











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Potato (Solanum tuberosum L.) is the world's fourth most important food crop after rice, wheat, and maize (Thongam et al., 2017; Seminario-Cunya et al., 2018). The total world potato production was estimated at 376,119,974 t in 2021. In Colombia, potatoes are important as a staple food for the population, reaching 2,621,344 t in 120,190 ha in 2021 (FAO, 2023), of which 254,446 t in 15,514 ha corresponded to diploids potatoes (Solanum tuberosum Phureja group) (Agronet, 2023). Furthermore, this country presents a high diversity of cultivars of S. tuberosum Phureja group (Estrada, 1996). The Phureja group potatoes are diploid cultivars characterized by a short crop cycle (3 to 4 months), limited tuber dormancy, and their sensitivity to photoperiod for tuberization, showing an adaptation to short days and fast maturation time (Lizarazo et al., 2022). This group is cultivated at altitudes between 2,200 and 3,200 m a.s.l. Therefore, compared to the tetraploid potato, the Phureja group is less cultivated and productive. However, this difference in yield is compensated by a high organoleptic quality, good acceptance for consumption, and, generally, a higher sale price (Rojas and Seminario, 2014), with highly heterogeneous shape and size of tubers, with a short postharvest time and rapid sprouting (Rojas and Seminario, 2014; Zárate-Polanco et al., 2014).
Potato is a very weather-sensitive crop. Its growth and production are influenced by weather, temperature, the use of improved varieties, nutrition, incidences of pests, and diseases (Estrada, 1996; Sánchez et al., 2005; Saldaña et al., 2015; Torres, 2020). Furthermore, in the tropics, planting time plays a critical role in potato production as manipulation of light, water availability, and temperature can be done to a certain extent by altering planting time (Singh et al., 2013; Thongam et al., 2017; Obiero et al., 2019). The environment is the primary variable affecting crop production, mainly the yield of potatoes. Hence, knowledge of how it influences crop development, growth, and yield is of great interest to researchers. Successful potato crop production requires the efficient use of the climatic resources of solar radiation, temperature, and water, among many others (Geremew et al., 2007).
Functional plant growth analysis refers to quantitative methods that describe the performance of whole plant systems grown under natural, semi-natural, or controlled conditions. One of the main applications is to analyze the differences in the yield potential of genotypes and also to know plant development over time (Khan et al., 2014). Plant growth analysis (PGA) is a collective technique used to quantify growth components, such as crop growth rate, relative growth rate, net assimilation rate, specific leaf area, etc. PGA regulates the crop yield that is a function of total dry matter production and its effective distribution to economically important plant organs (Mbah and Eke-Okoro, 2015). Plant growth analysis provides an explanatory, holistic, and integrative approach to interpreting plant form and function (Pommerening and Muszta, 2016). It has been widely used to study the factors that influence plant growth (Saldaña and Cotes, 2020).
In this context, the concept of plant growth analysis relative to plant size has proved to be a powerful tool in comparative studies of the growth performance of plants with applications in dry weight, biomass, leaf area, stem volume, basal area, and stem diameter (Pommerening and Muszta, 2016). In potato, knowledge about the rate of tuber formation and development, it is important to know the dynamics of leaf and stem formation and maintenance, responsible for photo assimilate production processes (Silva et al., 2020). Since the translocation of assimilates is fundamental for the final performance, understanding the biomass distribution in the different potato organs and the influence of the environment and the genotype will allow a better selection of materials for some region or purpose (Saldaña et al., 2015). The knowledge of the accumulation and distribution of biomass in potato genotypes will allow the measurement of growth rate and tuber yield (Ñústez et al., 2009). Also, this information is important to determine the best stages for performing cultural treatments, such as the planting density to be used, heaping, fertilizer timing and dose application, phytosanitary management, and tuber harvest time (Silva et al., 2020).
From an economic point of view, dry matter accumulation is an essential indicator in potato cultivation since the tuber is a storage organ for photo assimilation (Saldaña et al., 2015). A better understanding of the growth and development of the plant and the temporal relationships between the different events will allow knowing the physiological bases that lead to the production of tubers (Celis-Gamboaet al., 2003). Describing the accumulation and distribution of the dry matter and the foliar area throughout the crop cycle will allow more precise agronomic management, such as the opportune moment to control pests and diseases. Finally, there is a need for studies on the growth and distribution pattern of photoassimilates in modern varieties obtained through plant breeding to adapt to the agronomic management of the crop (Santos et al., 2010). Due to those mentioned above, and because diploid potato is highly adaptable to different environmental conditions, the functional growth analysis is relevant to understanding and comparing varieties in physiological and productive behavior. In this sense, the present study aims to analyze the growth of four diploid potatoes Phureja group varieties in the Colombian high Andean region.
MATERIALS AND METHODS
Location
This work was developed between November 2019 and July 2020, at the Estación Agraria Paysandú of the Universidad Nacional de Colombia, Medellín - Colombia, located at 6°12'36" N and 75°30'25" W, at an elevation of 2,538 m a.s.l., classified as a lower montane forest zone (bh-MB) (Holdridge, 1967).
Experiment design
A randomized complete block design was used with four treatments and three replications. The treatments consist of four registered diploid potato varieties: Paola, Primavera, Paysandú, and Violeta, obtained by Universidad Nacional de Colombia in 2015 (Ñústez, 2018). Each experimental unit consisted of 24 plants planted 0.9 m between rows and 0.25 m between plants. Two growing seasons were carried out, the first between September 2019 and January 2020 and the second between March and July 2020. The seed used for the two growing seasons was taken from the basic seed cluster that is part of the seed maintenance program of the evaluated varieties. Properties of the soil used in the experiment were Loam texture, pH (4.9), OM (22.2 %), phosphorus (44 mg kg-1 soil), sulfur (17 mg kg-1 soil), Ca (3.7 cmolc kg-1), Mg (1.2 cmolc kg-1 ), K (0.77 cmolc kg-1), ECEC (7.2 cmolc kg-1), Fe (98 mg kg-1), Mn (1 mg kg-1), Cu (1 mg kg-1), Zn (4 mg kg-1) and B (0.7 mg kg-1). Based on the soil condition, creole potato plant nutrition was applied using chemical fertilization: N (2.55 g), P2O5 (2.55 g), and K2O (2.55 g) per plant at sowing and the exact doses 30 days after sowing.
Weather conditions
During the evaluation period, a Watch Dog Series 2000 portable climatic station was used to daily measure temperature (°C), relative humidity (%), and precipitation (mm). The average temperature and relative humidity in the 2019 season were 16.8°C and 80.43%; in 2020, it was 17.6 °C and 76.78%, respectively. The total precipitation during the cultivation cycle was 615.6 and 594 mm for the 2019 and 2020 seasons, respectively (Fig. 1).
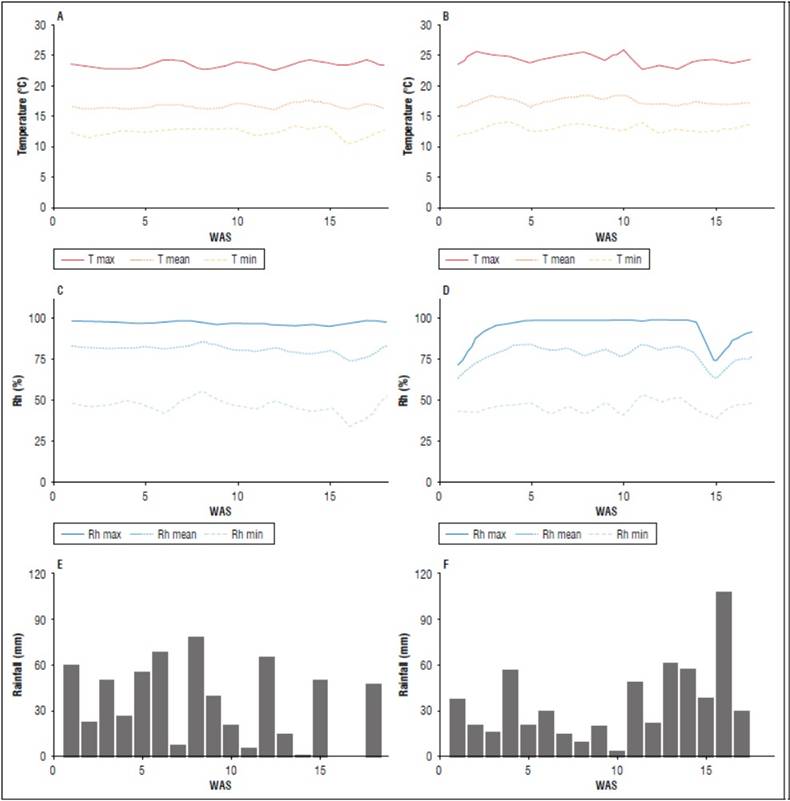
Figure 1. Climatic variables during the growing season. a) and c) are the weekly maximum (Tmax), mean (Tmean), and minimum (Tmin) temperatures in 2019 and 2020, respectively. b) and d) are the weekly maximum (Rhmax), mean (Rhmean), and minimum (Rhmin) relative humidity (Rh) in 2019 and 2020, respectively. e) and f) are weekly accumulated rainfall in 2019 and 2020, respectively. WAS (weeks after sowing).
Variables evaluated
From the sixth week after sowing the number of main stems, stolons, tubers, and dry weight per organ (leaves, stems, roots, stolons, tubers, flowers, and fruits) per experimental plant was determined during 12 weeks in 2019 and 11 weeks in 2020. Each organ was placed in drying ovens (Memmert type UL 80) at 70 °C until reaching a constant weight. The final weight was considered the dry matter in each structure, and their sum was the plant's total dry weight. Before drying, the whole leaf area produced by each plant was estimated using a portable Li-Cor (Li-3000) leaf area analyzer. For the final tuber harvest, the observational criterion proposed by Ñústez and Rodríguez (2020) was used, which is based on vigorously rubbing the skin of the tuber, considering it at physiological maturity or harvest point, when the skin is firm, that is, when the tuber does not peel. In the Phureja group, harvest maturity for tubers occurs without foliage senescence.
Plant Phenology
It used the BBCH scale for Solanum tuberosum L. to determine the beginning and duration of flowering, fruiting, and tuber (Enz and Dachler, 1998). The beginning of the flowering and fruiting stages (61 and 71) were considered when 50% of the evaluated plants had at least one open flower and one fruit appeared, respectively. The end of flowering was established when less than 50% of the plants had no flowers (69), and the fruit's development was when the maximum weight of the fruits per plant was reached (79). The beginning of a tuber was defined when this organ had a diameter greater than 0.5 cm, and when it got its maximum dry weight, it was considered phenological stage 48.
Growth analysis
From the data of dry matter by organ obtained, a traditional functional growth analysis was made. The Net Assimilation Rate (NAR), Relative Growth Rate (RGR), Leaf Area Ratio (LAR), and the Power of the Source and Demand were determined (Kooman and Rabbinge, 1996; Hunt, 2003). The number and weight of tuber (g), yield per plant (kg), and harvest index were determined at harvest.
Thermal time
The plant growth was measured through growing degree days (AGDD - °Cd-1), using equation (1) to calculate degree days (Riaño et al., 2005).
Tmax and Tmin correspond to the maximum and minimum temperature on the day, respectively, and Tbase is the base temperature. A base temperature of 2°C and an upper limit of 29°C were used (Kooman and Rabbinge, 1996; Saldaña et al., 2015; Soto et al., 2018).
Statistical analysis
Functional analysis of the distribution of dry weight, leaf area, growth rates, percentage of plants with flowers and fruits, power of sink-source, and the number of stems, stolons, and tubers throughout was made. For the phenological stage duration and tubers yield, variances among different treatments were analyzed by one-way ANOVA and multiple comparisons by the Kruskal-Wallis test at 5% probability because assumptions for validating the analysis of variance were not fulfilled. The R project "Agricolae" package (R Core Team, 2020) was used.
RESULTS AND DISCUSSION
There was a difference in the accumulation of growing degree days (AGDD) necessary for the harvest between both crop seasons, 1816 AGDD (112 days after sowing - DAS) 2019 season and 1895 AGDD (125 DAS) for the 2020 season. Historically, potato growers have used calendar days and experience to predict key plant developmental stages. However, there may be better indicators than the calendar day to measure the growth situation of crops (Liao et al., 2022). Developing and utilizing the growing degree day system may be a more accurate predictor of these stages throughout the season to determine optimal planting dates and yields compared to calendar days (Worthington and Hutchinson, 2005). According to the accumulated growing degree days (AGDD) concept, the crops can only continue to grow when a certain accumulative heat is reached for a specific phenological phase (Liao et al., 2022).
According to Thongam et al. (2017), the sowing date affects the days this species requires to complete the different development phases and significantly influences further growth and performance parameters. Cooler and higher temperatures reduce net assimilation to the tubers, while higher temperatures may prevent tuber initiation. According to Soto et al. (2018), it has been shown in several studies that the temperature influences the beginning of tuberization and also affects dry matter production through the daily growth rate and indirectly through its relationship with development (Kooman and Haverkort, 1995; Santos, 2010).
The leaf area of a variety is related to the leaf area index and the conformation of the canopy for light absorption (Cabezas and Corchuelo, 2005). In both production seasons, the varieties with the highest leaf area were Violeta and Paola. In 2019, Paola produced 7,313 cm2 at 1285 AGDD (78 DAS); while Violeta reached 9,337 cm2 at 930 AGDD (57 DAS). In 2020, the largest leaf area observed was 16,854 and 16,406 cm2 for the Paola and Violeta varieties, achieved in both varieties at 1089 AGDD (63 DAS). The Paysandú variety stood out as the variety that produced the smallest leaf area in both seasons (Fig. 2). The greater leaf area was reached in 2020, possibly associated with a higher temperature. In this sense, Santos (2010) stated that there is a lower development of the foliar area at a higher altitude and lower temperature as an adaptation to more significant direct radiation.
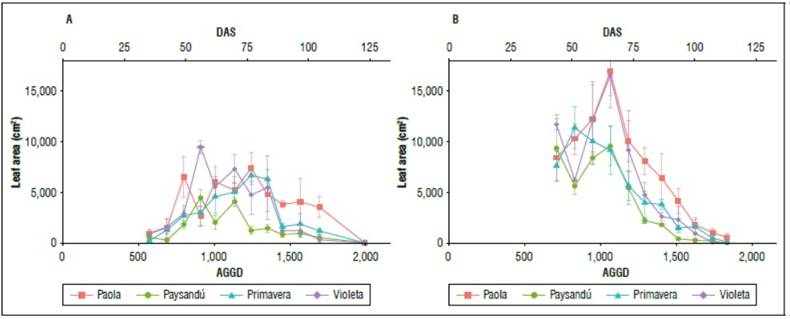
Figure 2. Leaf area during the crop growing. a) 2019 season and b) 2020 season. Accumulated growth degree days (AGDD) and days after sowing (DAS). Data are mean ± standard error.
Potato cultivars show considerable diversity in growth rates due to their genetic makeup and environmental interaction. Therefore, studying dry matter partitioning and distribution in the various plant organs during crop development is essential to determine a cultivar's growth rate and tuber yield. Concerning the accumulated dry matter in the different organs evaluated, the Paola variety had the greater biomass in both cycles. Instead, Paysandú had the lower total dry matter along the cycle. Paola reached a total dry matter of 284.79 g and 305.91 g by the plant in cycle I and II, at 1,726 AGDD (106 DAS) and 1,780 AGDD (105 DAS), respectively (Fig. 3). Similar results were reported by Bello and Pinzón (1997) for the "Colombia" diploid potato variety. The authors found a maximum dry matter accumulation in stems, leaves, and floral structures at 107 DAS in altitudinal conditions similar to the present study, although with a different variety. On the other hand, Santos (2010), evaluating the "Guaneña" diploid potato variety, observed contrasting results at 2,572 m a.s.l., reporting a maximum dry matter between 40 and 77 days after plant emergence (DAPE), exceeding 72% of the biomass accumulated by the variety Criolla Colombia.
The leaf, stem, and tuber were the organs that accumulated greater dry matter. Towards the middle of the crop development, tubers become the organ with the highest accumulation, also observed by Santos (2010), between 45 and 55 DAPE. The variety with the highest biomass accumulation in fruits was Paola, with 4.32 and 8.96 g in 2019 and 2020, respectively. At the end of the season, there was a decrease in total biomass in some varieties; this could be related to a cessation of biomass accumulation and the translocation of nutrients toward the landfill organs (Soto et al., 2018).
The amount of biomass in the root is low concerning other organs, which has also been evidenced in other studies (Saldaña and Cotes, 2020). Soto et al. (2018) mention that the stems, leaves, stolons, and roots present exponential growth at the beginning of development, which is related to the relative growth rate behavior, reaching its maximum biomass accumulation between 800 and 900 AGDD. At this point, the exponential growth of tubers is also generally observed for the varieties studied.
The accumulation of dry matter in the aerial organs (leaves and stems) tends to decrease after reaching a maximum weight, except for Paola in 2019, increasing again around 1,700 AGDD, possibly due to temporary storage in the stems. After closing the rows with their canopy, senescence begins in the lower foliage, perhaps indicating the transport of photoassimilates to tubers (Cabezas and Corchuelo, 2005). On the other hand, the dry matter accumulation in the tubers had a rapid growth pattern at the beginning, followed by a plateau period and rapid shift towards the end, more noticeable in the Paola and Primavera varieties (Fig. 3). It has been assumed that towards the end of the crop cycle, when the root system is reduced, the tubers take up nitrogen from the leaves, affecting the longevity of leaves and production (Kooman and Rabbinge, 1996).
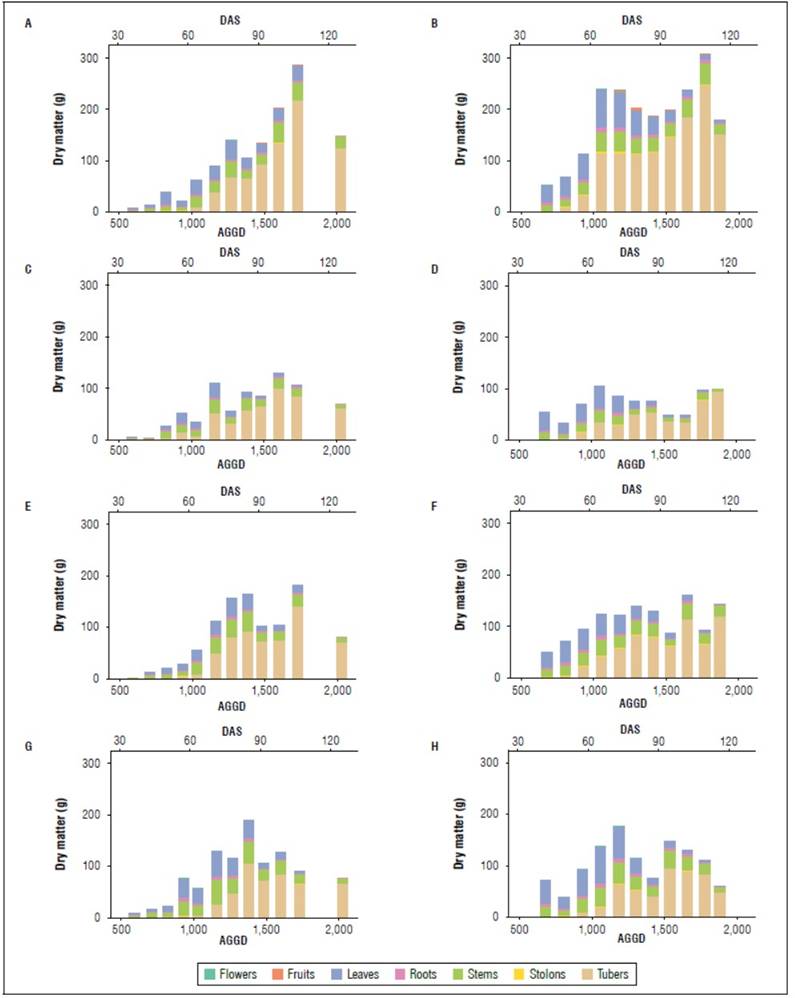
Figure 3. Distribution of biomass throughout the two growing seasons in the organs of the plant (stolons, fruits, leaves, roots, flowers, fruits, roots, and tubers) for the varieties Paola, Paysandú, Primavera, and Violeta. a) 2019 and b) 2020. Accumulated growing degree days (AGDD) and days after sowing (DAS).
The early flowering varieties in the 2019 season was in Paola and Primavera, the last Paysandú. The beginning of tuberization began later in the Primavera variety than in the others. According to Lizarazo (2020), the onset of tuber development occurs around 330 AGDD. It accelerates at the end of the cycle, higher than was found in this study (Tab. 1). For the rest of the variables evaluated, there were no significant differences between varieties. Cabezas and Corchuelo (2005) mention that Solanum phureja, cv. On average flowering stage, Criolla Colombia occurred at 60 DAPE, the maximum tuber activity filling between 75 and 80 DAPE, and the tuber physiological maturity between 90 and 95 DAPE. Sánchez et al. (2005), also in the Criolla Colombia variety, state that the first flower buds originate at 44 DAS.
Table 1. Duration of the flowering, fruit, and tuber development stages according to the BBCH scale, in accumulated growing degree days (AGDD) for the Paola, Primavera, Paysandú, and Violeta varieties.
| Season | Variety | BF | EF | BT | MT | BFG | EFG |
|---|---|---|---|---|---|---|---|
| 2019 | Paola | 946.00 b | 1304.85 a | 855.33 a | 1682.38 a | 1247.87 | 1682.38 a |
| Primavera | 962.10 b | 1304.42 a | 924.92 a | 1782.77 a | 1460.65* | 1483.70 a | |
| Paysandú | 1071.13 a | 1231.42 a | 855.33 a | 1599.58 a | 1377.85* | 1377.85 a | |
| Violeta | 994.40 b | 1316.27 a | 818.05 a | 1410.93 a | 1317.45* | 1493.07 a | |
| 2020 | Paola | 849.10 a | 1323.63 a | 725.50 b | 1780.55 a | 1089.05 | 1285.51 a |
| Primavera | 889.55 a | 1436.65 a | 889.55 a | 1806.56 a | 1436.65 | 1473.88 a | |
| Paysandú | 970.45 a | 1286.85 a | 725.50 b | 1845.85 a | 1436.65* | 1435.71 a | |
| Violeta | 969.53 a | 1360.42 a | 766.70 b | 1663.81 a | 1322.30* | 1285.51 a |
BF: Beginning of flowering (60); EF: End of flowering (69); BT: Beginning of tuber formation (40); MT: Maximum weight of tubers (48). BFG: Beginning fruit growth (70); EFG: End of fruit growth (79). The number in parentheses is the two-digit code assigned by the BBCH scale. Means values with different lowercase letters show significant differences according to the Kruskal-Wallis test (P<0.05). *Maximum value of plants with fruits (50% or more) was never achieved.
Monroy and Reynales (2001) studied diploid potato materials at 2,543 m a.s.l., they stated that the onset of flowering was between 30 and 45 DAS, tuberization between 45 and 70, and fruiting between 85 and 100 DAS. Bello and Pinzón (1997), in the variety Colombia, identified the beginning of tuberization between 51 and 58 DAS, which occurred almost simultaneously with the flowering phase. Santos (2010) reported that tubers accumulate dry matter from 21-28 DAPE and reach the maximum dry matter between 90 and 98 DAPE. Lizarazo (2020) determined the duration in AGDD of the flowering and fruiting development stages (600-609 and 700-749 on the BBCH scale) in the Violeta and Paola variety occurred between 400-450 AGDD and 1,250 AGDD for flowering and 750 and 1,250 AGDD for fruit development, which started earlier concerning this study.
Regarding the duration of reproductive and productive stages for all diploid potato varieties, the flowering, and the tuber formation began earlier, with a more extended period in the 2020 season than in 2019 (Fig. 4). Santos (2010) states that higher altitudes and lower temperatures can increase the phenological stages' start, duration, and completion. Monroy and Reynales (2001) found that flowering in diploid potato clones started up to 40 days earlier at lower altitude locations (2,543 m a.s.l.) compared to higher altitude locations (3,100 m a.s.l.), probably influenced by temperature. For the formation of the tuber, Paysandú stands out, which was the one that required the most AGDD for this phase in the 2019 season (Fig. 4). Regarding fruiting, Paola was characterized by presenting a long fruiting stage (434 and 196 AGDD in 2019 and 2020, respectively), while Primavera had a short duration in this phase (23 and 37 AGDD). However, for the present investigation's conditions, the Paysandú variety did not produce fruits in the first cycle and very few in the second (Fig. 4). The stage of flowering development is essential since it can be related to the elongation of the stolons and the beginning of the transport of photoassimilates to the tubers (Cabezas and Corchuelo, 2005).
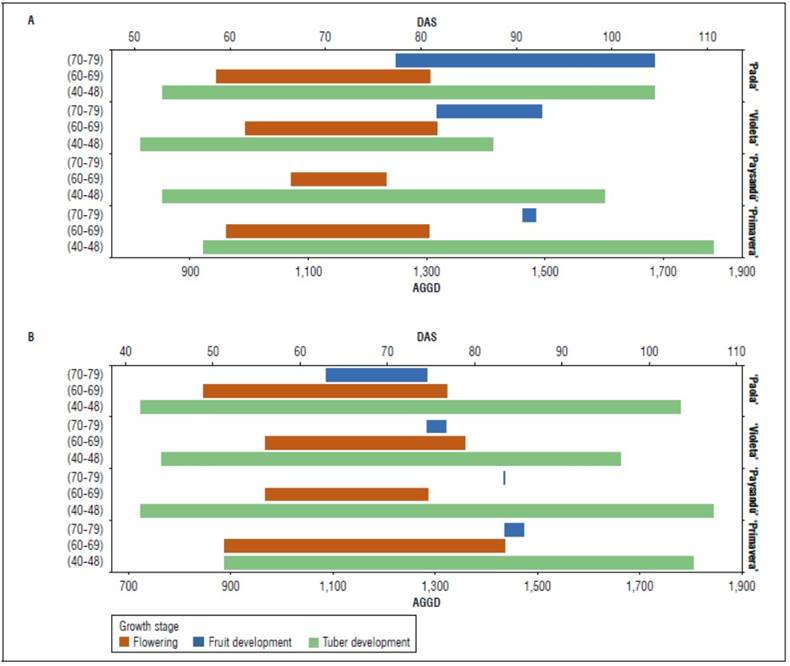
Figure 4. Duration of the flowering, fruit, and tuber growth stages in accumulated growing degree days (AGDD) for both seasons (2019 and 2020) according to the BBCH scale.
The main stems and aerial growth axes were 5 and 6 in the cultivar Paola, 5 and 5 in Primavera and Violeta, and 4 and 3 for Paysandú, respectively, for 2019 and 2020. This variable remained relatively constant during the crop cycle in all varieties (Fig. 5). Regarding the number of tubers and stolons, Paola produced the highest quantity in each organ. According to Haverkort and Struik (2015) the number of sprouts that will develop depends on how many stems there will be produced per seed, and how many stems will survive and become tuber-bearing will depend on variety, seed tuber size, seed tuber health, physiological age, and environmental conditions. Vice versa, the number of above-ground organs per plant is mainly determined by the size of the tuber and its condition, and the pre-sprouting period also influences it (Al-Mahmud et al. 2014). In this sense, farmers manipulate the average tuber size and tuber size distribution to reduce seed quality as a yield-limiting factor. However, in the case of potatoes, the influence of the seed's quality affects the crop's yielding ability and goes beyond affecting the initial growth rate and vigor.
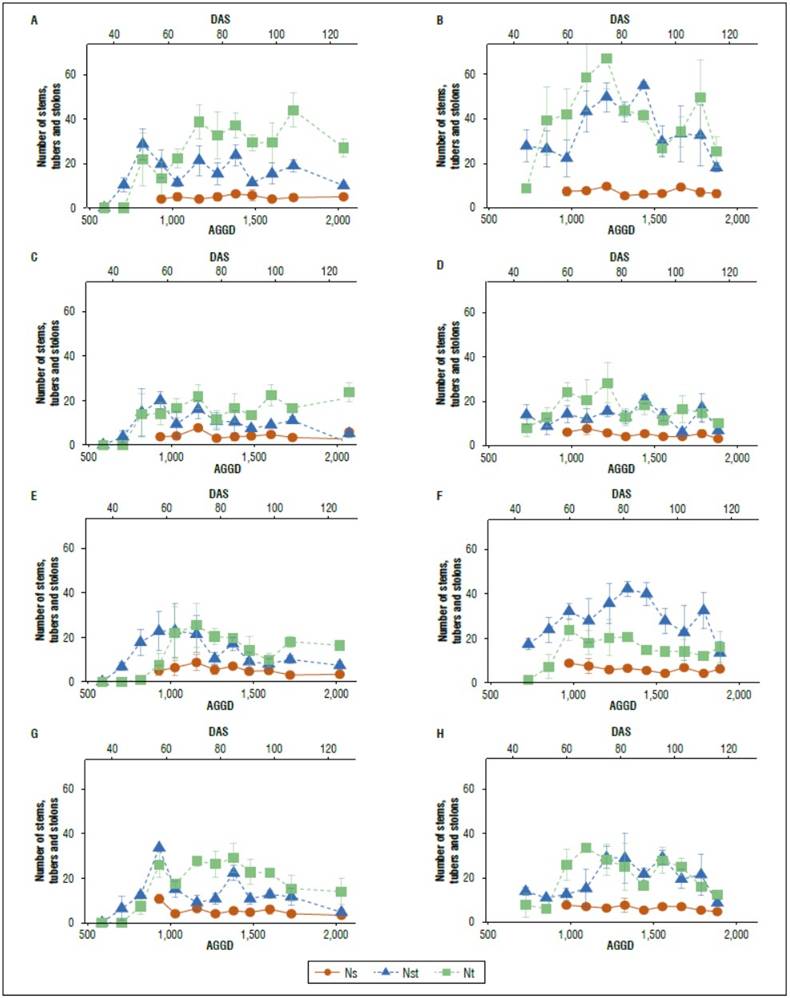
Figure 5. The number of stems (Ns), tubers (Nt), and stolons (Nst) for Paola, Paysandú. Primavera and Violeta varieties. a) 2019 and b) 2020. Data are mean ± standard error.
‘Paola’ required 1,726 AGDD (106 DAS) and 1,222 AGDD (70 DAS) to reach the maximum number of tubers, while ‘Primavera’ needed 1,158 AGDD (71 DAS) and 970 AGDD (56 DAS) for the 2019 and 2020 growing seasons, respectively (Fig. 5). Soto et al. (2018) state that stolons increase their biomass until 900 AGDD and reduce their emission rate to give way to the growth of tubers, similar to what was found in the 2019 season. Bello and Pinzón (1997) registered a growth of roots and stolons before 33 DAS and up to 51 and 58 DAS.
Regarding the NAR and RGR, a series of fluctuations occurred throughout the crop-growing cycle, observing a relationship between both indices. In RGR, a slightly decreasing trend is marked with an increase towards the end (Fig. 6C and 6D), at which time the NAR also increases (Fig. 6A and 6B). Saldaña and Cotes (2020) highlight that in several studies, the behavior of NAR tends to decrease over time. At the end of the cycle, an increase is observed, which could be associated with greater photosynthetic activity and more significant accumulation and distribution of biomass towards the tubers. An adequate NAR is related to good tuber production (Cabezas and Corchuelo, 2005). High RGR values at the beginning of the growing cycle could be associated with the rapid growth of leaves and lateral shoots to capture more photosynthetically active radiation (Santos, 2010). In both seasons, the leaf area ratio (LAR) decreased towards the end of the growing period for all diploid potato varieties. However, the maximum LAR was observed in the 2020 season. Santos et al. (2010) found that the materials that reached the maximum LAR faster developed their photosynthetic apparatus more quickly and had a shorter growing period. In the 2019 season, the Paysandú variety presented the lowest LAR among all the varieties (Fig. 6E), which could relate to the one with the lowest leaf area and yield per plant (Fig. 2, Tab. 2).
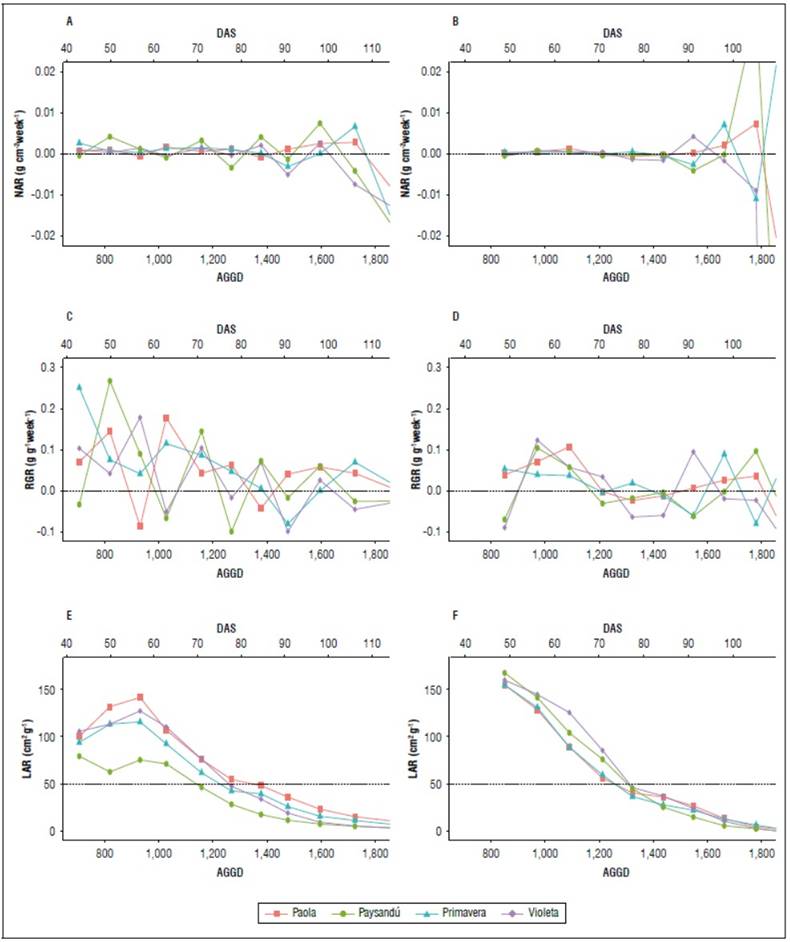
Figure 6. Growth rates during the growing period in the varieties studied: a) and b) Net assimilation rate (NAR); c) and d) Relative growth rate (RGR) and e) and f) Leaf area ratio (LAR) in the 2019 and 2020 season, respectively. Accumulated growing degree days (AGDD) and days after sowing (DAS).
In both growing seasons, a correlation is found between the power of the source and demand in all varieties. The Pearson correlation coefficient between these two variables was 0.66 and 0.98 in Paola, 0.81 and 0.85 in Primavera, 0.69 and 0.7 in Violeta, and 0.88 and 0.79 in Paysandú for the 2019 and 2020 seasons, respectively. According to Santos et al. (2010), a high translocation of photoassimilates causes a high photosynthetic rate in many plants. The demand's power is defined as the tubers' capacity to compete for assimilation with other organs (Kooman and Rabbinge, 1996). The source power is the capacity of an organ to produce photoassimilates (Santos, 2010).
In the Paola variety, high values of the source and demand were presented more continuously than in the other varieties (Fig. 7A and Fig. 7B), which could be related to its higher yield per plant in both cycles (Tab. 2). The moments when the power of the demand was higher than the source could be due to temporary storage in the stems (Santos et al., 2010), as evidenced in the Paola and Primavera variety towards the end of the production cycle and the other varieties in some moments. Paola's highest source and demand power was at 1,089 AGDD (63 DAS) in 2020, with 20.97 and 20.53 g/week, respectively, showing high coordination between the source and the demand. High coordination has agronomic implications due to leaf phytosanitary problems that can influence the yield (Santos et al., 2010).
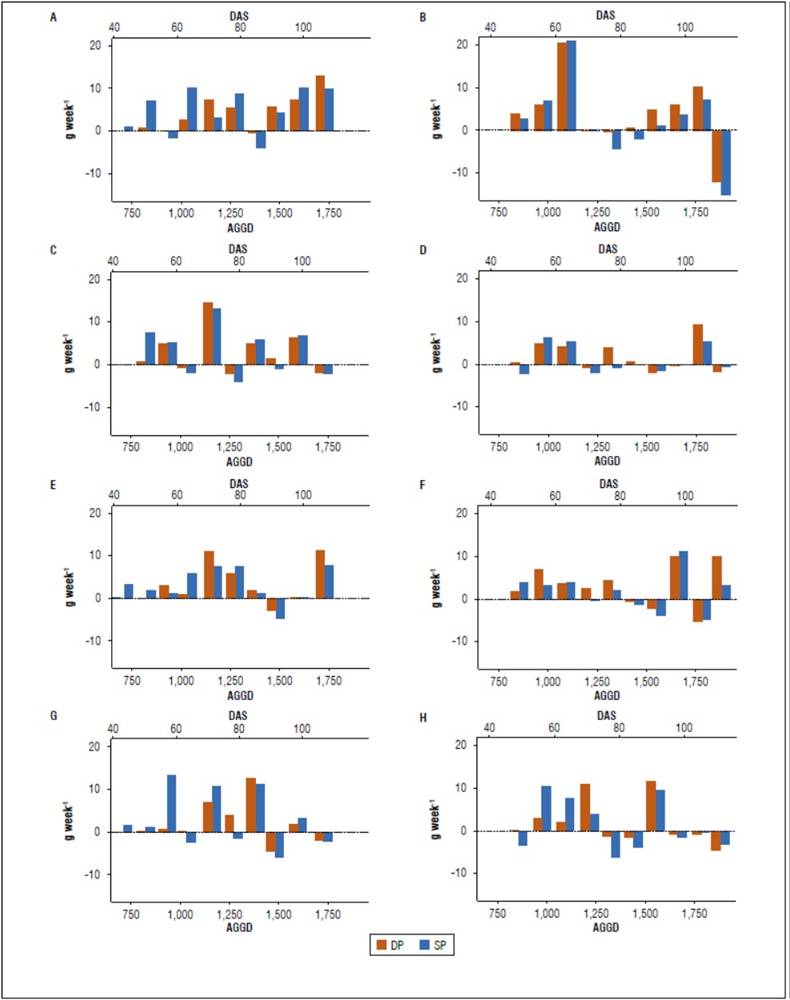
Figure 7. Source power (SP) and demand (DP) of each variety during the crop cycle. a) 2019 and b) 2020. Accumulated growing degree days (AGDD) and days after sowing (DAS).
Regarding tuber yield, in both production cycles, the Paola and Primavera varieties differed significantly in the number of tubers per plant, average weight per tuber, and yield per plant for the Violeta and Paysandú varieties (Tab. 2). Lizarazo (2020) highlighted the Violeta variety as the lowest production evaluated. It is reported that the Paola and Primavera varieties have a productive potential between 600 and 1,000 g of tubers per plant, while Paysandú is less than 600 g and Violeta is greater than 1,000 g per plant (Ñústez, 2018). According to the present work results, the Paola variety achieved similar yields in both cycles to those reported for this genotype. However, Primavera, Paysandú, and Violeta presented yields much lower than their productive potential. This yield decrease may be because no fungicides were applied during the growing cycle in this study. The harvest index observed in these varieties could highlight their precocity compared with varieties of Solanum tuberosum ssp. andigena, which in other studies for 9 to 11 weeks found a CI below 50% (Santos, 2010).
Table 2 Yield and yield components of the Paola, Primavera, Paysandú, and Violeta varieties for the 2019 and 2020 growing seasons.
| Cycle | Variety | Tubers number (plant) | Tuber weight (g)* | Yield (g/plant) | DM (%) | HI (%) |
|---|---|---|---|---|---|---|
| 1 | Paola | 27.75 a | 24.28 a | 691.86 a | 23.23 a | 83.56 a |
| Primavera | 19.00 ab | 23.27 a | 418.22 a | 19.27 a | 81.93 a | |
| Paysandú | 11.75 b | 16.85 b | 184.79 b | 21.40 a | 86.56 a | |
| Violeta | 14.33 b | 17.51 b | 249.85 b | 19.98 a | 80.93 a | |
| 2 | Paola | 24.66 a | 20.27 b | 519.83 a | 26.07 a | 85.18 a |
| Primavera | 17.41 b | 25.86 a | 456.74 a | 21.95 a | 74.33 a | |
| Paysandú | 11.50 c | 19.41 b | 221.78 b | 24.68 a | 91.71 a | |
| Violeta | 12.75 c | 16.93 b | 215.13 b | 22.97 a | 76.83 a |
DM: Percentage of dry matter in tubers; HI: Harvest index (%). Means values with different letters show significant differences according to the Kruskal-Wallis test (P<0.05).
In the last measurements of the Violeta and Paysandú variety and the last one of Paola in the second growing season (Fig. 7), a decrease in the power of the source and demand was observed, which is related with the decrease in the demand for assimilation by the tuber, since this organ reaches physiological maturity. On the other hand, the reduction in the source is associated with the age of the leaves, which, although at the time of harvest, occurs without foliage senescence, the photosynthetic is reduced at the end of the leaf's life (Santos et al., 2010).
The growth and development of the varieties could be affected by the sowing seasons (2019 and 2020) in which the crop was established. The 2019 season was planted in September 2019 and harvested in January 2020; at this time, for the region under study, there are short days towards November and December, and the lowest temperatures were recorded (Fig. 1).
Instead, during the 2020 season, between March and July 2020, there is a transition from short to long days in the northern hemisphere with the highest temperatures (Fig. 1). For optimal yields, potato crops need long days during vegetative growth and short days during tuberization. On the other hand, the main factors that control tuber formation are night temperature and day length (Thongam et al., 2017). The best yield in the first season was associated with short days during the tuberization stage and night temperatures of 12.5°C on average in 2019 and 13.1°C in 2020. Besides growth, the fact that potatoes require long days for better vegetative development, as indicated above, the largest leaf area and the largest number of stolons produced by all varieties were higher in the second season, with concerning the first, with an average temperature of 17.6°C, about that registered in the first cycle of 16.8°C. Therefore, a greater vegetative development relating to the productive one implies a reduction in the landfill source ratio in favor of a greater accumulation of biomass and formation of productive organs such as tubers and, consequently, a reduction in the yield per plant.
CONCLUSION
Season planting affects the tuber yields for some varieties, so this information is necessary to determine the best physiological stages for performing cultural practices and planting time to cultivate diploid potatoes for Phureja group. Paola and Primavera had the highest accumulation of dry matter in each organ and the highest yield of tubers per plant, however, Violeta and Paysandú previously reached the greatest accumulation of matter. The NAR and RGR had a series of fluctuations during the growing season, although they had some correlation between them with an increase towards the end of the growing period. The source and sink power presented a correlation in general for all varieties.

