











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Feeding the world population is a major challenge for the future when one considers that in 2050 there will be an expected >9 billion people on this planet. Consequently, food production must be increased and intensified (FAO 2010). At the same time, there is growing concern about the environmental impact of agricultural production, in particular of livestock (Steinfeld et al. 2006). While past agricultural research focused primarily on increased production, it is now well recognized that ecological concerns must be addressed as well if environment-friendly production strategies are to be developed and sustainable intensification (SI) is to be achieved (Garnett et al. 2013; The Montpellier Panel 2013). SI encompasses increased production from existing farmland without negatively affecting the environment, and the approach has been adopted as a policy goal for a number of national and international organizations working towards sustainable development goals. This SI policy goal applies also to research on tropical pastures and forages and is reflected, for example, in the theme of the last International Grassland Congress (New Delhi, India, November 2015): Sustainable Use of Grassland Resources for Forage Production, Biodiversity and Environmental Protection.
Two recent overview analyses of tropical forage-based livestock production systems vis-à-vis the environment and the need for SI concluded that such systems can have a positive impact on the environment (Peters et al. 2013; Rao et al. 2015). In tropical production systems, the term 'forages' refers mostly to grasses, since adoption of legume technology in the past has been rather low (Shelton et al. 2005). We hypothesize, however, that tropical forage legumes do have the potential to play a particular, positive role in addressing environmental concerns.
Therefore, complementing the above-mentioned overviews and in order to contribute to the development of research strategies, in this paper we analyze the effects of tropical forage legumes (pasture plants for grazing or fodder plants for cut-and-carry or browsing) on the environment. For this, we briefly: outline the main anthropogenic environmental issues; highlight some essentials related to livestock production and the environment; and discuss the key attributes of forage legumes that contribute to natural resource conservation and environmental protection with a particular emphasis on adaptation to and mitigation of climate change. We then examine the potential of tropical forage legumes to have a positive impact on environmental issues and provide ecosystem services.
Environmental issues
The main, human-induced environmental problems, as currently perceived, are related to: natural resources, including biodiversity; and climate change.
Regarding natural resources, it is generally accepted that the major issues are: (1) ecosystem destruction and degradation; (2) soil degradation and loss; (3) water degradation and loss; and (4) biodiversity degradation and loss. Obviously, these problem areas are all interrelated.
Regarding climate change and its major manifestations (global warming leading to modifications of rainfall regimes and both flooding and drought phenomena), IPCC (2014) states that the main driver is increased anthropogenic greenhouse gas (GHG) emissions, mainly carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O).
Livestock production and the environment: Some background essentials
When considering livestock production in the tropics and its impact on the environment, a few issues should be highlighted:
In the scientific and non-scientific communities, livestock production, including grazing, is blamed for severe negative impacts on the environment (e.g. Steinfeld et al. 2006; Hyner 2015). Livestock production is estimated to contribute 14.5% of all anthropogenic GHG emissions globally (Gerber et al. 2013).
The demand for animal products, especially foods derived from livestock, is expected to increase considerably in the future, particularly in South, East and Southeast Asia, and to a lesser extent in Sub-Saharan Africa, as a consequence of increasing living standards (Rosegrant et al. 2009; Robinson and Pozzi 2011).
In view of physical limitations to expansion of land area for agricultural production (both crop and livestock), future production increases must come mainly from intensification of production systems (The Montpellier Panel 2013).
Ruminant livestock (e.g. cattle, buffalo, sheep, goats) play an important role as they convert vegetation, which is unsuitable as food for humans, into high-quality products for human consumption. Nonetheless ruminants are fed grain-based diets (such as in feedlots), and this practice is in direct competition with humans for that food source (Mottet et al. 2017).
Tropical grazing lands often occupy marginal land that is unsuitable or only marginally suitable for crop production, because of constraints imposed by soil physical and chemical properties, topography (including slopes and waterlogging) and climatic conditions (Rao et al. 2015). Similarly, some lands are temporarily unsuitable for crop production due to drought or excess of water, and these areas are expected to increase in the tropics (Zabel et al. 2014). Here, crop production could benefit from a crop-forage rotation. • As far as research on tropical pastures and forages is concerned, the literature provides almost no indication that, in the past, environmental issues have played a major role in forage development and utilization. Notable exceptions are the concerns expressed by McIvor et al. (1997; 2005) and Noble et al. (2000).
Key attributes of legumes
The main 5 features of this plant family in general are summarized as follows:
Legumes in the Papilionoideae subfamily and in what used to be the Mimosoideae subfamily [now the ' mimosoid clade' in the newly defined Caesalpinioideae subfamily (LPWG 2017)1 and a few taxa in the Caesalpinioideae subfamily can fix, in symbiosis with rhizobia (Bradyrhizobium, Rhizobium), atmospheric nitrogen (N). Therefore they have the potential to: (1) be N self-sufficient; and (2) increase N availability in the soil for associated or subsequent crops, forage grasses and soil biota. Depending on legume species, effectiveness of rhizobium strains, nutrient supply (mainly phosphorus, potassium and the trace element molybdenum), climatic conditions and assessment method applied, published data for symbiotic N fixation (SNF) by tropical forage legumes cover a wide range, e.g. 15-158 kg N/ha/yr using 15N methodologies (Thomas 1995); a recent example is the range of 123-280 kg symbiotically fixed N/ha/yr in 6 Arachis glabrata cultivars, reported by Dubeux et al. (2017a). Total input of SNF to mixed grass-legume pasture systems can range from 98 to 135 kg N/ha/yr (Boddey et al. 2015). This attribute is particularly important in production systems that depend on external N inputs (Douxchamps et al. 2014).
Most forage legumes have high nutritive value for ruminants, mainly in terms of concentration of crude protein (CP) (percentage N x 6.25) but also of energy (Lüscher et al. 2014). This feature can be particularly significant in mixtures with, or as complement to, grasses with CP levels often below livestock maintenance requirements or when low-CP and low-digestibility crop residues are fed.
Many legumes have a deep-reaching taproot system, providing access to water and nutrients in deeper soil layers (Rao 1998; Dubeux et al. 2015), which contributes to improved cycling of both N and P (Thomas 1995; Oberson et al. 2006).
There is an enormous organismal (taxonomic) and genetic diversity in the Leguminosae (or Fabaceae) family with almost 20,000 species (Williams 1983; Lewis et al. 2005) in formerly 3, now 6 (LPWG 2017), subfamilies. This includes annuals and peren-nials, growth forms ranging from herbaceous, prostrate species (e.g. Arachis pintoi) to vines (e.g. Centrosema spp.), subshrubs (many Stylosanthes spp.), shrubs (e.g. Cratylia argentea) and trees (e.g. Leucaena spp.). Such diversity suggests that a very wide range of production-relevant features, in terms of adaptations to abiotic and biotic constraints, biomass production potential etc., could be expected; they warrant further exploration.
A wide range of phytochemicals (secondary metabolites) occur in many species of the Leguminosae. These are often referred to as ' antinutritional factors' when legume feeding to livestock is considered (Kumar and DMello 1995).
These key features imply that legumes can have a significant ecological advantage over other plant families. However, it is also via this ecological advantage that a legume can become a weed that threatens biodiversity and/or agricultural productivity and can also affect productivity via soil acidification (see below).
Tropical forage legumes and natural resources
Concern 1. Ecosystem destruction and degradation
This concern encompasses both the destruction of natural ecosystems such as forests and the degradation of areas that have already undergone land use changes, such as unproductive, mismanaged pastures. 'Prevention is better than cure' - so the initial approach to this problem should be taking measures to avoid ecosystem destruction and land degradation in the first place. Solving this issue does not require development of technology but rather application of existing appropriate land use policies and strategies.
Among them is the SI policy goal of concentrating production on existing agricultural land (Garnett et al. 2013; The Montpellier Panel 2013), thereby lowering the colonization pressure on natural ecosystems that should be considered as ecological and biodiversity reserves. Intensification, however, is usually closely linked to N fertilization and its detrimental consequences for the environment (nitrate leaching and emission of N2O, a potent GHG; see below).
Forage legumes can contribute to SI by providing N to the soil-plant system and high quality forage to livestock. By this, the productivity of land and livestock can be substantially increased in production systems with grass-legume pastures and/or legume-only protein banks. In Table 1 a number of examples in the tropics are presented.
Table 1 Effects of tropical forage legumes on liveweight gain of cattle (extracted from Rao et al. 2015).
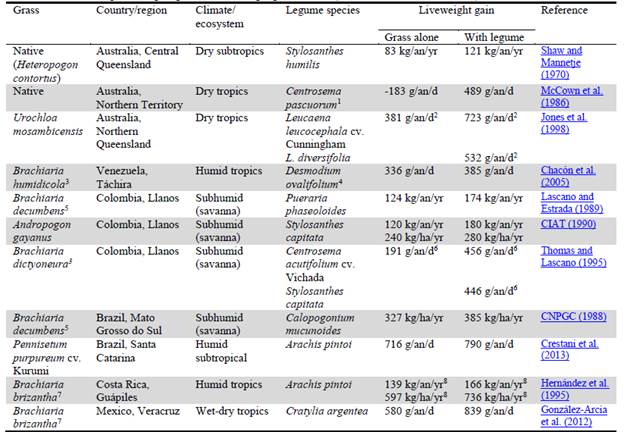
Supplementation as ley during the main dry season; 2192 grazing days; 3Now classified as Urochloa humidicola; 4Now classified as Desmodium heterocarpon subsp. ovalifolium; 5Now classified as Urochloa decumbens; 6Means of 3 grazing cycles totalling 385 days, newly established pastures; 7Now classified as Urochloa brizantha; 8Mean of 2 stocking rates (low and high).
There is also significant potential to increase overall land productivity via mixed-production systems such as agropastoral systems (Ayarza et al. 2007; Boddey et al. 2015), including intercropping forage legumes (Hassen et al. 2017), and (agro) silvopastoral systems (Nair et al. 2008; Dubeux et al. 2015). Multi-purpose legumes serve multiple roles, e.g. Leucaena leucocephala provides wood and forage, while Desmodium heterocarpon subsp. ovalifolium (' D. ovalifolium' ) and Arachis pintoi can control erosion, suppress weed growth and provide forage. Dubeux et al. (2017b) reviewed the role of tree legumes and their benefits in warm-climate silvopastoral systems and concluded that they were a key component for the SI of livestock systems in that climatic zone. Research has shown that, once mismanaged land has become unproductive, both herbaceous (Ramesh et al. 2005) and woody legumes (Chaer et al. 2011) can be used successfully for rehabilitation of degraded land, including degraded cattle ranching land (Murgueitio et al. 2011).
Concern 2. Soil degradation and loss
Soil degradation and loss are intimately linked to the previous concern, ecosystem destruction and degradation. The loss of top soil, where most soil organic carbon (SOC) and plant nutrients are concentrated (Lal 2010), leads not only to loss of a stratum that is crucial for plant production but also to oxidation of SOC and subsequent liberation of the GHG, CO2. Since this carbon stems from recent (= not fossil) photosynthesis, it does not alter the longer-term CO2 balance in the atmosphere. However, it is lost from a key carbon sink: soil organic matter (SOM).
Among the multiple possibilities (most of which are based on legume N contribution, soil-covering growth habit and deep root system) to contribute to the mitigation of this environmental problem, are:
Soil conservation by: cover legumes such as Alysicarpus vaginalis, Arachis pintoi and Desmodium 'ovalifolium ' which prevent erosion; contour-hedges with shrub species such as D. cinereum and Flemingia macrophylla; and leguminous trees such as Erythrina spp. and Leucaena spp.
Rehabilitation of degraded soils by pioneering legumes such as Stylosanthes spp., Macrotyloma axillare and Flemingia spp., which are deep-rooted and adapted to infertile soils, with soil improvement resulting from cycling of minerals from deeper soil layers and enhanced concentration of SOM through litter production (Amézquita et al. 2004); ( Boddey et al. 2015). In the case of tannin-rich species, such as F. macrophylla, litter has a marked impact as it decomposes slowly (Budelman 1988) and provides a longer-lasting soil cover and slow nutrient release.
Exploring and exploiting the potential of legumes to ameliorate compacted soil, as shown by e.g. Rochester et al. (2001) for Lablabpurpureus (among other, more temperate grain legumes) and Lesturguez et al. (2004) for Stylosanthes hamata.
Exploring and exploiting the potential adaptation of species to soil salinity. There seems to be some potential in a few genera such as Acaciella, Desmanthus, Neptunia and Sesbania(Cook et al. 2005).
Concern 3. Water degradation and loss
On a global scale, water and its decreasing availability, accessibility and quality, are major concerns (Rogers et al. 2006). As far as tropical pastures and forages are concerned, we see the role of legumes primarily in the following areas:
Use of drought-adapted species, e.g. deep-rooted herbs and subshrubs such as Centrosema brasilianum and Stylosanthes guianensis; shrubs and trees such as Cratylia argentea and Leucaena leucocephala(Cook et al. 2005) ; or species with physiological mechanisms for avoiding and/or tolerating water stress (annual life cycle, narrow leaflets, leaf movements, tolerance of very low leaf water potentials), such as Centrosema pascuorum(Ludlow et al. 1983; Clements 1990).
Reducing sedimentation of water bodies. Sedimentation is a major issue with devastating consequences in times of excessive rainfall and is, obviously, intimately linked to soil erosion by water. Consequently, the potential role of legumes consists primarily in prevention of soil erosion (see above). Additional potential lies in watershed protection through productive, N self-sufficient multipurpose trees.
Enhancement of water infiltration via the potential amelioration effect on soil structure of legumes (see above).
Using cover legumes to control weed growth in oil palm and rubber plantations as an attractive alternative to the use of herbicides.
Replacing N fertilizer, at least partly, by a legume. This could reduce nitrate leaching and water eutroph-ication as both groundwater contamination by nitrate leaching and N-eutrophication of water bodies as a consequence of surface runoff are recognized negative consequences of N fertilization in tropical pastures (Vendramini et al. 2007).
Concern 4. Biodiversity degradation and loss
Any land use change, such as the establishment of forage species, has profound implications for biological diversity (Alkemade et al. 2013) in terms of plant and animal species and ecotypes, including entomofauna and the whole soil biota in the area concerned. This is particularly true if a monospecific grass sward is established, as is common in the tropics. While this is an area of considerable knowledge gaps, we claim that the inclusion of an N-fixing and, subsequently, SOM-increasing legume in a mixture with a grass will mitigate the overall negative effects of such a land-use change on biodiversity, namely entomofauna and soil biota (Ayarza et al. 2007). In their review which focused on temperate conditions, Phelan et al. (2015) reported on positive effects of legumes on the diversity and abundance of pollinating insects and earthworms.
In this context, the possible mitigating effects on biodiversity loss of using mixtures of legume species should be explored. Mixtures of herbaceous cover legumes are commonly used for weed control in Southeast Asian tree plantations, e.g. Calopogonium mucunoides, C. caeruleum, Centrosema pubescens (now classified as C. molle), Desmodium ovalifolium (now classified as Desmodium heterocarpon subsp. ovalifolium) and Pueraria phaseoloides(Jalani et al. 1998). Such mixtures might also improve functional biodiversity.
A related area is the role that forage legumes can play in combating agricultural pests through exudation of chemical compounds. A significant example is the increasing use of Desmodium intortum and D. uncinatum as intercrops to control maize stemborer and Striga spp. in the so-called push-pull systems in East Africa (Khan et al. 2010; icipe 2015).
Negative aspects of tropical forage legumes
Two negative aspects of tropical forage legumes must be recognized:
Weed potential. The danger that an exotic legume could become a serious invasive weed that threatens local biodiversity and/or affects crop production must be considered. According to available literature, this risk seems to be a particular concern in Australia, even to the point that Low (1997) suggested that introduction of exotic forage germplasm should cease with the focus changing to developing cultivars from native species. Among the factors contributing to the weed potential are (Driscoll et al. 2014): region- or production system-specific lack of grazing or browsing animals; unpalatability or low palatability to livestock, due to presence of secondary metabolites; prolific seeding; and presence of thorns and spines. Tropical legume species currently listed among the 32 land plant species of "100 of the world's worst invasive alien species" (Lowe et al. 2004) include: Acacia mearnsii, Leucaena leucocephala, Mimosa pigra, Prosopis glandulosa and Pueraria montana var. lobata. It is well recognized that attributes which make a legume a useful pasture species are the same as those which allow it to become potentially a serious weed.
Even if a legume might not represent a risk to biodiversity on a larger scale, at the pasture level soil N accumulation following eventual legume dominance could lead to changes in species composition: nitro-philous weeds can become an agroecological problem (McIvor et al. 1996).
Soil acidification. Continuous use of legume-only or legume-dominated swards can result in soil acidification as Noble et al. (1997) and Liu et al. (1999) reported for Stylosanthes species in Australia and China, respectively.
It has been suggested that increased presence of a grass reduces the problem (Scott et al. 2000).
Tropical forage legumes and climate change
Increase in GHG emissions is claimed to be the main causal agent of climate change (Adger and Brown 1994). In low-income countries, that is, in the developing world, agriculture and land use changes are estimated to contribute 20 and 50%, respectively, to overall GHG emissions (The World Bank 2010). Climate change is expected to: (1) raise temperatures across the planet; and (2) disturb rainfall patterns, but regional differences will occur, resulting in increases of both drought-stricken and waterlogged areas, and salinization of agricultural soils (IPCC 2014; Zabel et al. 2014; Brown et al. 2015 ).
General strategies to cope with climate change are: adaptation to the modified climatic conditions; and mitigating GHG emissions that lead to climate change. Both are examined in relation to tropical forage legumes as follows:
Adaptation potential
We suggest that research make use of the large organismal (= taxonomic) and genetic diversity of tropical forage legumes that is available in the world's major germplasm collections, e.g. particularly those held by the Australian Pastures Genebank, CIAT (Centro Internacional de Agricultura Tropical), Embrapa (Empresa Brasileira de Pesquisa Agropecuária) and ILRI (International Livestock Research Institute). Collections can be screened for adaptation to constraints such as high temperatures and tolerance of drought, waterlogging or soil salinity (Baron and Bélanger 2007). As a result of phenotypic evaluation within the naturally available diversity, promising germplasm can be developed further via selection or breeding (Araújo et al. 2015).
In this context, existing legume germplasm collections need to be complemented by further gathering of wild germplasm in the field. Collecting missions should focus on areas which experience drought or waterlogging or soil salinity problems, i.e. areas where naturally occurring plants can be expected to have the desired adaptations for survival and productivity.
Mitigation potential
While a recent overview (Peters et al. 2013) concluded that tropical pastures and forages in general have the potential to play a significant role in mitigation of climate change, the following discussion refers specifically to the contribution of forage legumes.
Carbon dioxide (CO2). The work of Fisher et al. (1994) in the Colombian Llanos showed that sown, deep-rooted tropical grasses can accumulate more SOC than native savanna, in fact, almost as much as under forest. When a legume was mixed with the grass, the amount of C stored in the soil (0-80 cm) increased by 20% to a total of 268 t C/ha. Tarré et al. (2001) reported that, in the humid tropics of Bahia, Brazil, soil C accumulation (0-100 cm soil depth) in a Brachiaria humidicola (now accepted as Urochloa humidicola)-Desmodium ovalifolium (now accepted as Desmodium heterocarpon subsp. ovalifolium) pasture over a 9-yr period was almost twice that of a B. humidicola pasture (1.17 vs. 0.66 t C/ha/yr). Contributions by non-tropical permanent pastures and perennial legumes to increased C accumulation in the soil are cited in the review of Jensen et al. (2012). According to these authors, the organic N provided by the legumes fosters C accumulation. As Smith et al. (2008) and Chaer et al. (2011) showed, trees in agroforestry systems, particularly leguminous trees, have the potential to increase C accumulation in the soil considerably, as well as accumulating C in their own biomass, especially on degraded land.
On the other hand, respiration by legume roots during the energy-consuming SNF process releases substantial amounts of CO2 to the atmosphere, even more CO2 per unit N than is emitted during the production of industrial N fertilizer (Jensen et al. 2012). As these authors point out, however, in contrast to CO2 from fertilizer production, CO2 produced during SNF stems from photosynthesis, so the atmospheric CO2-concentration balance is not altered.
The particular role of SOM merits further emphasis. This is the most important carbon sink and can be larger than the above-ground C in a tropical rainforest (Lal 2010). If soil erodes, this eventually leads to oxidation of C to CO2, which is released to the atmosphere (Olson et al. 2016). Therefore, perennial plants, e.g. grasses and legumes, which provide soil cover and prevent erosion, play a particularly significant role in mitigating CO2 emissions in tropical production systems. To guarantee this environmental benefit, vegetation/pasture management must be such that there is always adequate soil cover. Creeping, stoloniferous species such as Desmodium ' ovalifolium' and Arachis pintoi that provide a dense soil cover - while supplying N-rich litter - appear to be of particular interest. It must, however, be mentioned that, because of the low C: N ratio of legumes, SOM under legume-only vegetation is less stable than under a grass-legume mixture (Sant-Anna et al. 2017).
Methane (CH4). Methane has 25 times greater global warming potential per unit mass (100-yr time horizon) than CO2. In agriculture, it is generated mainly by enteric fermentation, manure management and rice cultivation. By nature ruminants produce enteric CH4 (Broucek 2014) and research is underway to determine how this might be modified. Options are either to increase the amount of meat or milk produced per unit of CH4 emitted or to decrease the amount of CH4 emitted per unit of feed intake through: (1) providing high quality forage, mainly in terms of CP concentration and digestibility; and (2) improving livestock breeds that are able to respond to improved forage quality with increased productivity (Gerber et al. 2013).
In a recent meta-analysis, Lee et al. (2017) showed that rising temperatures lead to decreased nutritive value of grasses and increased CH4 emissions by ruminant livestock, which worsens the global warming scenario. On the other hand, forage legumes have high nutritive value and can contribute to lower emissions of CH4 per unit of livestock product or unit of feed ingested. A study by Molina et al. (2016) of methane emissions of Lucerna heifers fed a Leucaena leucocephala-stargrass mixture or grass only demonstrated the benefits of the legume in the diet in reducing methane emissions per unit gain. The optimal situation is to have improved livestock feeding, based on high quality forage including legumes, combined with improved livestock breeds that can more efficiently use such improved feed.
In addition to this general quality-based role of forage legumes regarding enteric CH4, another meta-analysis (Jayanegara et al. 2012) showed that polyphenols such as condensed tannins, i.e. secondary metabolites that occur in many tropical forage legumes, decrease CH4 emissions. According to an analysis based on 22 in vivo studies, ruminants fed warm-climate legumes produced less CH4 per kg OM intake than ruminants fed cold-climate legumes, C3 grasses and C4 grasses (Archimède et al. 2011). Low-molecular weight tannins, such as those in L. leucocephala(Molina et al. 2016), can also play a role. It is important to ensure that tannins in the diet do not reduce protein digestibility, compromising animal intake and thus its performance, which in turn will affect CH4 emissions per unit of livestock product. Working with subterranean clover (Trifolium subterraneum) Kaur et al. (2017) showed that a plant breeding approach to reduce methanogenesis has potential.
Nitrous oxide (N2O). Nitrous oxide has 300 times greater global warming potential per unit mass (100-yr time horizon) than CO2. Its production by soil microorganisms during nitrification and denitrification processes is very much related to the use of N fertilizers in agriculture (Subbarao et al. 2013). In their meta-analysis, Jensen et al. (2012) concluded that there is a tendency for lower N2O production from soil under legumes than from systems based on industrial N fertilizer, depending on the amount of N fertilizer applied. This seems to be an area of considerable knowledge gaps in relation to tropical forage legumes.
In view of the recent detection of biological nitrification inhibition (BNI) in some tropical forage grasses, particularly Brachiaria (now Urochloa) humidicola(Subbarao et al. 2009; 2017), the challenge is to determine whether such a mechanism might also exist in tropical forage legumes. It might then be possible to exploit the synergy between SNF and BNI to the benefit of both agriculture and the environment. Due to BNI, symbiotically fixed N might be available for longer periods and less prone to loss by nitrate leaching and N2O production.
Discussion and Conclusions
Ecosystem services
In the preceding sections, we showed that tropical forage legumes have considerable potential to increase productivity of forage-based livestock systems, while providing benefits to the environment. The environmental benefits, subsumed under 'ecosystem services', comprise positive effects on: soil conservation and soil chemical, physical and biological properties; water balance; mitigation of global warming and of groundwater contamination; saving of fossil energy; functional biodiversity (soil, entomofauna); and rehabilitation of degraded land. The combination of these features makes tropical forage legumes particularly valuable at all levels of the system because of their interaction with plants, soil, animals and the atmosphere. This environmental role could be considered as a 'new' important dimension of tropical forage legumes.
A crucial aspect, however, is: During past decades the beneficial role of tropical forage legumes was promoted with the sole focus on livestock production and soil fertility; what must be done to have legume-based technologies more readily adopted by farmers now that general environmental benefits are recognized?
Legume technology adoption and payment for ecosystem services
In their review paper, which examined the role of forage legumes in general (though they focused primarily on temperate zones), Phelan et al. (2015) reported a low and even declining use of forage legumes. We must recognize that in the tropics adoption of legume-based technologies has, in general, been disappointing - in spite of many success stories with tropical forage legumes worldwide (see the 33 contributions in Tropical Grasslands Vol. 39, No. 4, 2005; goo.gl/Qf5VJu). The reasons were analyzed by Shelton et al. (2005) and include a number of issues that should be taken into account when planning R&D programs promoting the use of tropical forage legumes. A particularly important issue is the organization of efficient seed production systems. The lack of seed availability is often cited as a key reason for adoption failure and the resulting vicious circle (lack of robust demand - lack of interest of the private seed production sector - lack of seed production and availability - lack of adoption) needs to be broken. Successful results have been achieved with contracting farmers for forage legume seed production and farmer to farmer seed sales, e.g. in Thailand, India and Bolivia. For large-scale adoption it will be essential to develop systems which ensure high seed quality and are commercially viable (Shelton et al. 2005).
We doubt that an eventual recognition of the 'new' ecosystem services role of legumes will modify farmers' lack of enthusiasm for legumes to a marked extent. Although promotional and educational activities, along with results from further research involving farmer participation, might be helpful, we expect that constraints imposed by the need for management skills and investments will remain, unless attractive economic incentives are offered to farmers (White et al. 2013). Such incentives should not be restricted to legume-based technologies but should extend to all tropical forage technologies which provide environmental services. We suggest that schemes of payment for ecosystem services (PES) (Pagiola et al. 2004; Van Noordwijk and Leimona 2010), applicable to both smallholders and large livestock producers, be explored, developed and implemented.
The need for life cycle assessments
Inputs of N are necessary in all pastures if livestock productivity is to be increased, such as within the concept of SI. Basically, there are 2 options: (1) planting legumes with SNF capability in mixtures with grasses; and (2) applying industrial N fertilizers to grass-only swards.
Greenhouse gas emissions from both approaches should be measured. We suggest that full life cycle assessments for tropical pastures addressing the whole carbon footprint (Eshel et al. 2014) should be performed. In their temperate climate-focused review, Phelan et al. (2015) reported that CO2-equivalent emissions for Trifolium repens-grass pastures were 11-23% lower than for N-fertilized grass. Such life cycle assessments must include the need for fossil energy and any benefits to any subsequent crop in a rotational system (de Vries and de Boer 2010; Jensen et al. 2012).
Research needs
The suboptimal adoption of forage legume technologies in the past - when only forage dry matter and/or livestock production was considered - has led to a substantial decrease in research on tropical forage legumes during the last 2 decades. We argue that, in view of current environmental concerns, this research should be resumed with adequate funding support at national and international levels.
We have shown that a substantial body of evidence suggests that forage legumes have potential to contribute significantly to environment-friendly agricultural land use and sustainably intensified livestock production in the tropics. However, there is still a lack of hard data, and several statements in our analysis are not yet well substantiated and need to be verified and confirmed. Further research is required to provide decision makers with a solid database on the ecosystem services from utilization of tropical forage legumes. Priorities in different regions will depend on differences in climate, soil types, land use, production systems etc. Preferably, such research should be conducted within a coordinated network or consortium, e.g. similar to those European initiatives with focus on temperate legumes (Lüscher et al. 2014).
We have compiled the following list of 'research for development' themes on tropical forage legumes as a result of our analysis:
life cycle assessments to compare the carbon footprints of livestock feeding based on forage legumes with that based on N fertilizer in different production systems;
potential of legumes for enhancing functional biodiversity, including in multi-species mixtures;
further understanding of the potential of forage legumes in (1) crop-livestock systems, (2) soil stabilization and (3) reversing land degradation;
further understanding of the impact of legumes on associated vegetation (species composition);
assessment of the impact of promising legume species on rumen methanogenesis;
identification of tanniniferous legumes which concurrently provide high quality forage in terms of digestibility in the rumen and reduced methane emission intensity;
identification of anti-methanogenic compounds other than tannins in legume forage;
assessment of the BNI potential of forage legumes;
development of methodologies for payment for ecosystem services;
optimization of SNF via enhanced exploration and exploitation of rhizobia diversity; and
targeted collection of wild legume germplasm for development of varieties with improved adaptation to climate variability and change.

