











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Nowadays, several anthropogenic pressures are jeopardizing animal populations (CEBALLOS et al., 2017). Environmental changes caused by landscape transformation may disrupt homeostasis affecting individual fitness of vertebrate populations (CODA et al., 2016; LAZIC et al., 2013). Particularly, livestock systems have been increasing in the last few decades and is likely to increase in the future, threatening biodiversity in developing countries (THORNTON et al., 2009) . However, studies assessing the stress of animal species in livestock systems are scarce, especially in some Neotropical countries such as Colombia.
One of the approaches to evaluate the degree of stress of animal species is the Fluctuating asymmetry (FA), which is defined as the random deviations from perfect symmetry (e.g. radial, dihedral, rotational, bilateral) in populations of organisms (GRAHAM et al., 2010) . FA can serve as a useful indicator of developmental instability, reflecting environmental and genetic stress of organisms (LEARY and ALLENDORF, 1989). For instance, species under environmental stress such as decreased food availability (WAUTERS, 1996), agrochemicals (CODA et al., 2016), and pathogens (ST- AMOUR et al., 2010), have higher levels of asymmetry. In addition, the decline of heterozygosity has an inverse relation to the levels of FA; in this sense, as heterozygosity decreases, FA increases (MESSIER and MITTON, 1996). Accordingly, FA seems to provide an adequate measure of stress since it has been useful in the evaluation of anthropogenic pressures on different animal groups such as insects (FLOATE and FOX, 2000), birds (LENS et al.,1999), amphibians (ST-AMOUR et al., 2010), reptiles (LAZIC et al., 2013) and mammals (CODA et al., 2016; WAUTERS, 1996).
Within mammals, bats are the unique taxonomic group with the ability for powered flight (CAMARGO and OLIVEIRA, 2012). To achieve this, bats have an integrated and multivariate fight apparatus: the wings (NORBERG and RAYNER, 1987). This structure is a special modification of forelimbs, characterized by a membrane of skin (dactylopatagia), stretched between elongated digits (digits II-V) (Wang et al., 2010). Although some studies have analyzed morphological differences in bat wings according to ecological characteristics (MARINELLO and BERNARD, 2014; NORBERG and RAYNER, 1987), differences in both intraspecific level and wing regions need more attention (STEVENS et al., 2013; CAMARGO and OLIVEIRA, 2012; FIGUEIREDO et al., 2015). For example, variation between sexes in terms of wing traits (i.e. sexual dimorphism) and variation among digits are poorly known.
The Caribbean region of Colombia is one of the most biodiverse in the country (RANGEL-CH, 2015). A priority ecosystem for conservation in this region is the tropical dry-forest, due to the historical loss of 92% of its original coverage (GÓMEZ and MORENO, 2016). Moreover, according to IUCN's Red List of Ecosystems criteria, it has been classified as critically endangered ecosystem (CR) (ETTER et al., 2015), and one of the mainly factors of its historic transformation is the expansion of livestock systems (PIZANO and GARCIA, 2014).
Particularly, in Cordoba department different studies have been done with bats in fragments of tropical dry-forest immersed in matrices of extensive livestock systems, analyzing ectoparasites load (CALONGE, 2012), reproductive phenology (VELA-VARGAS, 2013), and the structure of bat assemblages (BALLESTEROS-CORREA, 2015). In these studies, Artibeus lituratus, Artibeus planirostris, and Carollia perspicillata are the species with the highest abundances in the assemblages. Furthermore, these species are involved in key roles such as seed dispersers of pioneering plants (e.g. Ficus, Cecropia, Solanum), enhancing the reforestation of disturbed areas in tropical dry-forest (RÍOS-BLANCO, 2010). Nevertheless, it is unknown the degree of environmental stress that these bat species are facing in extensive livestock systems, which are quite common in the Caribbean region. The use of FA can be an important tool for this purpose, especially in key structures like bat wings. Therefore, the present study aimed BLANCO, 2010; OLAYA-RODRÍGUEZ, 2009), master (CALONGE, 2012; VELA-VARGAS, 2013) and doctoral thesis (BALLESTEROS-CORREA, 2015).
Specimen selection. Exclusion criteria were used in order to avoid noise factors such as developmental stage, misleading taxonomic identification, and specimens badly preserved. In doing so, bat vouchers selected were only adults, according to data of the tags and field notebooks of each specimen. The identification of each specimen was corroborated with the keys of GARDNER (2007). All specimens in bad state of preservation (i.e. broken wings) were excluded from data analysis.
Wing traits. To analyze FA, 11 wing traits were measured from both right and left sides (Figure 1). The measurements were taken with a digital caliper (Mitutoyo Calibrador Vernier Mod Cd6 -csx 150 Mm) with an accuracy of 0.01 mm and were done by the same researcher (Dennis Castillo-Figueroa).
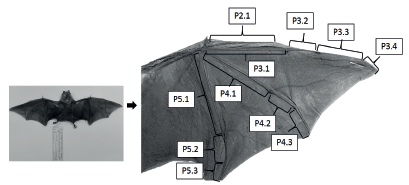
Figure 1 Wing traits used for fluctuating asymmetry analysis. P2.1 Second digit, metacarpal. P3.1 Third digit, metacarpal. P3.2 Third digit, first phalanx. P3.3 Third digit, second phalanx. P3.4 Third digit, third phalanx P4.1 Fourth digit, metacarpal P4.2 Fourth digit, first phalanx P4.3 Fourth digit, second phalanx P5.1 Fifth digit, metacarpal P5.2 Fifth digit, first phalanx P5.3 Fifth digit, second phalanx.
Data analysis. After testing the assumptions of normality (Shapiro-Wilk test) and homoscedasticity (Levene test), wing traits data were nonparametric. Hence, to assess the existence of fluctuating asymmetry between the right and left sides of bat wings, nonparametric U test of Wilcoxon-Mann-Whitney was applied. The null hypothesis for this test establishes no significant differences between the means of wing sides for each trait (FIGUEIREDO et al., 2015). Statistical means and standard deviations are presented for right and left sides of each wing trait.
To evaluate the existence of significant differences among digits of bat wings, the traits were grouped into the following categories: III digit (P3.1+P3.2+P3.3+P3.4), IV digit (P4.1 + P4.2 + P4.3) and V digit (P5.1+P5.2+P5.3). These categories present metacarpals and phalanges that support the main structure of dactylopatagium, which is important for flight performance (NORBERG and RAYNER, 1987). Therefore, a general analysis of bat wing asymmetry was conducted, through the three principal digits that support this wing membrane. The asymmetry value was calculated according to PALMER (1994):
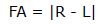
In the formula, FA is the fluctuating asymmetry value, R is the measurement of the right side of the wing, as well as L represents the left side of the wing. Nonparametric one-way analysis of variance was done (Kruskal-Wallis), taking the digit as the factor and the III, IV and V digits as the levels of the factor. In all the tests, p<0.05 defined the level of statistically significant difference. All the analysis presented here, were done separately for males and females in the statistical software Rwizard 2.1. Because of some works have shown sexual dimorphism in Neotropical bats based on wing traits (CAMARGO and OLIVEIRA, 2012; STEVENS et al., 2013), multivariate analysis of variance (MANOVA) was conducted in InfoStat 1.0. Due to wing traits were nonparametric, transformations of all variables to normal scores (z values) were done in order to use MANOVA test.
RESULTS
In total, 114 bat specimens were analyzed: 33 of A. lituratus (15 males, 18 females), 40 of A. planirostris (15 males, 25 females), and 41 of C. perspicillata (21 males, 20 females). The nonparametric Wilcoxon-Mann-Whitney test only showed the existence of significant differences between wing sides of the first phalanx of the third digit in males (P3.2) and the third phalanx of the third digit (P3.4) in females of C. perspicillata (p<0.05, Table 1). For both Artibeus species there were no statistical differences in any wing trait (Table 2 and 3).
Table 1 Wing traits of Carollia perspicillata specimens in extensive livestock systems (Córdoba, Colombia).
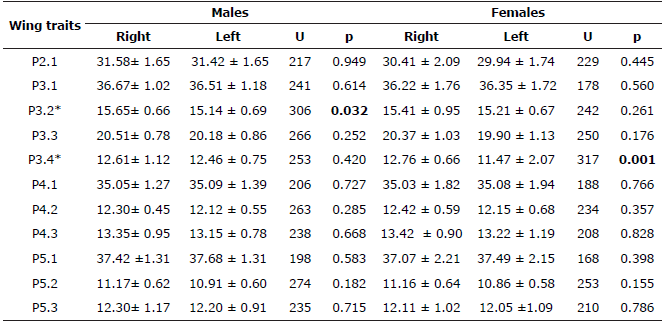
Statistical mean ± standard deviation for Right and Left wings presented in millimeters (mm). U= Non-parametric U test. p= Significance level of Wilcoxon-Mann-Whitney test. * Statistical differences.
Table 2 Wing traits of Artibeus lituratus specimens in extensive livestock systems (Córdoba, Colombia).
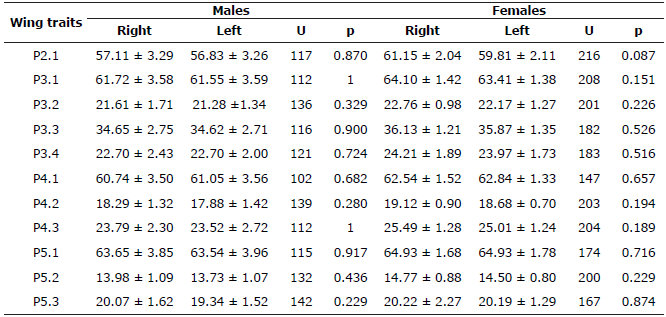
Statistical mean ± standard deviation for Right and Left wings presented in millimeters (mm). U= Non-parametric U test. p= Significance level of Wilcoxon- Mann-Whitney test.
Table 3 Wing traits of Artibeus planirostris specimens in extensive livestock systems (Córdoba, Colombia).
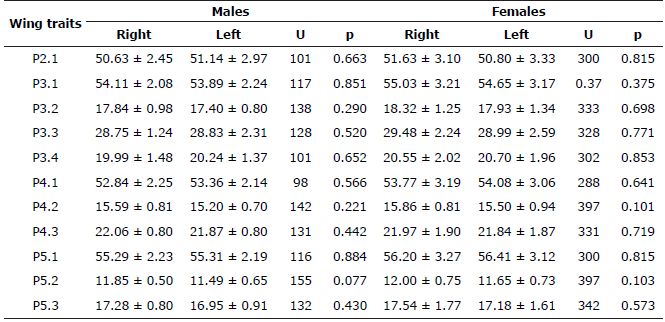
Statistical mean ± standard deviation for Right and Left wings presented in millimeters (mm). U= Non-parametric U test. p= Significance level of Wilcoxon- Mann-Whitney test.
For the digit analysis, only the females of A. planirostris presented significant differences (H= 3.370, p= 0.035, Figure 2). In the other two species, there were no significant difference between the digits in terms of fluctuating asymmetry values (Figure 2).
On the other hand, A. lituratus was the unique of the three bat species analyzed, which showed sexual dimorphism according to wing traits (Wilks' Lambda =0.72, F=2.83, p=0.01). For all the wing measurements, females presented higher values in the statistical means than males (Table 2).
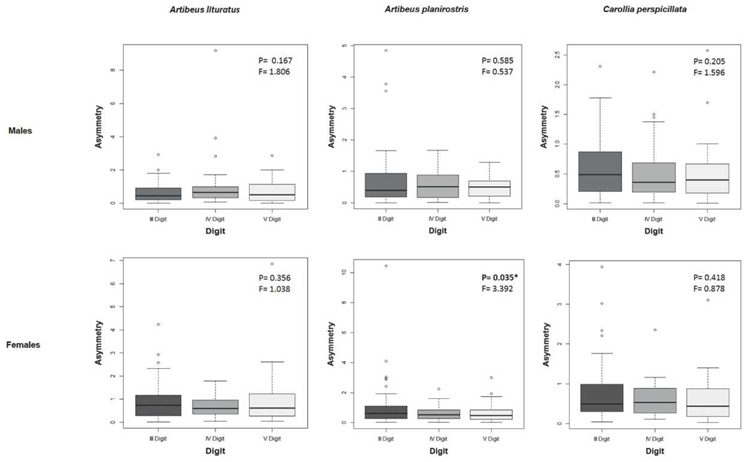
Figure 2 Box-plots for asymmetry values of the III, IV and V digits in females and males of Artibeus planirostris, Artibeus lituratus and Carollia perspicillata specimens in extensive livestock systems (Córdoba, Colombia). p= Significance level of Kruskal-Wallis. H= Kruskal-Wallis test.
By contrast, neither A. planirostris (Wilks' Lambda =0.88, F=0.95, p=0.494) nor C. perspicillata (Wilks' Lambda =0.89, F=0.89, p=0.548) presented sexual dimorphism in wing traits.
DISCUSSION
Fluctuating asymmetry of bat species and their tolerance to habitat perturbations. In general terms, the results showed no statistical difference (p>0,05) in most of the wing traits, suggesting that environmental conditions of tropical dry-forest fragments immersed in extensive livestock systems, were not enough to cause several changes in the wing morphology of these bat species. One of the possible reasons of the low degree of asymmetry is that natural selection may favor symmetrical individuals rather than asymmetrical. The "Differential-mortality" hypothesis establishes that individuals with less FA have more possibilities for survival unlike individuals with high FA (FLOATE and FOX, 2000). Symmetry may favor the locomotion leading a better ability for flight performance and food capture (VOIGT et al., 2005). Hence, the selective pressure on bats eliminates asymmetrical individuals, selecting symmetrical ones for next generations (VOIGT et al., 2005), which are probably the specimens analyzed in this work.
Additionally, variation in wing traits is more conservative due to its functionality on flight performance (NORBERG and RAYNER, 1987), thermoregulation (MAKANYA and MORTOLA, 2007), and food manipulation (VANDOROS and DUMONT, 2004). Comparing with other body regions such as head and hind-limbs, forelimbs have less asymmetry in bats (FIGUEIREDO et al., 2015).
Moreover, the three bat species are common frugivorous with a wide geographical distribution throughout the country (MUÑOZ, 2001). Typically, C. perspicillata and A. lituratus are often used as ecological indicators of fragmented landscapes (GALINDO, 2004). All the three species are reported in different environments such as monocultures (BREVIGLIERI and UIEDA, 2014; ORTEGÓN-MARTÍNEZ and PÉREZ-TORRES, 2007), urban systems (FIGUEIREDO et al., 2015; ALBERICO et al., 2005) and extensive livestock systems (BALLESTEROS-CORREA, 2015). Based on the findings of this research, but also on the high abundances of these bat species reported for extensive livestock systems (BALLESTEROS-CORREA, 2015), it is possible to infer that, at first glance, these species are not hampered by livestock systems because of their high tolerance to habitat perturbation. Further studies should evaluate the FA of bat species from other guilds (e.g. insectivores, carnivores, nectarivores, omnivores), and with other ecological characteristics (e.g. high habitat specificity, rare species), to test differential responses of species to livestock systems.
Some differences in wing traits and sexual dimorphism. Differences were detected in two wing traits of the third digit for C. perspicillata (P3.2, P3.4, Table 1). The third digit is a measure of hand-wing length and enable a fast and economic flight (DIETZ et al., 2006). Recently, it has been found a wing defect (i.e. nonsymmetrical digit) in specimens of C. perspicillata associated to livestock systems (CASTILLO-FIGUEROA and PÉREZ-TORRES, 2018). This wing anomaly refers to the difference of the length in P3.4 between left and right side (CASTILLO-FIGUEROA and PÉREZ-TORRES, 2018), which is the wing trait that showed statistical difference in the present study (Table 1).
For A. planirostris, other studies conducted in urban environments from Brazil neither found a high asymmetry (FIGUEIREDO et al., 2015). The present study shows the low degree of stress for this species in extensive livestock systems, only with statistical difference between the digits of females (Figure 2), but without differences in the individual wing traits for both males and females (Table 3). A. planirostris exhibits a great ecological plasticity using a variety of roost structures in sites with high availability of food (BREVIGLIERI and UIEDA, 2014). In this sense, fragments of tropical dry-forest may provide enough food source for developmental stability in this species.
On the other hand, females of A. lituratus showed for all the statistical means of wing traits, higher values compared with males (Table 2). The "Big Mother" hypothesis proposes that larger females have greater reproductive success, because they can birth larger offspring and bring a greater amount of resources to them (RALLS, 1976). Previously, STEVENS et al. (2013) demonstrated the existence of this phenomenon using wing traits of A. lituratus from 15 sites in eastern Paraguay and five sites in north-eastern Argentina. This study highlighted that larger wing traits are useful for better flight performance in females to forage resources, carrying babies, and compensate a higher body weight (STEVENS et al., 2013). The results presented here support the "Big Mother" hypothesis for A. lituratus in terms of wing traits.
Final considerations. The asymmetry analysis with multiple traits provides better understanding of the influence of stress (ST-AMOUR et al., 2010; LEUNG et al., 2000; LEARY and ALLENDORF, 1989). In this study, detailed analyzes of 11 wing traits were conducted (i.e. metacarpals and phalanges), but also general analyzes using three main structures of the wings were made (i.e. III digit, IV digit, V digit).
One of the limitations of the present study is the low sample size, which can affect the detection of normality deviations (BENÍTEZ and PARRA, 2011; GRAHAM et al., 2010). However, this issue depends mainly on the specimens available in biological collections (CASTILLO-FIGUEROA, in press;PYKE and EHRLICH, 2010), being outside the researcher's control. It is important to increase the number of specimens in museum collections for address, robustly, different ecological questions using more samples. This study shows the relevance of biological collections in the contribution of key data, which can be useful to parse out the effect of landscape transformation on the ecological responses of species (CASTILLO-FIGUEROA, in press).
In conclusion, wing traits of these bat species did not show a considerably asymmetry in tropical dry-forest fragments immersed in extensive livestock systems of Cordoba department. The results of this work suggest a low degree of stress for the three bat species, and highlight that livestock systems do not severely affect them due to their high tolerance to habitat perturbation. Nevertheless, this study did not distinguish the type of management of extensive livestock system. It would be important to compare silvopastoral management with conventional management to know, with more detail, if someone of them provides better conditions for bat species. Finally, it would be important to consider bat species of other guilds and with a high habitat specificity, because in the present study only frugivorous and common species were considered.

