06b | 350.3a | 409.3ab | 511.7b | 913.9ab | 1026.9ab | 976.1ab | 1082ab | 1417.1abc | 1692.5bcd | 1870.3bcd |
| TUN | 53.822a | 106.33a | 214.82ab | 342.9a | 393.4b | 485.3b | 901.7ab | 1000.5ab | 964.4ab | 1069.1ab | 1332.2bcd | 1615.1bcde | 1825.4bcd |
| BLA | 53.296a | 106.33a | 217.35a | 339.6a | 458.09a | 618.7a | 1048.95a | 1186.68a | 1126.66a | 1234.2a | 1618a | 1911.94a | 2239a |
| S.D. | 0,15 | 0,29 | 7,82 | 8,31 | 17,22 | 36,32 | 74,33 | 85,05 | 82,489 | 85,71 | 120,67 | 136,23 | 154,95 |
| CV (%) | 0.28 | 0.28 | 3.66 | 2.48 | 4.25 | 7.19 | 8.36 | 8.49 | 8.63 | 8.11 | 8.82 | 8.39 | 8.45 |
| MEAN | 53.78 | 106.24 | 213.76 | 335.20 | 405.31 | 505.23 | 889.14 | 1001.78 | 955.85 | 1056.86 | 1368.24 | 1623.795 | 1833.8 |
In each column means followed by same letters in column do not differ significantly based on the Tukey test (P≤0.05).
The use of ATU helped to describe the duration of phenological phases of quinoa, which can be useful to manage the crop in all phases best, guiding the time for fertilization, plant protection, irrigation and in genotype selection to originate different maturity groupings (Anandhi, 2016). Moreover, predictability ATU of phenological phases can direct selection in quinoa to face climate changes (Sharma et al., 2021).
When the two sowing dates are considered, progenies accumulated less heat units in May/August 2019, although the number of days was higher. This could be explained by uncontrolled factors, as moisture availability. The experiment conducted in March/June 2018 received more water because, in addition to irrigation, there was considerable rainfall in the period, whereas in May/August experiment relied almost entirely on irrigation. Excess water in the first experiment and lower temperatures in the second could help explaining differences in ATU and in days to each phase (Tab. 1). Differences for ATU among the phases, although were statistically different, showed a similar pattern in the groupings (Tab. 2). Higher temperatures in the second experiment could explain the anticipation of growth and development phases in quinoa (Asseng et al., 2011; Parra-Coronado et al., 2015).
In general, progenies accumulating lower thermal units were BRQ4 (1.676,8), BRQ1 (1,685), and AUR (1,691). The ones with higher ATU were BLA (2.239), BRQ3 (1,929.1 GDD), and BRQ8 (1,895) (Tab. 2). Late progenies were identified from BBCH-13 onwards, presenting higher ATU values, a characteristic that was maintained until BBCH-89 (Tab. 2).
Between the beginning of the reproductive phase BBCH-50 and the mature grain phase BBCH-89, an ATU of 944.66 (Tab. 2) was presented, representing 51% of the total of the studied cycle.
Temperatures, higher than in Andean Valleys and in Colombia, conditioned reduction in days to plant cycle, accelerating enzymatic activities in the plant and the phenological phases (Asseng et al., 2011; Souza et al., 2017).
Euclidean distance of 5 was used for the 12 progenies based on accumulated thermal units in the 13 codes of the BBCH scale, according to (Hair et al., 2005). Five hierarchical groups of similarity were defined (Fig. 3).
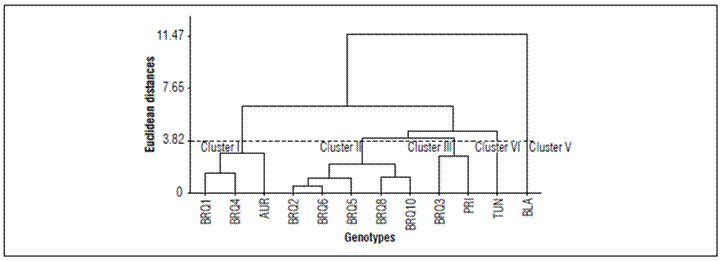
Figure 3. Dendrogram for hierarchical grouping of 12 quinoa progenies by using accumulated thermal units (ATU).
Group I was formed by three progenies, two selected from BRS Syetetuba (BRQ 1 and BRQ 4) and one Aurora (AUR). These progenies presented lower thermal units, therefore they were the most precocious.
Grupo II contained five progenies, all selected from BRS Syetetuba (BRQ2, BRQ6, BRQ5, BRQ8, BRQ10). These differed from the preceding groups, with higher ATU.
Group III had two progenies, one from BRS Syetetuba (BRQ3) one from Piartal (PRI), dissimilar to the group I.
Group IV is made up of one progeny of Tunkahuan (TUN), dissimilar to preceding groups I, with higher ATU.
Group V had one progeny of Blanca Dulce de Jerico (BLA), being the highest thermal unit accumulator and dissimilar to other groups.
Hierarchical clustering allowed determining the similarity of genotypes in the accumulation of thermal units, showing a significant difference between genotypes in the first and fourth groups. This helped to identify early, medium and late genotypes. The grouping helped to visualize the relationship among progenies and relate the plant maturity cycle to the ATU. There were other factors differentiating the experiments as excess water in the first sowing date that could have caused a departure from expected results. Even though they were statistically different, there was a similar trend, turning the predictable definition of maturity groups based on ATU.
CONCLUSION
The use of accumulated thermal units allows the prediction of phenological events in quinoa of different maturity groups. The BRQ4, BRQ1 and AUR progenies were the earliest with the lowest Accumulated Thermal Units (ATU) values, averaging 1676, 1685 and 1691, respectively. However, the progenies BLA and BRQ 3 were the later with ATU values of 2239 and 1929, respectively. The accumulated thermal units for BBCH-89 ranged from 1565.25 to 2381, with a difference between the earliest and latest genotypes of 815.75. Progenies selected from existing cultivars are different in thermal unit accumulation, ensuing efficiency in cultivar acquisition to stagger quinoa cultivation. Accumulated thermal units explain the range of plant maturity cycles in selection. The calculation of ATU for the BBCH scale codes is an efficient tool to predict the phenological cycle of quinoa.

