











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Historically, the livestock production model practiced by most South American ranchers has a strong extractive aspect with little concern for protecting and renewing the natural resources. This absence of the use of technical criteria in the utilization of natural resources results in accelerated degradation of pasture areas, where degradation is mainly characterized by loss in the productive capacity of the forage grass due to the severe loss of soil fertility and the increase in weed infestation (Lima and Pozzobon 2005). Renewal of the area is the most rational solution when a pasture becomes degraded. Renewal consists basically of destroying the old vegetation, correcting soil fertility and planting the appropriate forage species for the local conditions, usually an exotic (introduced) species (Macedo 2009). The process of renovating the degraded pasture, however, is ineffective in removing the seeds left by weeds, so weed and pasture seeds germinate together, which initiates a new degradation cycle (Martins et al. 2007).
Brachiaria grass (Brachiaria brizantha, now Urochloa brizantha) is one of the most cultivated forages in the warm regions of South America and supports a large portion of the cattle herd. It has high forage yield, persistence, good capacity for regrowth and relative tolerance to attacks from spittlebugs such as Deois sp. The average annual productivity is 4,000-8,000 kg DM/ha and can reach 20,000 kg DM/ha (Benett et al. 2008). It is considered an excellent source of good quality feed when appropriately fertilized and managed. However, nutritional value declines rapidly following flowering (Valle et al. 2000).
All studies we found in the literature on weed control in pastures focused on studying the effects of herbicides on controlling unwanted plants (Silva et al. 2005; Santos et al. 2007; Trigueiro et al. 2007). Practically no attention has been given to the study of interference relations between weeds and grasses, especially with regards to productivity and carrying capacity of the pasture.
This work sought to study the effects of increasing periods of weed presence on the initial development of Brachiaria brizantha cv. Marandu forage.
Materials and Methods
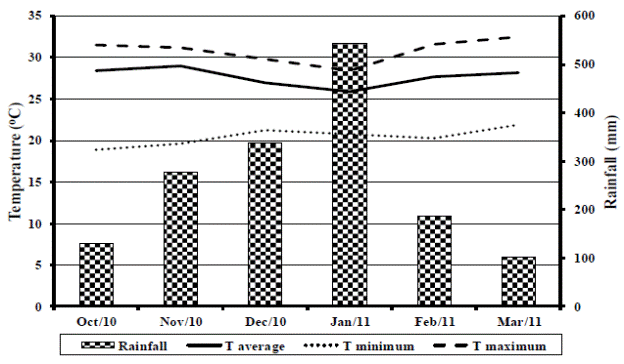
The experimental phase of this work was conducted in a pasture renewal area in the municipality of Sinop, Mato Grosso, Brazil, (11°11'29" S, 55°15'13" W), where, according to the Kõppen (1948) classification, the climate is of type Aw. Rainfall data and average, minimum and maximum temperatures recorded during the experimental period are shown in Figure 1. The average annual temperature is 27 °C, varying between 17 and 40 °C. Average annual precipitation is 1,500 mm, varying from 1,200 to 1,800 mm.
Representative soil samples (latosol) were collected and sent for laboratory analysis, which revealed the following chemical characteristics: pH (H2O): 3.9; soil organic matter: 18.45 g/dm3; P: 0.04 g/dm3 (Mehlich-1); K+: 0.04 cmmol/dm3; Ca2+: 0.09 cmmol/dm3; Mg2+: 0.03 cmmol/dm3; Al3+: 0.3 cmmol/dm3 (KCl 1 mol/L); base saturation: 0.16 cmmol/dm3; and effective CEC of 21%. Physical analysis indicated 866 g sand/kg, 39 g silt/kg and 91 g clay/kg, characterizing the soil as having a sandy texture. Based on these results, calcium lime was applied in the third week of September 2010 at rates of 2,000 kg/ha, while 62.5 kg P/ha (as single superphosphate), 25 kg N/ha (as urea) and 37.5 kg K/ha (as KCl) were broadcast on all areas in the first week of November 2010.
Existing forage was removed from the experimental area by herbicide application (glyphosate at 2.5 L/ha) followed by mechanical tillage of soil before the study commenced in November 2010. The area was fenced to exclude animals for the duration of the study. Seed of cv. Marandu with 70% maximum germination was broadcast on the area at a rate of 6.0 kg/ha. Seedling emergence commenced in 7 days and complete emergence occurred by 10 days. The experimental area was divided into 32 plots, each of 16 m2 (4.0 x 4.0 m) with the central 9.0 m2 of each plot used as the sampling area. The experimental design was a complete randomized block, with 8 treatments representing different periods of coexistence between forage and weed species (0, 15, 30, 45, 60, 75, 90 and 120 days after emergence) and 4 replications.
Assessments of the pasture communities for each treatment were performed at the end of the coexistence period for that treatment with the aid of a 1.0 m2 metal square randomly cast within the sampling area of each plot. All weed species within the metal square were separately identified, counted, cut at stem base and taken to the laboratory, where they were placed in properly labeled and perforated paper bags to be dried in a forced-air oven at 60-63 °C for 72 hours. After this procedure, the dry weight of the stems and leaves of each collected species was determined by using a 0.01 g precision balance.
The relative importance (RI) of the weeds was calculated by the formula:
RI = (IVIe/IVIt) x 100 (%), where: IVIe refers to the importance index of a determined species; and IVIt signifies the sum of the importance indices of all components of the community. The importance index of each species is estimated by the formula:
IVI = DeR + DoR + RF, where: DeR refers to the relative density of each species, estimated by the formula:
DeR = (De/Dt) x 100 (%), where: De is the density of each weed species and Dt is the total weed density; DoR refers to the relative dominance of each species and is estimated by the formula: DoR = (DMe/DMt) x 100 (%), where: DMe is the dry matter of each weed species and DMt is the total dry matter accumulated by the weed community; and RF refers to the relative frequency of each species and is estimated by the formula: RF = (FAe/FAt) x 100 (%), where: FAe is the absolute frequency of each species calculated by the expression: FAe = (NAe/NAt) x 100, where: NAe indicates the number of samples for a determined species; NAt is the total number of samples obtained; and FAt is the sum of the absolute frequencies of all species of the weed community (Mueller-Dombois and Ellenberg 1974).
All weeds were removed from the respective plots at the end of each period of coexistence by spraying with herbicide, and thereafter any emerging weeds were removed by applying 1.5 L/ha of herbicide formulated with 40 g acid equivalent/L of aminopyralid and 320 g acid equivalent/L of 2, 4-D post-emergence.
The grass was evaluated only at the end of the experimental period, corresponding with 120 days after emergence of the seedlings, when the first grass inflorescences emerged. At this time plant height, number of tillers per plant and number of plants per square meter were measured. Forage samples were collected by cutting the plants to 10 cm from the ground within the area enclosed by the 1.0 m2 metal square, cast randomly in the sampling area of the experimental unit.
The samples were sent to the laboratory and sorted into green leaves, green stems and dead matter. The green inflorescences present were considered as part of the stem. The various fractions were duly packed in properly labeled and perforated paper bags and dried in a forced-air circulation oven at 60-63 °C for 72 hours. Dry matter (DM) yields for the different fractions [green leaf dry matter (GLDM), green stem dry matter (GSDM), dead material dry matter (DMDM) and total dry matter (TDM)] were calculated. Dry matter yields for the different treatments were compared with those for the 0 days treatment (control, = weed-free throughout) to determine the suppression in yield by exposure to weeds for the various times according to the formula:
ROF = [(MST - MSPC)/MST] x 100, where: ROF is the reduction in forage offering in percent; MST is the total DM produced by the forage species that remained weed-free for 120 days; and MSPC is the DM produced by treatments with differing periods of weed infestation.
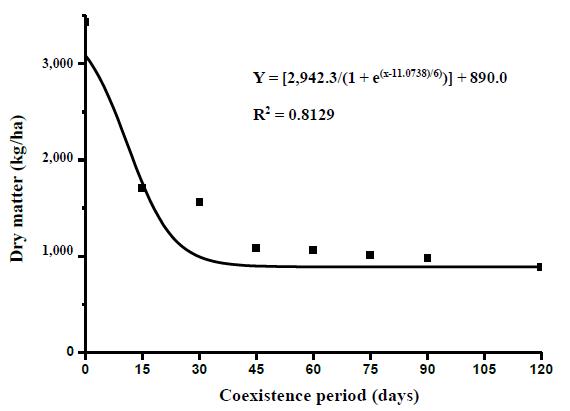
The values obtained were submitted to analysis of variance by the F-test and the effects of the treatments were compared by the Scott-Knott test at 5% probability. The average values of total DM produced by Marandu and observed at 120 days were also adjusted according to the Boltzmann model for better understanding of the effects of coexistence, as used by Kuva et al. (2001). This model conforms to the following equation:
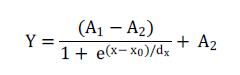
where: Y is the estimated DM yield of the forage in g/m2; x is the upper limit of the coexistence period or control considered; A1 is the estimated yield obtained in the plots maintained clean throughout the cycle; A2 is the minimum estimated production obtained in the units maintained with weeds throughout the cycle; xo is the upper limit of the control or coexistence period corresponding to the intermediate value between maximum and minimum production; and dx is the parameter that indicates the velocity loss or production gain (tg a at point xo).
Results
Only 3 weed species emerged during the experimental period: Hyptis suaveolens (L.) Poit. (Lamiaceae) (local name: cheirosa), Senna obtusifolia (L.) H.S. Irwin & Barneby (Leguminosae - Caesalpinioideae) (local name: fedegoso) and Sida rhombifolia L. (Malvaceae) (local name: guanxuma). Senna obtusifolia accumulated the greatest amount of DM up to 45 days after emergence (DAE), but was the only species that showed a reduction in DM accumulation at 120 DAE (Figure 2). Over the full 120 days, H. suaveolens showed the highest DM accumulation with rapid growth from 45 to 120 DAE; DM accumulation was almost 2.5 times that of S. obtusifolia (Figure 2).
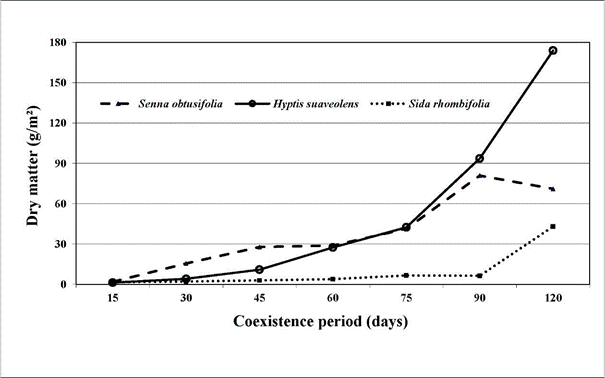
Sida rhombifolia produced little growth up to 90 DAE, when compared with the other 2 species, but accumulation of DM increased considerably from 90 DAE. At 120 DAE total DM produced by the weeds was 288 g/m2 (Figure 2).
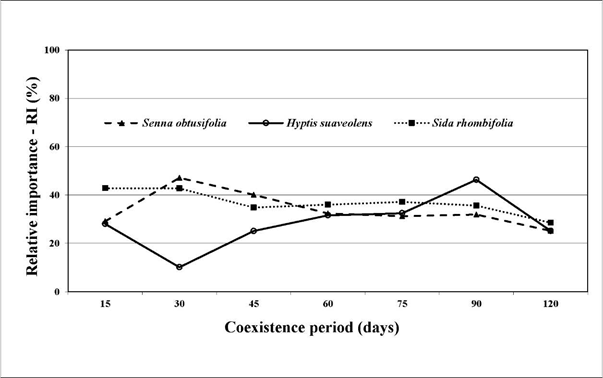
The relative importance, obtained from density, dominance and frequency of each of the 3 species, was relatively equal throughout the experimental period (Figure 3).
The height of Marandu was significantly altered by coexistence with the weeds; height of the control forage at 120 days was 62 cm compared with a mean of 38 cm for the remaining treatments (Table 1).
Table 1 Effects of duration of weed competition in a Marandu pasture on height, number of tillers per plant and density of Marandu plants at 120 days after emergence.
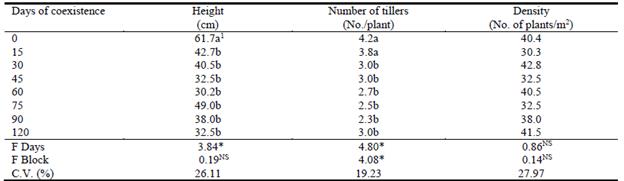
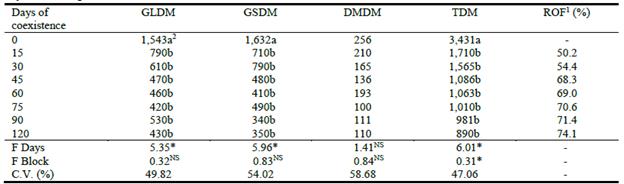
Similarly, exposure to weeds for periods of >15 days significantly reduced the number of tillers per plant (4.2 and 3.8 for 0 and 15 days, respectively, vs. a mean of 2.7 for the remaining treatments; P<0.05) (Table 1). Density of Marandu plants did not differ between treatments (P>0.05) with a mean of 37.3 plants/m2 at 120 days post emergence (Table 1). The production of green leaf, stem and total forage dry matter was significantly reduced by the presence of weeds in the sward, even for as little as 15 days (P<0.05; Table 2). After 120 days of growth, total DM production was suppressed by 50%, when weeds remained in the pasture for as little as 15 days after emergence. This reduction in yield had increased to 74% when weeds remained in the pasture for the full 120 days (Table 2), but there were no significant differences between yields for the differing times that weeds remained in the pasture.
Table 2 Effects of duration of weed competition on the production (kg/ha) of green leaf dry matter (GLDM), green stem dry matter (GSDM), dead material dry matter (DMDM) and total dry matter (TDM) of Marandu and reduction in forage offering (ROF) at 120 days after emergence.
The relationship between total DM yield and number of days that weeds remained in the pasture is presented in Figure 4. Accordingly the suppressant effects of weeds on pasture growth had been expressed by 45 days after emergence.
Discussion
This study has shown that weeds can provide severe competition for freshly sown stands of Marandu, which reduces DM production of the grass, even if weeds are present only for a short period initially, e.g. as little as 15 days. It could be concluded that this is due to competition for available resources, especially nutrients, sunlight and moisture.
Interference of the weeds was most evident on the height and number of tillers generated by Marandu plants. Nepomuceno et al. (2007) reported that plant height is not an adequate characteristic to evaluate the competition between species, since plants subjected to competition prioritize height growth in search of light rather than the accumulation of total DM. Vilela (2011) commented that tillering is a predominant feature in most grasses, and in the case of a pasture, the success of its production is related to good tillering and the consequent occupation of spaces between plants, thereby complicating the establishment of weed species. In this study the opposite was observed, i.e. the weeds dominated the spaces and adversely affected the emergence of tillers in the forage, even when present for only 15 days. In addition grass plants subjected to competition from weed plants for only 15 days showed lower vertical growth than those free of weed competition and failed to compensate when weeds were removed.
Both S. obtusifolia and H. suaveolens are annual weeds and grow rapidly early in life, growing much more rapidly than Marandu and the perennial S. rhombifolia. This may explain why the main suppressant effect of weeds on Marandu growth happened in the first 15 days following emergence. These weeds are native to the American continent with wide distribution in South America, especially in Brazil, and often infest areas of annual crops, perennials and pastures (Fleck et al. 2003; Souza et al. 2011). Besides the competition for environmental resources such as water, light and nutrients, they can also be toxic to animals if ingested during grazing (Pellegrini et al. 2007; Braga et al. 2012).
We found that S. obtusifolia grew most rapidly between 21 and 91 DAE, in agreement with the findings of Erasmo et al. (1997), after which growth rate decreased significantly. Those authors suggested that the reduction in growth was due to the natural senescence process of the species, with resources at that time being directed to reproduction. Studies performed by Gravena et al. (2002) demonstrated that H. suaveolens accumulated only 31% of total DM up to 45 DAE, with most rapid growth between 60 and 140 DAE. Our findings support these results with most rapid growth of this species between 75 and 120 DAE. It is significant that DM yields of this species at 120 days were equal to those of Marandu in the unweeded plots. Sida rhombifolia, despite the low DM accumulation during virtually the entire period, began to show greater vegetative growth only after 90 DAE, which is in agreement with findings of Bianco et al. (2014). When analyzing growth and mineral nutrition of S. rhombifolia, these authors found that the species actually has slow vegetative growth up to 80 DAE, with peak DM accumulation occurring after 140 DAE. However, the authors also found that, although initial growth is slow, this species has a high capacity for absorbing nutrients from about 35 DAE and has a comparatively higher capacity to absorb nutrients than other weeds. It is important to note that S. obtusifolia and H. suaveolens are annuals and S. rhombifolia is a perennial species. Annual species grow more quickly than perennials and this was probably the main factor suppressing growth of Marandu.
The different patterns of DM accumulation of the 3 weed species help to explain the similarity of the relative importance obtained for the various coexistence periods used in this study. This behavior may be correlated with competitiveness and the consequent coexistence of the 3 weed species in the area. In other words, the weed species complement one another, with S. obtusifolia having greater competitive ability in the early growth stages, while H. suaveolens and S. rhombifolia are able to grow slowly during this period, being more prominent as Senna matures (Gravena et al. 2002).
With this complementary growth, there was always at least one weed species exerting competitive pressure on Marandu, resulting in similar suppression of grass growth regardless of when weeds were removed. It must be remembered that the annual weeds are more adapted to the environmental conditions, have characteristics that make better use of the light, and are more efficient in the use of nutrients, since they are native in the region, while the majority of forage grasses are exotic in South America (Peron and Evangelista 2004; Benett et al. 2008). Most of them are also perennials with slow growth initially.
It is noteworthy that the main broadleaf weed control method in grazing areas is the application of specific selective herbicides. This represents a significant operating expense. Our study provides an indication of the possible reduction in pasture growth as a result of weed infestation, which may aid a farmer in making a decision to spray weeds or to let them grow. It seems that, if the weeds are not treated early after emergence, it is not worth treating them in terms of the effects on pasture growth in that season, as they will still have a depressant effect on pasture growth, even if removed. There is a small window of opportunity to take action. The decision then becomes one of preventing seed set to reduce the soil seed bank of the weeds for subsequent seasons. Our results show that the reduced growth of Marandu from weed competition was a function not of fewer grass plants but reduced growth of the same number of plants, through reduced numbers of tillers and reduced vertical growth. We are not aware of similar findings being reported in the scientific literature.
We consider that the regression equation obtained in this study should be of assistance to all professionals involved in the livestock production chain, especially the meat production chain, in which the productivity of a forage area is heavily dependent on the presence or absence of weeds. It should be used as a component when making decisions about weed control measures to ensure that the decision is soundly based economically. However, delaying a decision to treat weeds could have long-term consequences, especially in terms of additional seed added to the soil seed bank. As this study lasted for only 120 days, it is difficult to forecast the productivity of the pasture over subsequent years. Longer-term studies are needed to determine how the suppression of growth of the grass observed here is reflected over the complete life of the pasture.

