











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
El pelaje es un derivado de la epidermis que es exclusivo de la clase Mammalia, es una característica muy distintiva que abarca una amplia variedad y patrones de coloración. Desde la antigüedad ha despertado interés en los ganaderos y técnicos con el objetivo primordial de identificar las diferentes razas que presentan patrones hereditarios dentro y entre ellos. El color del pelaje es un carácter de herencia de tipo de cualitativo existiendo una gran variación. Hasta el presente se han identificado más de 150 genes que determinan el color del pelaje y cada uno de ellos tienen dos o más variantes que influyen en la pigmentación de diferentes maneras 1).
A pesar de la existencia de tantos genes el número de colores es bastante reducido ya que se pueden identificar únicamente pelos blancos, colorados o rojos con varias tonalidades formando dos grandes clasificaciones, simples y compuestas, de las cuales se derivan 10 grupos que en términos generales incluyen 16 colores 2. La amplia variabilidad y patrones de coloración cumplen un importante papel en la adaptación, protección y supervivencia 1,3,4,5,6,7,8.
La base del color en los mamíferos radica en la síntesis y distribución de la melanina en la piel, pelo y mucosas 9. La melanina se sintetiza en los melanocitos, que se transfieren por exocitosis a los keratinocitos de la piel y a los bulbos pilosos donde se determinan los diferentes patrones de color. Existen dos clases de melanina: eumelanina que se produce si la hormona MSH se une al melanocito y es la responsable de la coloración negra/castaña, y la feomelanina se produce por ausencia de unión de la hormona MSH con el melanocito y es responsable de la coloración roja/amarilla 10,11. La coloración de los mamíferos está determinada por la distribución de estos dos pigmentos, y la cantidad relativa de cada uno de ellos 12,13,14; los fenotipos de la coloración son frecuentemente polimórficos y pueden representar adaptaciones a las condiciones ecológicas cambiantes 14.
Uno de los genes que controla el color del pelo es el gen E que lleva la información para la síntesis del receptor 1 de la hormona MSH; para este gen E se han detectado tres variantes genéticas que dan origen a tres receptores diferentes en la membrana del melanocito. El receptor ED (negro dominante) es el responsable del pelaje negro, típico en Angus, Brangus y Holstein. El receptor e (rojo recesivo) es el responsable del pelaje colorado, típico en Angus, Hereford, Limousine, Brangus, Bradford y El receptor E+ (salvaje o ancestral) es responsable de muchos pelajes diferentes: negro, gris, negro-parduzco, marrón, marrón-oscuro, marón-rojizo, negro-rojizo y rojo. El color de la capa es innato de la raza, pero algunos factores como la edad, el clima o los cruces pueden influir en la tonalidad 1.
Para la raza Sahiwal se han identificado los genes candidatos clave y las regiones genómicas putativas asociadas con rasgos económicos utilizando datos de polimorfismo de un solo nucleótido, los cuales revelaron la selección para la producción de leche, el color del pelaje y la adaptabilidad al clima tropical 15.
El color del pelaje y otras características morfológicas de este tales como la longitud, densidad y peso del pelo se han asociado con parámetros fisiológicos del animal que son indicadores de la adaptación al calor en regiones tropicales y ecuatoriales; en la raza criolla Caqueteña el color claro, la menor longitud, densidad y peso del pelo se asociaron con menores frecuencias respiratorias y baja temperatura superficial comparado con los grupos raciales Bos taurus taurus y Bos taurus indicus, en la Amazonía colombiana 16
En Colombia no existen trabajos que expliquen la herencia del color del pelaje en las razas criollas, por lo tanto, el objetivo de este trabajo es realizar el estudio de la herencia del color del pelaje en el ganado criollo Romosinuano, a partir de una población de animales pertenecientes al hato de conservación y preservación de dicha raza, existente en el C.I Turipaná el cual se ha manejado como hato cerrado desde el año 1936 17.
MATERIALES Y MÉTODOS
Descripción del hato. Los datos utilizados en esta investigación provienen del hato de ganado criollo Romosinuano, del Centro de Investigación Turipaná de la Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA). El Centro se encuentra localizado en el Valle del Sinú en el en el departamento de Córdoba, nordeste de Colombia, a 8° y 49' de latitud norte y los 762 y 27', y 752 y 35' de longitud oeste del meridiano de Greenwich, altura sobre el nivel del mar es de 20 metros 17.
La temperatura promedia es de 28°C, siendo los meses más calurosos diciembre, enero, febrero y marzo. La humedad relativa oscila en un rango entre 79 % y 85 %. El 85% de la precipitación ocurre entre abril y noviembre; las lluvias se distribuyen en períodos definidos, iniciándose en el mes de marzo y aumentando su intensidad en los meses de abril y mayo, con una precipitación promedio anual de 1120-1200 milímetros. Por lo general entre junio y julio se presenta un veranillo, durante el cual hay pocas lluvias, pero de fuerte intensidad, motivo por el cual estos meses aparecen con un total de precipitación elevada. En agosto, septiembre y octubre, continúa el invierno con lluvias intensas, hasta mediados de noviembre, cuando se inicia la época seca. Los valores máximos de índice de carga de calor (LHI) en el C.I. Turipaná indican que, en determinados momentos del día o en épocas específicas, los animales allí ubicados están en riesgo de padecer estrés calórico, el cual es un factor que ha causado disminución de ingesta de los bovinos cebú comercial en sistemas intensivos de producción de carne en el Valle del Sinú 18
Los suelos se consideran entre los más fértiles del Valle del Sinú. El pH varía de 4,0 a 8,2, siendo el promedio 7,0. Existe marcado desequilibrio entre el contenido de Ca y Mg y el P asimilable es bajo.
Manejo del hato. El hato Romosinuano del CI Turipaná desde sus inicios ha sido manejado bajo condiciones de pastoreo, predominando los pastos Angleton (Dichantium aristatum) y el Tanzania (Megathyrsus maximus cv tanzania). Los animales reciben sal mineralizada y agua a voluntad en sus respectivas praderas; durante la época más seca se les suministra ensilaje de maíz (Zea mays).
En la etapa de formación del hato en 1936, se buscaba que lograra estar conformado por un grupo importante de animales. A partir de 1959 fue desarrollado un plan de apareamiento, para evitar la consanguinidad. Durante el periodo comprendido entre los años de 1959 a 1963, el período de apareamiento se realizó durante todo el año siendo más usual en el periodo de enero a noviembre. A partir de 1964 hasta el presente, el período de apareamiento fue restringido a cuatro meses durante el año, con el fin de que la estación de nacimientos coincida con el período seco del año; práctica de manejo que ha permitido obtener una menor mortalidad de los terneros.
Durante la estación de monta el pastoreo es alterno y al finalizar ésta los animales pasan a un sistema rotacional. Cada potrero tiene una extensión de aproximadamente cinco hectáreas.
La edad para exponer a las novillas al toro es aproximadamente de 24 meses. La formación de grupos de apareamiento es hecha mediante familias, teniendo en cuenta la familia de la madre de cada animal. Para controlar la consanguinidad se estableció el apareamiento circular entre vacas y toros, nunca se aparean toros con hembras de la misma familia; el apareamiento se reordena cada tres años.
Los terneros son destetados a los ocho meses de edad aproximadamente. A este tiempo se efectúa la primera elección de los futuros toros de acuerdo con su conformación y peso. Las novillas descartadas son aquellas que no presentan los caracteres propios de la raza. A los 16 meses de edad se pesan de nuevo todos los animales y se escogen definitivamente los machos para el apareamiento del próximo año y se descartan las novillas de poco desarrollo.
Todos los animales recibieron tratamientos curativos y preventivos contra las enfermedades comunes de la región. En ningún caso fueron descartados antes de los 16 meses de edad y ningún macho fue castrado antes de esta edad.
Colección de los datos. Para este trabajo se utilizaron todos los animales del Banco de Germoplasma de Conservación de la raza Romosinuano del C.I Turipaná, los cuales disponían de su genealogía y adicionalmente se determinó el color tanto de los padres como de los hijos, y con base en esto se analizó a qué tipo de herencia cualitativa correspondía. El número total de animales utilizado fue de 325, incluyendo hembras y machos.
Durante el año del 2007 se realizó un registro de los animales clasificándolos por el color de su pelaje, basándose en la clasificación dada en el hato para los colores: bayo, hosco, mono y rojo cereza; para esto se hizo la observación visual de cada animal anotándose en una libreta el número de identificación y el color.
Las variables correspondientes al número del animal, identificación de los padres y el color del pelaje del padre, madre e hijos fueron registrados a una base de Excell y con base en esta información se determinó la frecuencia de cada fenotipo y el tipo de acción génica al que correspondía.
Descripción fenotípica de los colores. A continuación, se describen las características fenotípicas que distinguen los diferentes colores.
Color bayo. Este color se describe como un tono amarillo pálido, presentando dentro de sus características mucosas pálidas en su mayor parte, aunque pueden encontrarse animales con tonos oscuros en el morro y pezuñas, este color es el que presenta mayor despigmentación, presentando piel rosada en la mayoría de los ejemplares, ojos claros, arco orbital rosado con pestañas rubias; además se observa en algunas ocasiones que animales de este color presentan zonas calvas pero que no corresponden a signos de enfermedad de la piel. El morro puede encontrarse en las tonalidades rosadas, grises o negras. El arco orbital presenta un color rosado, con pestañas rubias. Las orejas presentan el mismo color amarillo pálido del resto del cuerpo, con pelos rubios y blanquecinos en su parte interna. El color de los órganos genitales y ubre es de un tono amarillento que contrasta con el resto del cuerpo. Las pezuñas se presentan en los tonos grises y en algunas ocasiones bastante claras. La borla de la cola en el Bayo es de color rubio entremezclado con tonos rojizos (Figura 1a).

Figura 1 a. Ejemplares de la raza Romosinuano de color bayo; b. Ejemplar de la raza Romosinuano de color hosco; c. Ejemplar de raza Romosinuano de color cerezo; d. Ejemplar de raza de la Romosinuano color mono.
Color hosco. Este presenta mayores características específicas, diferenciándose ampliamente ante los demás colores por el negro de la capa de la cara, extremidades y parte baja de la piel del escroto. El morro siempre es negro, al igual que el arco orbital, pestañas y ojos. Los bordes de las orejas presentan pelos negros que pueden estar mezclados con blancos y monos. El ápice de la bolsa escrotal y el prepucio son de color negro. La parte de las rodillas y corvejones hacia abajo son de color negro. Las pezuñas son bien pigmentadas. La borla de la cola es abundante y con pelos negros y amarillos (Figura 1b).
Color rojo cereza. También conocido como rojo chino, presenta una tonalidad uniforme y muy vistosa que gusta mucho por su tono encendido, mostrando una expresión de salud y vigor representado en su capa bien adherida, de pelos cortos y grasosos. El color del morro pude ser negro o gris. El arco orbital en su mayoría es de color negro, pero se observa también despigmentación en algunos animales; las pestañas son rubias totalmente o con algunos pelos negros. Las orejas tienen el mismo color del resto del cuerpo y en algunos casos se pueden observar pelos negros entremezclados en el borde. En la parte baja de alguna de sus extremidades se les pueden observar pelos negros con sus pezuñas negras o grises oscuras. La borla de la cola tiene mechones rubios, rojizos y algunas veces pelos negros (Figura 1c).
Color mono. El color mono es el tono más claro de las variantes del colorado, es una transición entre el rojo cereza y el bayo, con pelos blancos intercalados, combina las características de los dos, este color puede estar uniformemente repartido en todo el cuerpo o presentar en el tren anterior un mono claro. Las orejas presentan una línea de pelo mono y con zonas de pelo blanco en su interior. El morro puede ser de color negro gris o rosado. Las pezuñas son grises o negras. La borla de la cola presenta tonalidades claras como el rubio rojizo y rubio claro con mechones de pelos blancos (pelicano) (Figura 1d).
Análisis de los datos. Para el análisis de los datos, se tomó como base el fenotipo de cada uno de los animales en los cuales se presentaron diferentes apareamientos: bayo x bayo; bayo x hosco; bayo x rojo cereza; bayo x mono; hosco x hosco; hosco x rojo cereza; hosco x mono; rojo cereza x rojo cereza; rojo cereza x mono y mono x mono y de acuerdo con los resultados se determinó el tipo de acción génica que controla el carácter del color del pelaje en la raza Romosinuano.
RESULTADOS
De acuerdo con el fenotipo obtenido de la población de animales analizados (49% de color bayo, el 25% hosco, 15% rojo cereza y 11% mono) se constató que el grado de dominancia es: bayo > hosco > rojo cereza > mono.
La nomenclatura para designar los genotipos correspondientes a los fenotipos del color del pelaje en la raza Romosinuano se describe en la Tabla 1.
Tabla 1 Denotación de la nomenclatura genotípica para los fenotipos de los cuatro colores del pelaje de la Raza Romosinuano.
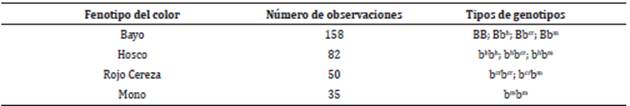
Fuente: elaboración propia
BB: B gen dominante bayo
Bbh: B gen dominante bayo y bh gen recesivo de hosco
Bbcr: B gen dominante bayo y bcr gen recesivo rojo cereza
Bbm: B gen dominante bayo y bm gen recesivo de mono
bhbh; bh gen recesivo hosco
bhbcr: bh gen recesivo hosco y bcr gen recesivo de rojo cereza
bhbm: bh gen recesivo hosco y bm gen recesivo de mono
bcrbcr: bcr gen recesivo rojo de cereza
bcrbm: bcr gen recesivo rojo de cereza y bm gen recesivo de mono
bm bm : bm gen recesivo de mono
En Romosinuano se identificaron cuatro fenotipos, el bayo, hosco, rojo cereza y mono, los cuales pueden presentar cuatro, tres, dos y un genotipo respectivamente, tal como se indica en la Tabla 1. Estos genes ocupan el mismo locus en cromosoma homólogo afectando el color del carácter de dominancia, por lo tanto, se denominan alelos múltiples.
DISCUSION
Teniendo en cuenta los principios de la genética clásica, ya se conoce que cuando existen dos caracteres y el tipo de acción es de dominancia; al aparear individuos heterocigotos la proporción es, por cada tres individuos dominantes, existe uno con el carácter recesivo (–: ¼).
La dominancia incompleta se caracteriza porque el heterocigoto se diferencia de los homocigotos, presentan un tipo de proporción genotípica y fenotípica iguales al aparearse dos heterocigotos, siendo esta: 1/4: 2/4: 1/4.
En los cruzamientos cuando el efecto genético es de codominancia, al aparearse una gran cantidad de animales heterocigotos los dos genes alelos se expresan, pero sin unirse. Por ejemplo, en la raza Shorthorn al aparear vacas rojas con toros blancos, la F1 es ruana y al aparearse entre sí una gran cantidad de individuos de color ruano se espera que se produzcan una proporción es de % rojos: % ruanos: % blancos.
De acuerdo con las acciones genéticas analizados anteriormente existen dos y/o tres fenotipos, respectivamente; estos efectos quedan descartados con relación al tipo de acción génica que gobierna el color del pelaje en la raza Romosinuano, ya que existen cuatro fenotipos distintos.
Si se tienen en cuenta dos caracteres que se heredan independientemente y en donde el uno es dominante y el otro es recesivo, del apareamiento de muchos individuos heterocigotos la proporción resultante es 9/16: 3/16: 3/16: 1/16. Cuando el tipo de acción génica es de dominancia y existe un apareamiento entre dihíbridos, se obtienen como resultado cuatro fenotipos distintos como en el caso del color del pelaje de la raza Romosinuano, pero entre los diferentes apareamientos no se ocurre la proporción esperada de 9:3:3:1; por lo tanto, este tipo de acción génica también queda descartado.
La epistasis se caracteriza por poseer tres fenotipos con las siguientes proporciones: 12/16: 3/16: 1/16; 9/16: 4/16: 3/16 o 9/16: 6/16: 1/16; o puede presentar también dos fenotipos con las siguientes proporciones: 9/16: 7/16; 15/16: 1/16 o 13/16:3/16. Para el color del pelaje en la raza Romosinuano, no se presentan ni tres, ni dos fenotipos; por lo tanto, este tipo acción génica, también queda descartada.
El color del pelaje de los bovinos es independiente del sexo; ya que tanto machos como hembras presentan cualquier color y, por lo tanto, se puede concluir que el tipo de acción génica bien sea ligado, limitado o influenciado por el sexo, no afecta dicho carácter.
El tipo de acción génica de alelos múltiples se caracteriza porque de diferentes apareamientos se presentan cuatro o más fenotipos diferentes y, además, existe cierto grado de dominancia.
Al analizar los apareamientos entre animales bayos entre si, se encontró que dichos padres podrían originar hijos de color: bayo, hosco, rojo cereza o mono; que cuando los padres eran hoscos, la descendencia podría tener color: hosco, rojo cereza o mono; si los padres eran de fenotipo rojo cereza, sus progenies podrían ser: rojo cereza o mono y cuando los padres eran de color mono siempre sus hijos eran de color mono. De lo anterior se deduce que existe un grado de dominancia; ya que el apareamiento entre bayos puede producir cualquier tipo de color, los hoscos pueden dar hoscos y dos colores adicionales; el apareamiento entre animales rojo cereza originó individuos rojos cereza y/o mono y el apareamiento de individuos mono entre sí solo originaron animales de color mono; por lo tanto, el tipo de acción génica que gobierna dicho carácter es de alelos múltiples.
Con base en los fenotipos de la población evaluada, puede concluirse que el color del pelaje en la raza Romosinuano, es un carácter cualitativo y hereditario, gobernado por alelos múltiples, siendo su orden de dominancia es bayo> hosco>rojo cereza > mono.
Se recomienda desarrollar estudios relacionados con los diferentes factores, que caracterizan la piel de la raza Romosinuano, como cantidad de glándulas sudoríparas y sebáceas, y espesura de la piel, la cuales posibilitan la adaptación al medio. Establecer estudios que ayuden a comprender la alta eficiencia reproductiva del Romosinuano, en el medio donde se desarrolló.

