Carlos J Tabares 1, MV; Ariel M Tarazona1, Zoot; Martha Olivera Ángel 1, MV, Dra. Sci. Agr.
1 Reproducción, Fisiología y Biotecnología, Corporación Biogénesis, Universidad de Antioquia, AA 1226, Medellín, Colombia.
jatabares@agronica.udea.edu.co
(Recibido: 3 marzo, 2004; aceptado: 8 junio, 2005)
El principal objetivo de esta revisión fue el estudio de los mecanismos que regulan la movilidad espermática en peces de agua dulce, dada su importancia en el desarrollo de biotecnologías para la conservación de sus gametos, los cuales una vez producidos en el testículo, se capacitan en el conducto espermático donde permanecen inmóviles por factores como el balance iónico, la osmolaridad, el pH o algunas proteínas. En el momento de la liberación de los espermatozoides al medio acuoso, estos son activados por un periodo de tiempo muy corto. En teleósteos de agua dulce, el choque hiposmótico induce cambios de potencial de membrana que conducen a la regulación de canales iónicos de K+ y de Ca2+, seguida por un flujo iónico que desencadena la cascada de activación. Muchas investigaciones se han enfocado en el estudio de la osmolaridad, el balance iónico, el pH y factores dependientes de AMPc y Ca2+; asociados con la activación de la movilidad espermática. De esta forma, los mecanismos fisiológicos de la activación han ganado interés para comprender los procesos por los cuales el espermatozoide adquiere la hipermovilidad para alcanzar el oocito y penetrar el micrópilo antes del cierre del mismo.
Palabras clave: balance iónico, choque hiposmótico, movilidad, osmolaridad, potencial de membrana.
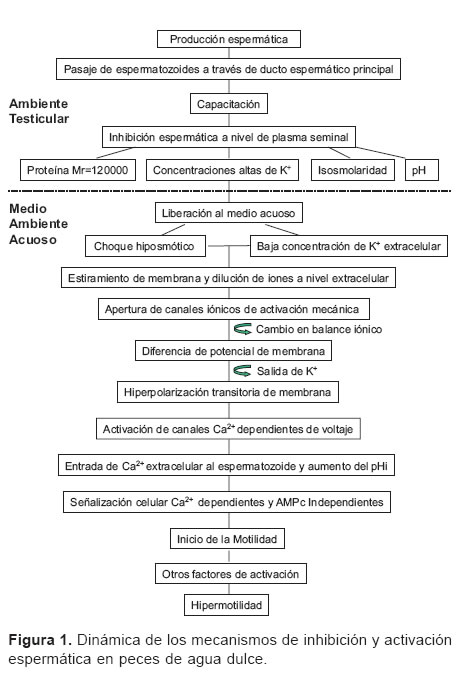
]]> IntroducciónLos espermatozoides de peces cuando son liberados y cierran su micrópilo, por lo cual los espermatozoides al agua cuentan con un breve periodo de activación liberados durante la espermiación deben activarse y (limitado a pocos segundos en varias especies) moverse en el agua para lograr la fertilización antes expresada en movilidad y velocidad de desplazamiento de este periodo. Es posible que estos mecanismos se progresivo para lograr la fertilización. Al contrario de hayan dado evolutivamente debido al corto periodo de lo que sucede en mamíferos, los espermatozoides de competencia para la fertilización (4), de allí la teleósteos no poseen acrosoma, ya que entran a través importancia de comprender los procesos fisiológicos de un orificio de la ova del llamado micrópilo (1,11). de la activación del espermatozoide en los peces (véase Al ser liberadas las ovas en el agua, éstas se hidratan Figura 1).
El conocimiento de los mecanismos que regulan la movilidad espermática en teleósteos, y particularmente, las rutas de señalización intracelular que la activan o inactivan, es importante para desarrollar biotécnologías apropiadas en el mantenimiento y conservación de germoplasma (22, 27). Por lo tanto, se recomienda realizar estudios que permitan avanzar en las técnicas para regular el potencial de movilidad, con el fin de ser usadas en el mejoramiento de la fertilización artificial.
Así mismo, conociendo los gametos desde el punto de vista biológico, fisiológico y metabólico, se pueden desarrollar tecnologías como la crioconservación, fertilización in vitro, conservación de embriones, androgénesis y ginogénesis (2,12) acordes con las estrategias reproductivas de las especies (20). De esta forma la conclusión de algunos estudios sobre biodiversidad proponen colectar y preservar los gametos de las diferentes especies, aun cuando no estén perfeccionadas las tecnologías enfocadas a repoblamiento, investigación o producción comercial de las mismas.
Generalidades del espermatozoide
Gametogénesis
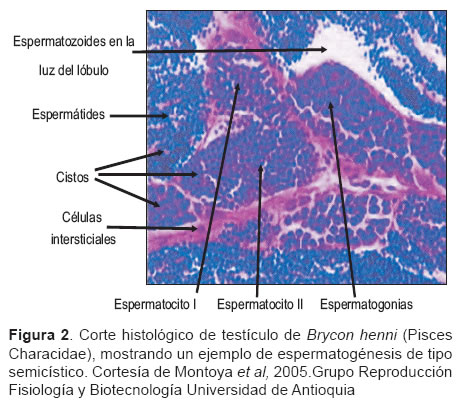
En teleósteos se pueden observar dos tipos de espermatogénesis: una de tipo cístico en la cual el proceso se lleva a cabo completamente dentro de lóbulos, y una de tipo semicístico (1), en la cual el desarrollo ocurre parcialmente fuera del lóbulo, como en moncholo (Hopplias malabaricus) (11).
Dentro de los testículos de Charácidos (espermatogénesis de tipo semicístico) (1) se pueden identificar: espermatogonias, espermatocitos, espermátides y espermatozoides (véase Figura 2). Durante la espermatocitogénesis, la espermatogonia se divide consecutivamente, reduce el diámetro del núcleo y culmina con la formación de células haploides llamadas espermátides, las cuales son liberadas a la luz de los túbulos seminíferos donde se lleva a cabo la espermiogénesis originando los espermatozoides (1).
]]>
El espermatocisto es la unidad espermatogénica de los peces (1) y consiste de un grupo de células germinales en el mismo estadío de desarrollo (11) circundadas por prolongaciones citoplasmáticas de una o más células de Sertoli que forman la pared (1,11) (véase Figura 2).
Las células de Sertoli, además de sus funciones de compartimentalización y secreción, actúan como barrera que impide el contacto entre las células germinales y el sistema vascular; además, fagocitan residuos de espermatozoides. Las células intersticiales o de Leydig, producen esteroides necesarios para la espermatogénesis y expresión de características sexuales secundarias, estas células están presentes en número variable durante el ciclo reproductivo, más abundantes al inicio de la maduración y disminuyen gradualmente su número durante la maduración avanzada y la espermiación (1).
En las espermátides, el citoplasma está simétricamente distribuido alrededor del núcleo, el cual tiene un diámetro de 2-8 micras y cromatina difusa (11). El complejo centriolar en estas células, está ubicado cerca del núcleo y se ancla a la membrana plasmática formando el flagelo. El sistema de endomembranas está bien desarrollado, incluyendo el aparato de Golgi, retículo endoplásmico y algunas vesículas. Las mitocondrias se hallan cerca del segmento inicial del axonema, y hay vesículas distribuidas a través de la pieza media. El flagelo esta rodeado por la membrana flagelar y tiene un aro membranoso. (11)
Al final de la espermiogénesis, el citoplasma residual es eliminado. Los procesos citoplasmáticos de las células de Sertoli se alejan, liberando los espermatozoides dentro del lumen de los túbulos seminíferos (11)
Morfología
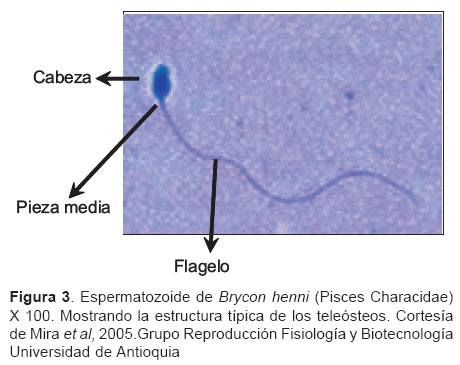
El espermatozoide de los peces con fertilización externa, tiene una estructura simple de tipo primitivo (1) (véase Figura 3), la cabeza mide entre 2-4 micras y es casi esférica con un collar que forma la pieza media donde se encuentran los centríolos y entre 2-9 mitocondrias, el flagelo por lo general está constituido por el axonema en arreglo de nueve pares de microtúbulos periféricos y un par central, sin embargo, algunos grupos taxonómicos como los angiliformes y elopiformes presentan solamente los nueve pares periféricos. El flagelo de algunos peces mide entre 20 y 100 mm y su membrana plasmática forma una especie de aleta en el plano horizontal confiriéndole una forma acintada (4), es posible que esta estructura se haya formado evolutivamente para favorecer el movimiento del espermatozoide bajo ciertas condiciones acuosas. El flagelo de espermatozoides del grupo Otophysi (Cypriniformes, Characiformes, Siluriformes y Gymnotiformes) caracterizado por poseer aparato de Weber, no muestra proyecciones laterales (11).
Los espermatozoides de peces se han clasificado en aquaespermatozoides y en introespermatozoides, según su modo de fertilización externo o interno. Los Cipriniformes, Characiformes y Siluriformes tienen el tipo aquaespermatozoides (11). Lo anterior sugiere que se han desarrollado adaptaciones evolutivas de las diferentes especies para favorecer la fertilización bajo condiciones diferentes, aunque hacen falta investigaciones enfocadas al respecto para esclarecerlo.
Membrana espermática
La membrana plasmática juega un papel fundamental en la respuesta del espermatozoide al entorno, las características propias de la membrana del espermatozoide le confieren la capacidad dinámica para regular diferentes actividades celulares y rutas de señalización que pueden conducir entre otros a la activación de la movilidad espermática. Es por ésto que la integridad de la membrana es un requerimiento absoluto para las funciones que cumple, consecuentemente su pérdida resultará en muerte celular (22).
]]> La composición y organización lateral de la membrana plasmática regula la afinidad por factores de adhesión, controla la permeabilidad de solutos hidrofílicos y dirige eventos de fusión y señalización celular (10). Las propiedades de la membrana están dadas por la proporción lipídica: 70% de fosfolípidos; 25% de lípidos neutros (especialmente colesterol) y 5% glicolípidos (7, 25) y por su organización asimétrica en los diferentes dominios de membrana que le dan una fluidez especial (10).Los estudios reportados sobre la disposición de los fosfolípidos entre las dos capas de la membrana plasmática muestran que la esfingomielina y la fosfatidilcolina son los principales componentes de la capa externa, mientras que los aminoesfingolípidos, fosfatidilserina y fosfatidiletanolamina son los principales componentes en la cara citoplasmática. Esta asimetría transmembranal resulta de la actividad de translocación de los aminofosfolípidos, dependiente de hidrólisis de ATP citoplasmático, los cuales transportan moléculas de fosfatidilserina y fosfatidiletalonamina desde la capa externa a la capa interna (25).
Mecanismos de capacitación, inhibición y activación de la movilidad
Capacitación
Aunque el mecanismo no se conoce completamente, las investigaciones desarrolladas hasta el momento sugieren que los espermatozoides maduran y adquieren capacidad de activación durante el pasaje a través del conducto espermático cuyo fluido (plasma seminal) es de pH básico y rico en bicarbonato (HCO3)-24). El movimiento transmembranal del HCO3 probablemente favorecido por antiportes Na+/H+ sería el responsable del aumento del pH intracelular (pHi) y también podría regular el metabolismo de adenosina monofosfáto cíclico (AMPc), ya que en los espermatozoides la adenilato ciclasa es estimulada directamente por este anión (27, 28).
La adquisición de la capacitación se comprobó al analizar espermatozoides de ayu (Plecoglossus altivelis) provenientes directamente de tejido testicular, donde los valores de pH y HCO3 son menores que en el conducto espermático y al tratar de inducir activación no se logró (26), lo cual conduce a pensar que las células epiteliales del conducto espermático ejercen el control sobre la capacidad de adquirir movilidad del espermatozoide, posiblemente regulando el pH del semen por secreciones ácido / bases (16).
El balance iónico también juega un papel en la adquisición de la capacidad para la activación de la movilidad y está relacionado con los cambios en las concentraciones extracelulares de K+, Na+, HCO3 , y/o iones hidrógeno (27). De lo anterior se observa que estos mecanismos aún se encuentran lejos de ser dilucidados completamente y se destaca la necesidad de investigación con el fin de poder tomar estas células espermáticas como modelos de células inducibles por factores fisicoquímicos.
Inhibición de la activación a nivel testicular
Los espermatozoides de los peces son inmóviles en el testículo (25), y en muchas especies también en el plasma seminal. Los espermatozoides adquieren progresivamente el potencial de movilidad a través de su pasaje por el conducto espermático, permaneciendo inmóviles hasta ser liberados al medio acuoso donde diferentes factores interactúan para desencadenar una respuesta que produce la activación de la movilidad. Los espermatozoides de los peces al ser liberados al medio ambiente acuoso, deben responder a condiciones fisicoquímicas como: cambios en la presión osmótica, balance iónico y pH (4).
Una vez terminada la espermatogénesis, los espermatozoides son liberados al conducto espermático y son bañados por un fluido testicular donde permanecen inmóviles y con un metabolismo bajo, las mitocondrias se encuentran con bajo potencial de membrana con el fin de preservar las pocas reservas energéticas, y disminuir la formación de compuestos de oxidación endógenos que pondrían en riesgo la integridad de membranas o compuestos citoplásmicos. Esta fase quiescente puede ser de diferente duración debido a la actividad sexual estacionaria de los peces, y termina cuando el semen es liberado en un medio externo donde el espermatozoide adquiere metabolismo activo y por consiguiente la movilidad (13).
]]> Los factores de inhibición de la movilidad en los conductos espermáticos en peces de agua dulce son: la osmolaridad en ciprínidos (18, 21), la presión osmótica, concentración de K+, concentración de sucrosa, y pH del plasma seminal menor a 7.0 en salmónidos (3, 6, 22) y una proteína con una tasa de migración (Migration rate) Mr = 120.000 específica del plasma seminal en algunos cíclidos (3, 22). Estos mecanismos se han venido clarificando cada vez más debido a su importancia en la reproducción de peces y muestran la diversidad adaptativa de las especies ante condiciones cambiantes, es probable que estos no sean los únicos factores involucrados, sin embargo, la falta de investigación en más grupos taxonómicos no permite elaborar conclusiones al respecto.Los factores que suprimen la movilidad son neutralizados en el momento de la espermiación por las condiciones del medio ambiente, así, los iones K+ en el plasma seminal de salmónidos o esturiones inhiben la movilidad espermática; este efecto inhibitorio se supera cuando el semen se diluye en el medio acuoso (4).
En otras especies soluciones fisiológicas electrolíticas que contienen K+ incluso a concentraciones altas, no inhiben la movilidad; ya que ocurren interacciones con otros componentes del medio externo como Na+, Ca2+ y Mg2+ (4). Se cree que hay antagonismo entre la concentración de Ca2+ y K+, y es así, como en espermatozoides de carpa, las concentraciones altas de K+ extracelular (más de 30 mM) y un pH extracelular menor de 7.5 inhiben la movilidad espermática; no obstante, son reversadas por la adición de Ca2+ (13). Además de esto, el plasma seminal contiene componentes de bajo peso molecular no identificados, que pueden inhibir la captación del calcio por lo espermatozoides, lo cual constituye un mecanismo de inhibición (7), sugiriendo que la interacción entre los diferentes componentes del medio es importante en la activación de este evento fisiológico.
La tilapia nilótica (Oreochromis niloticus) carece de conducto espermático principal, y como sus espermatozoides son móviles una vez producidos, ni el K+ ni la osmolaridad suprimen su movilidad en el testículo, lo que la inhibe es una proteína de Mr = 120.000, presente en el plasma seminal, que hace parte del factor de inmovilización del espermatozoide. Esta proteína secretada por las células de Sertoli se localiza en la cabeza del espermatozoide y en el plasma seminal formando polímeros que producen un ambiente altamente viscoso que impide la movilidad (24).
Para otras especies como algunos esturiones (Acipenser sp.) y salmónidos como la trucha (Salmo trutta), los ambientes con alta tensión de CO2 (hipercapnia), desestabilizan el balance óxido-reductor produciendo acidosis metabólica y disrupción en la función respiratoria de la mitocondria; lo cual a su vez inhibe la adquisición de la capacidad de movilidad en los espermatozoides dentro de los conductos espermáticos (16). Este es también el mecanismo que se cree actúa para inhibir la movilidad de los espermatozoides del pez escorpión (Cottus hangiongensis), ya que estos son móviles in vitro en el plasma seminal aún cuando no son liberados en un medio acuso, el factor de inhibición de la movilidad en los conductos espermáticos, sería también la alta tensión de CO2 (27).
Activación
En el momento de la espermiación, el semen es diluido en el agua desencadenándose una serie de eventos que conducen a la activación (4,22) mediada por factores como: choque hiposmótico (18), balance iónico (23), pH (27), oxido nítrico (14), AMPc (28), radicales libres (28) y calmodulina (19).
Choque hiposmótico. La movilidad espermática en peces de agua dulce y en peces marinos es parcialmente controlada por la presión osmótica (21). En dulceacuícolas, el agua tiene una baja osmolaridad en comparación con el plasma seminal, así, ocurre un choque hiposmótico que genera la señal inicial para los eventos que conducen a la activación (4), y que en la mayoría de peces de agua dulce tiene un rango de duración entre 30-40 segundos (23).
El choque hiposmótico produce una respuesta celular, resultando en la activación de la movilidad flagelar de las células espermáticas y también en un incremento del pH intracelular (23) alterado por el incremento o disminución de la concentración iónica interna. El flujo transmembranal de agua se da más rápidamente en espermatozoides de trucha y rodaballo (Psetta maxima) gracias a la estructura en aleta de la membrana flagelar (4) que en espermatozoides con flagelo cilíndrico como es el caso del tetra de lomo rojo (Brycon affinis) (1), en el primer caso, el plegamiento de esta membrana sobre el flagelo aplanado le provee al espermatozoide una superficie de contacto mayor, y así, permite la movilización de agua rápidamente (influjo en dulceacuícolas o eflujo en peces marinos) y facilita la adaptación del volumen intracelular (4).
La integridad de la membrana es especie específica y necesaria para que haya una permeabilidad diferencial de agua, iones y compuestos en solución (5), pero aún se desconoce la señalización transmembranal que induce la iniciación de la movilidad por la presión osmótica (19). El principal efecto del choque hiposmótico en la cascada de la iniciación de la activación de la movilidad del espermatozoide es inducir una inmediata hiperpolarización de la membrana (4) lo cual conlleva a un desarrollo progresivo de la activación de la movilidad.
]]> Este mecanismo es variable entre diferentes grupos taxonómicos dependiendo de las adaptaciones evolutivas al medio en el cual viven, incluso, se pueden encontrar diferencias en el umbral de osmolaridad que induce la iniciación de la movilidad entre células espermáticas individuales en el eyaculado de un mismo espécimen (27), lo que permite pensar que este es un proceso versátil que favorece la reproducción de los peces en condiciones ambientales variables donde el rango dependerá de las variaciones adaptativas entre e intra especies.En el pez remo (Polyodon spathula) donde la actividad está inhibida por altas tensiones de CO2, la osmolaridad del fluido seminal también puede influir en la velocidad de avance del espermatozoide (21), aunque se desconoce cual es exactamente la relación entre los dos eventos.
Soluciones hipotónicas median la iniciación de la movilidad en espermatozoides de peces de agua dulce tales como la carpa común (Cyprinus carpio), el pez dorado (Carassius auratus), la carpa de aleta roja (Tribolodum hakonensis) (4) y el pez remo (Polyodon spathula) (21). El rango óptimo se encuentra entre 100 y 200 miliosmoles (mOsm), por debajo de este rango se activan, pero la duración de la movilidad es menor; esto sugiere que la fuerza de batido podría ser más importante que la duración de la movilidad (4). Los espermatozoides se activan aún a 0 mOsm, sin embargo en medios por debajo de 75 mOsm se producen daños estructurales de membrana (23) morfológicamente reversibles cuando la osmolaridad es corregida (4, 23). Esto se comprobó en espermatozoides de pez zebra (Brachydanio rerio) realizando reactivación repetidas veces, sometiéndolos a diferentes osmolaridades por intervalos breves de tiempo (4, 27). Cuando el choque es prolongado se modifica la permeabilidad de la membrana y la organización de la bicapa lipídica, terminando en ruptura de la misma y muerte celular (19).
Medios hipotónicos, isotónicos y en cierto grado hipertónicos, a base de manitol, sucrosa o glucosa, de osmolaridad variada, inducen movilidad en algunos salmónidos. La osmolaridad entre 100 y 200 mOsm produce periodos más largos de movilidad, mientras que ésta declina en osmolaridades extremas de 0 ó 300 mOsm. Aún a osmolaridades de 450 mOsm se da un breve periodo de movilidad, sugiriendo que el choque hiposmótico no es el único factor de activación (4).
La activación de la movilidad en carpa, es dependiente de los cambios de osmolaridad del medio que los rodea. Estos espermatozoides son quiescentes en un medio isosmótico al plasma seminal no importando su composición, y totalmente activados en un medio diluyente donde la osmolaridad sea menor o igual a 150 mOsm (13); en medios hiposmóticos con cualquier composición iónica se dispara la activación al provocar un estiramiento de las células, lo que desencadena la movilidad por apertura de canales iónicos (4).
Hay evidencia de al menos dos clases de activadores osmóticos (switch) para la iniciación de la movilidad; un activador reacciona en respuesta a la presión osmótica por encima de 300 mOsm, con un umbral diferente para cada célula; el otro activador común para todas las células espermáticas, reacciona en respuesta a la presión osmótica por debajo de 200 mOsm (27).
Así, se ha encontrado que el choque hiposmótico es el primer factor relacionado con la activación de la movilidad en los peces de agua dulce, los resultados contradictorios entre algunas especies sugieren que éste no es el único factor de regulación y que existen otros factores que actúan en conjunto con el choque hiposmótico.
Potencial de membrana. El potencial transmembranal juega un papel importante en las señales de transducción y diferenciación en muchas células, ya que los cambios ocurren pocos segundos después de la unión de un ligando a un receptor apropiado, y estos median una respuesta fisiológica y metabólica subsecuente en las células involucradas (9); al sufrir el choque hiposmótico se hiperpolariza la membrana espermática y genera una reacción traducida en activación. La hiperpolarización de la membrana se puede producir a partir del choque hiposmótico o la combinación de éste con otros factores ambientales como pH y balance iónico (3), e incluso el factor de activación del huevo producido por la hembra (4).
La medida absoluta del potencial de membrana puede ser estimada en escala de milivoltios (mV) por medio de fluorescencia en citometría de flujo, tal como fue descrito por Miklós Emri (9), o por métodos electrofisiológicos como lo describió Zoltán Krasznai (17).
En la carpa, los espermatozoides inmóviles tienen un potencial de membrana bajo, debido a que la carga es mayor en el plasma seminal (82.4 mM de K+) que a nivel intracelular (60.5 mM de K+). En la iniciación de la movilidad inducida por osmolaridad, disminuye la presión osmótica extracelular y la concentración de K+; resultando en una hiperpolarización transitoria de la membrana (19).
]]> La activación no sólo está relacionada con la hiperpolarización transitoria de la membrana sino también, con la formación de complejos supramoleculares que influyen en la modificación de la fluidez lateral, despliegue enzimático y activación de canales iónicos (22). Así, el cambio de potencial de membrana generado por el choque hiposmótico es el segundo factor involucrado con la activación de los espermatozoides y puede desencadenar respuestas celulares que discutiremos a continuación.Canales iónicos. Según el mecanismo de activación se han identificado varios tipos de canales iónicos, siendo los mas estudiados en el espermatozoide, los activados mecánicamente y los dependientes de voltaje.
Los canales activados mecánicamente, son altamente sensibles a la tensión de membrana, lo cual modifica la conformación de algunas proteínas membranales (4) permitiendo el intercambio de iones con el medio extracelular, lo que produce un cambio en el potencial de membrana, activando los canales dependientes de voltaje. De esta forma, el Ca2+ ingresa al espermatozoide, y participa en la regulación del inicio de la activación (19); desencadenando la transducción de señales y actuando como enzima efectora a través de la adenilato ciclasa, la cual regula el metabolismo del AMPc (28).
El espermatozoide de carpa (Cyprinus carpio), cuenta con un número limitado de canales de K+, por lo cual la apertura o cierre de un solo canal implica la hiperpolarización o despolarización transitoria local de la membrana plasmática, lo cual resulta en un influjo de Ca2+ que conduce a la activación de la movilidad flagelar, de esta forma el incremento de Ca2+ intracelular en dos tercios en los primeros segundos constituye una señal para la iniciación de la movilidad espermática (19). Sin embargo el mecanismo exacto de señalización, aún no está bien definido.
Ensayos en carpa mostraron, que los inhibidores de canales de Na+ y los inhibidores de canales aniónicos no afectaron la movilidad; pero el bloqueador de canales de K+ (4-aminopiridina) la disminuye o suprime de forma reversible y dependiendo de la dosis y el tiempo de incubación (18), sugiriendo que el ingreso de K+ es un evento necesario corriente arriba, en la cascada de señalización mediada por Ca2+.
Así pues, los bloqueadores de Ca2+ (verapamilo) y K+ (4 aminopiridina) bloquean completamente el aumento de Ca2+ intracelular y la movilidad espermática, sin embargo, la movilidad se recupera si los bloqueadores son removidos por lavado (19).
Se sabe de la participación de otros canales iónicos en el proceso de activación espermática en peces teleósteos, no obstante, su importancia aún no es clara, es así como en estudios conducidos por Teréz Márián et al (23), se concluyó que los intercambiadores de Na+/H+ son inactivos bajo condiciones fisiológicas de 305 mOsm (23) por lo cual su efecto en la activación de la movilidad, está por aclararse.
La cascada de eventos que conduce a la activación de la motilidad es compleja e involucra el cambio en el balance iónico dado por la regulación de canales e intercambiadores. Harían falta nuevas investigaciones, para comprender mejor la complejidad de la membrana, de los canales y de los eventos que en conjunto los regulan.
Balance iónico. Los iones más ampliamente estudiados en la activación de la movilidad de los peces de agua dulce son el K+ y el Ca2+. Se ha observado, que a partir de la osmolaridad, el cambio en el balance iónico es el que desencadena la movilidad de los espermatozoides (4).
El mecanismo de regulación iónica está dado por la interacción entre osmolaridad, potencial de membrana y canales iónicos, así como, por el CO2 disuelto en equilibrio con el NaHCO3-que también contribuyen a la concentración iónica y a la osmolaridad (4).
]]> Los iones K+ inhiben la activación de los espermatozoides a baja concentración; 0.01 mM en esturiones (4) y 0.5 mM en pez remo (Polyodon spathula) (21). De acuerdo a ésto, se ha reportado que la inhibición de la acción del K+ es regulada principalmente por iones Ca2+, posiblemente debido al flujo simultáneo de Ca2+ y K+ (4); la salida de K+ favorece la apertura de canales de Ca2+, y el ingreso del mismo hacia el interior; el Ca2+ que ingresa, favorece la liberación de Ca2+ intracelular almacenado y modifica el pHi (pH intracelular) produciéndose la primera señal Ca2+ dependiente y AMPc independiente para el inicio de la movilidad (19).El modelo de activación de la movilidad en los espermatozoides, al menos para carpa es como sigue: los espermatozoides se encuentran en el semen en un ambiente isosmótico donde la concentración de K+ es mayor en el exterior que en el interior de la célula; una vez los espermatozoides son liberados, se enfrentan a un choque hiposmótico donde la concentración de K+ extracelular es disminuida; esto activa los canales de K+ y ocurre un eflujo que polariza la membrana transitoriamente, posteriormente, la membrana se despolariza activando los canales de Ca2+, lo que lleva a un cambio del balance iónico (19) desencadenando una cascada de eventos que involucran la señalización de membrana, seguida por la fosforilación de proteínas dependiente de AMPc (4); sin embargo en la trucha, otros iones divalentes como Cs2+, Sr2+ y Ba2+, también indujeron la movilidad del espermatozoide de forma similar al Ca2+ (4), mostrando que el mecanismo es complejo e involucra diferentes factores que actúan en conjunto y no de forma aislada para lograr la activación.
Algunos estudios reportan interacción entre Ca2+ y K+, ya que al adicionar pequeñas cantidades de Ca2+ a soluciones de K+ que no activaban espermatozoides de truchas, se induce la movilidad (4). Este mismo efecto se observó en espermatozoides de pez remo (Polyodon spathula), en el cual el movimiento no está influenciado por la presión osmótica, pero sí, por el control recíproco de las concentraciones de Ca2+ y K+ (21). Por otra parte, en espermatozoides de trucha, los iones Na+ y Mg2+; también participan en el proceso de activación, reduciendo la acción inhibitoria de los iones K+ (3).
Debido a que el influjo de Ca2+ es prerrequisito para la activación de espermatozoides, en algunos teleósteos la iniciación de la movilidad no puede ser producida en soluciones hipoosmóticas libres de Ca2+ (19), contrario a esto el espermatozoide de carpa no es sensible al Ca2+ externo; y la movilidad puede darse aún cuando se bloquean los canales de Ca2+, sin embargo, espermatozoides desmembranados no pueden activarse con ATP en ausencia total de Ca2+, pero recuperan su actividad si el Ca2+ es adicionado (4) evidenciando la función de este ion en el proceso de activación.
Así, el Ca2+ es efector de la movilidad espermática en esturiones, tilapia, róbalo, pez remo y roncacho (Micropogonias undulatus), pero en silúridos, rodaballos, lenguados y anguilas, su función no parece ser importante para este proceso (4), sugiriendo que hay mecanismos aún no descubiertos independientes de Ca2+ que pueden iniciar la movilidad espermática.
PH. El pH extracelular es otro de los factores que controlaría los parámetros de movilidad, y sus valores óptimos son necesarios pero no suficientes para las condiciones de activación espermática (23), alteraciones del pH interno (un posible segundo mensajero celular) interfieren con la movilidad en diferentes especies (4). Por lo tanto, la modificación del pH intracelular por acción del Ca2+ y otros componentes parecen ser necesarios en el proceso de activación.
La proporción de movilidad espermática disminuye abruptamente hasta 0% cuando el pHi o extracelular se aproxima a valores de 5.4 y/o 5.0 respectivamente (23). El cambio de pHi depende en gran parte del flujo de iones a través de la membrana, y se cree que genera un medio adecuado para la interacción de diferentes compuestos involucrados con la activación (28), la inhibición de canales de K+ afecta la movilidad pero no el cambio de pHi, en contraste, los inhibidores de canales de Na+ no afectan la movilidad manteniendo constante el pHi, lo cual sugiere que el pHi no juega un papel esencial en la activación de la movilidad (18). Teréz Márián et al (23), encontraron que la activación esta acompañada por una rápida alcalinización de las células espermáticas (23), relacionada con la apertura de canales de Ca2+ y entrada del mismo al citoplasma. Además, el pH no se alteró con el uso de bloqueadores de canales de Na+ (amiloride), sugiriendo que el intercambiador de Na+/H+ sí juega un papel en el proceso de activación (23), por otra parte, al usar bloqueadores de canales de K+ como la 4aminopiridina la medida de amplitud y la cinética del cambio de pH interno fue igual que en la ausencia de este inhibidor (18), quedando sin respuesta clara la pregunta acerca del papel del pH en la regulación de la activación.
Algunos estudios han reportado que el pH extracelular por debajo de 5.5 inhibe completamente la movilidad espermática; pero el pH alcalino extracelular no alteró la duración de la movilidad (6,9). En trucha arco iris (Oncorhynchus mikiss) la activación de la movilidad ocurre cuando el pH del medio de incubación se aumenta por encima de ocho (27). La movilidad flagelar depende también del pH intracelular en un rango de 6.5 - 8.5, por encima o debajo de este rango, se reduce significativamente (23).
La regulación del pH intracelular; incluye el intercambio de Na+/H+ y K+/H+ a nivel de la membrana plasmática y el cotransporte de NaCl y HCO3, y ATPasas e H+; por lo tanto, el único factor aparente de regulación del pHi parece ser el potencial de membrana plasmática el cual en si mismo es dependiente en parte del pHe y del mismo pHi, por lo cual el pHi es dependiente del pH extracelular (13) de esta forma, los efectos del pH sobre la movilidad espermática son parcialmente reversibles (23) siempre y cuando no se comprometa la permeabilidad de membrana. Lo anterior muestra nuevamente la complejidad de los eventos que actúan de forma orquestada en la regulación del inicio de la activación de la movilidad.
La temperatura del medio es un factor que altera diversas características en el semen; en un estudio realizado por Emri et al (8) tomando carpas adaptadas al frío y al clima medio, encontraron que el pH del plasma seminal de los animales adaptados al frío fue de 8.6 siendo mayor al pH del plasma seminal de los animales del clima medio el cual tuvo un valor de 8.3; la concentración de Na+ en plasma seminal de los animales adaptados al frío (83mM) fue más alta, comparado con la de los adaptados al clima medio (63mM); la concentración de K+ en los de clima frío fue más baja (64mM) comparándola con los de clima medio (87mM); sin embargo la fracción mótil y la duración de la movilidad de ambos fue idéntica; sugiriendo que los espermatozoides de estos peces se adaptan a diferentes rangos en los parámetros del medio (8).
]]> La medida del pH intracelular puede llevarse a cado mediante resonancia magnética nuclear, por la prueba de fluorescencia 9 amino-acridina y también por la distribución de radiación en pruebas radioactivas (13).Teréz Márián et al (23) sugieren que el pH intracelular no tiene un papel regulatorio primario sobre la inducción de la movilidad espermática, no obstante se ha visto que juega algún papel en el proceso de activación de los espermatozoides, al igual que otros parámetros menos conocidos, por lo cual es conveniente que se desarrollen nuevos estudios integrando la mayor cantidad de factores para tratar de comprender en conjunto la relación y función de ellos en este evento, el cual es de importancia para el desarrollo de tecnologías que permitan la reproducción de especies en cautiverio bajo diferentes condiciones ambientales.
Otros factores de activación de movilidad. En algunas especies se han estudiado otros factores de activación de la movilidad, por ejemplo, la movilidad de los espermatozoides de salmón es aumentada por el fluido ovárico, el cual es liberado con los huevos, la duración de la movilidad en estas condiciones es hasta el doble de la encontrada en ausencia del mismo. Algunos factores promotores de movilidad encontrados en el fluido ovárico incluyen: astaxantina, beta carotenos o sustancias inespecíficas de bajo peso molecular (4).
También se sabe de otras sustancias biológicas que aumentan la movilidad y representan parte integral del proceso de fertilización, en los peces vivíparos, la movilidad se aumenta por la presencia de un azúcar reducible; además, en algunos peces como en el arenque (Clupea harengus) ocurre atracción del espermatozoide por sustancias extracelulares presentes en la región micropilar, las cuales son llamadas factor promotor de la movilidad, y son un componente menor del corion del huevo (4).
La temperatura del medio de igual forma afecta el periodo de movilidad, es así como bajas temperaturas resultan en prolongados periodos de duración de la movilidad, pero con una reducida velocidad (4).
El AMPc juega un papel importante en la activación de espermatozoides en algunas especies, se ha observado que en espermatozoides de trucha tanto las concentraciones intracelulares de AMPc como las de Ca2+ aumentan después de la activación (4); sin embargo en espermatozoides de carpa la concentración de iones de Ca2+ aumentó después de la activación, pero los niveles de AMPc no cambiaron, y los inhibidores de proteinas-kinasas no afectaron la movilidad, sugiriendo que no es necesario un sistema dependiente de AMPc para la regulación de la movilidad espermática en carpa (4,19).
Las prostaglandinas y hormonas esteroideas que alteran los niveles de AMPc en otros tejidos pueden influenciar la movilidad en el plasma seminal afectando los niveles de AMPc espermáticos y la actividad de la adenilato-ciclasa (7).
El AMPc y su subsecuente cascada de fosforilación de proteínas en el proceso de la movilidad de los espermatozoides parece estar restringido solo a salmónidos (4). Así, la forma de regulación del metabolismo del AMPc puede estar relacionada con el cambio en el flujo de los iones Ca2+ y HCO3-, ya que ambos son capaces de estimular la actividad de la adenilato-ciclasa (AC) en los espermatozoides, pero los mecanismos de como se produce esta activación se desconocen (28).
El hecho de que en algunos peces como el guppy (Poecilia reticulata), los espermatozoides sean liberados en grupos (espermatozeuma), y la movilidad se inicie espontáneamente con la disociación de la espermatozeuma, evidencia que ni el balance iónico, ni la osmolaridad tienen efecto sobre el inicio de la movilidad en esta especie, lo cual sugiere la existencia de otros mecanismos de inhibición y activación espermática aún desconocidos (4).
Se hace evidente que son muchos los factores involucrados en la regulación de la activación de la movilidad en los peces de agua dulce, sin embargo, sea cual sea el mecanismo particular de cada especie, el propósito final de la cascada de señalización, será activar la maquinaria celular necesaria para mover el flagelo he impulsar al espermatozoide hacia delante en su camino hacia el huevo.
]]> Actividad flagelarLa movilidad es uno de los mayores indicadores de funcionalidad; en ella están involucrados el flagelo, el complejo axonema y la maquinaria energética (mitocondrias) ubicadas en forma helicoidal en la pieza media (5).
Entre los peces con fertilización externa, las características de comportamiento de la movilidad del flagelo son muy similares en muchos aspectos, con excepción del espermatozoide de anguila el cual puede nadar por largos periodos de tiempo, con un patrón de onda diferente, moviéndose en forma de rollo (3).
Se ha observado un efecto de la osmolaridad sobre el axonema mediante la desmembranación y reactivación del espermatozoide de carpa; en soluciones iónicas que solamente contenían Mg2+ y ATP, se mantuvo la movilidad hasta una osmolaridad de 550 mOsm mientras que los espermatozoides con membrana perdieron movilidad desde los 250 mOsm (4). La desmembranación se obtiene por la aplicación de un medio detergente no iónico (Como TritonX100) y la reactivación es reiniciada por la adición de ATP-Mg2+, al sustrato de dineína ATPasa del flagelo (21).
Cambios drásticos en el ambiente osmótico al cual se enfrenta el espermatozoide al diluirse el plasma seminal, puede afectar el transporte de ATP hacia la parte distal del flagelo; disminuyendo la amplitud de la onda (3). La forma de la onda del flagelo puede ser afectada por la concentración iónica, osmolaridad y CO2 (4).
Los mecanismos moleculares por los cuales se inicia la movilidad no están totalmente explicados (23); se relacionan usualmente el ATP, como proveedor de energía, AMPc como mensajero y factores ambientales no determinados que activan el sistema ATP Mg2+ (4) sin embargo es claro que, la energía (ATP) requerida para la movilidad es producida por las mitocondrias en la pieza media (3); además, el mantenimiento de dicha energía también se ha relacionado con la concentración y el radio de iones en el fluido seminal (21).
Los factores mejor caracterizados para la señalización de la cascada en la regulación de la movilidad espermática son el AMPc y el Ca2+, así, la fosforilación de las proteínas del axonema dependientes de AMPc, regulan la movilidad del espermatozoide en salmónidos. El Ca2+ posiblemente actúa como cofactor de proteinas-kinasas o de fosfatasas importantes para la movilidad. Los movimientos flagelares en células eucarióticas son causados por el deslizamiento activo de los brazos externos de dineína de los pares de microtúbulos externos del axonema (15). Cada par de microtúbulos periféricos del axonema tiene un túbulo A y un túbulo B, el A lleva dos brazos de dineínas (ATPasas) los cuales se disponen formando rutas a lo largo del microtúbulo. Bajo hidrólisis de ATP, estos brazos de dineína interactúan con la tubulina del túbulo B causando un deslizamiento (4, 7), que conduce al plegamiento flagelar y la propagación de la onda en conjunto con otras estructuras del axonema como el par central y los radios (15); durante el movimiento, la tubulina espermática se encuentra fosforilada de una manera AMPc dependiente y esto se correlaciona con un incremento en la movilidad espermática (7). Lo anterior podría explicar en parte algunas de las rutas de señalización planteadas previamente, las cuales involucran al Ca2+ corriente arriba y al AMPc corriente abajo.
Sin embargo, se ha visto que en salmónidos existen otros mecanismos regulatorios para la movilidad espermática: uno, dependiente de la fosforilación de una proteína con una banda de 15 kDa por una tirosina-kinasa en la parte basal del flagelo y el segundo dependiente de la fosforilación de una proteína con una banda de 22 kDa por una proteina kinasa dependiente de AMPc a lo largo del flagelo del espermatozoide; los proteosomas pueden jugar un papel clave en la activación de estas fosforilaciones ya que su actividad es regulada por proteinas kinasas y por Ca2+ cuando se inicia la movilidad del espermatozoide. La red enzimática integral responsable de la activación del deslizamiento de los microtúbulos esta constituida por el proteosoma, las proteinas kinasas dependientes de AMPc y los brazos externos de dineina; se sabe además por microscopia inmunoelectrónica que los proteososmas están ubicados cerca de los brazos externos de dineína (15) apoyando así esta hipótesis.
El principal papel del AMPc en el espermatozoide es mediar la fosforilación de proteínas esenciales para iniciar o mantener la movilidad. En el espermatozoide de mamíferos el ATP es convertido a AMPc por una adenilciclasa dependiente de Ca2+ que puede ser controlada parcialmente por la calmodulina; múltiples genes que codifican calmodulina son expresados durante la espermatogénesis (7). Es posible que la fosforilación de la proteína con una banda de 22 Kda pueda estar relacionada estrechamente con los rápidos cambios en la movilidad como es la transición del estado inmóvil al estado móvil en el primer segundo cuando los espermatozoides son liberados al medio (15).
Conclusiones ]]> La reproducción de teleósteos particularmente de salmónidos y ciprínidos, ha sido estudiada por largo tiempo debido a su interés económico; las dos especies tienen fertilización externa y su espermiación ocurre en agua dulce, siendo la activación de la movilidad espermática dependiente de factores externos diferentes en ambos grupos: en salmónidos los iones externos H+, K+, Ca2+, y en ciprínidos, la presión osmótica (13). Sin embargo, los mecanismos moleculares involucrados en la activación espermática no son bien conocidos (23).
Se ha planteado un mecanismo general de activación espermática en el cual el choque hiposmótico y el pH extracelular actúan sobre la movilidad de los espermatozoides mediante su efecto sobre el potencial transmembranal; cuando el pH extracelular es bajo, la membrana plasmática se despolariza, al igual que cuando las concentraciones externas de K+ están elevadas (4,13). Por otra parte, durante la movilidad espermática, ocurre un incremento en las concentraciones internas de Ca2+ debido a la apertura de los canales del mismo dependientes de voltaje o un desplazamiento de Ca2+ desde la membrana o desde reservas internas; así, los efectos inhibitorios del pH extracelular y el K+ pueden ser sorteados por la adición de iones divalentes al medio externo que median un efecto sobre la carga de superficie de la membrana plasmática y también sobre su potencial electro químico. Estos cambios del potencial de membrana y el Ca2+ interno al momento de la dilución son también acompañados por: aumento en el AMPc, fosforilación de una proteína de membrana de15 kDa (13) y aumento del pHi (4). El AMPc estimula la actividad dineína-ATPasa y causa el movimiento del axonema (13).
El movimiento flagelar es entonces el resultado final de una compleja red de interacciones y reacciones celulares ante cambios de su medio ambiente interno y externo, se evidencian las diferencias evolutivas de adaptación y la necesidad de nuevas investigaciones que conduzcan a la comprensión global del proceso.
Perspectivas
La investigación básica dirigida a la comprensión de las intrincadas relaciones entre los espermatozoides como células altamente especializadas, el entorno circundante, las respuestas internas y la señalización que conducen finalmente a la activación e inicio de la movilidad, es sin duda fundamental para avanzar en el entendimiento de la biología de las especies, la vida misma, la evolución y el comportamiento.
Falta mucho terreno por recorrer en el campo de la investigación sobre los mecanismos de activación espermática en peces, y aún quedan infinidad de factores por estudiar, como aquellos presentes en el agua misma, que podrían estar regulando los ciclos reproductivos de los peces, el balance hormonal y su relación con el medio ambiente y hasta el mismo comportamiento de los peces y sus comunidades, lo cual, sólo integrándolo permitirá aclarar conceptos que permitan intervenir de forma segura en el campo de la conservación, la investigación o la producción.
Summary
Spermatozoa activation physiology in freshwater fishThis review pretended to study the regulation of spermatozoa motility in freshwater fish, improving biotechnology development focused on gametes preservation, which are produced into the testes. Spermatozoa gets capacitation and stays non- motil due to different factors such as ionic balance, osmolality, pH, and some proteins. After release to the freshwater, the teleost spermatozoa get activated during a very short period, due to hiposmotic shock, producing membrane potential changes, allow regulation of K+ and Ca2+ channels, followed by an ionic flow that trigger the activation. During the last years, investigations focused on the osmolality, ionic balance, pH and AMPc and Ca2+ dependent factors; associated to spermatozoa motility activation. In this way, understanding spermatozoa hipermotility process to reach and enter the egg´s micropile before closing, has increased interest in the physiological activation mechanisms
Key words: hiposmotic shock, ionic balance, motility, osmolarity.
]]>Referencias
1. Andrade RF, Bazzoli N, Rizzo E, Sato Y. Continuous gametogenesis in the neotropical freshwater teleost, Bryconops affinis (Pisces: Characidae). Tissue & Cell 2001; 33:524-532. [ Links ]
2. Corley-Smith G, Brandhorst B. Preservation of endangered species and populations: A role for genome banking, somatic cell cloning, and androgénesis. Mol Reprod and Dev 1999; 53: 363-367. [ Links ]
3. Cosson J, Linhart O, Mims SD, Shelton WL, Rodina M. Analysis of motility parameters from paddlefish and shovelnose sturgeon spermatozoa. J Fish Biol 2000; 56: 1348- 1367. [ Links ]
4. Cosson J, Billard R, Cibert C, Dréanno C. Ionic factors regulating the motility of fish sperm. In: Gagnon C, editor. The Male Gamete: from basic knowledge to clinical applications. Vienna: Cache river press; 1999. p. 161-186. [ Links ]
5. Critser JK, Noiles EE. Bioassays of sperm function. Seminars in reproductive endocrinology 1993;11:1-16. [ Links ]
6. Darszon A, Labarca P, Nishigaki T, Espinosa F. Ion Channels in Sperm Physiology. Phys Rev 1999;79:481-510. [ Links ]
7. Eddy EM, O´Brien DA. The espermatozoon. In: Knobil E, Neill JD, editors. 2nd edition. The Physiology of Reproduction. New York: Raven Press; 1994. p.29-77. [ Links ]
8. Emri M, Marian T, Trón L, Balkay L, Krasznai Z. Temperature adaptation changes ion concentrations in spermatozoa and seminal plasma of common carp without affecting sperm motility. Aquaculture 1998;167:85-94. [ Links ]
9. Emri M, Balkay L, Krasznai Z, Trón L, Márián T. Wide applicability of a flow cytometric assay to measure absolute absolute membrane potentials on the millivolt scale. Eur Biophys J 1998; 28: 78-83. [ Links ]
10. Frits MF, Baren MG. Dynamic of the mammalian sperm plasma membrane in the process of fertilization. Bioch et Bioph Acta 2000;1469:197-235. [ Links ]
11. Grassiotto Q, Negräu JN, Carvalho ED, Foresti F. Ultraestructure of spermatogenic cells and spermatozoa in Hoplias malabaricus (Teleostei, Characiformes, Erythrinidae) J Fish Biol 2001;59:1494-1502. [ Links ]
12. Gwo JC, Ohta H, Okusawa K, Wu HC. Cryopreservation of sperm from the endangered Formosan landlocked salmon (Oncorhynchus masou formosanus). Theriogenology 1999;51:569-582. [ Links ]
13. Hamamah S, Gatti JL. Role of the ionic environment and internal pH on sperm activity. Hum Reprod 1998;13 Suppl 4:20-30. [ Links ]
14. Herrero MB, Gagnon C. Nitric Oxide: A novel mediator of sperm function. J Andr 2001; 22:349-356. [ Links ]
15. Inaba K, Morisawa S, Morisawa M. Proteasomes regulate the motility of salmonid fish sperm through modulation of AMPc-dependent phosphorylation of an outer arm dynein light chain. J cell sci 1998;111: 1105-1115. [ Links ]
16. Ingerman RL, Holcomb M, Robinson ML, Cloud JG. Carbon dioxide and pH affect sperm motility of white sturgeon (Acipenser transmontanus). J Exp Biol 2002; 205:2885-2890. [ Links ]
17. Krasznai Z, Márián T, Balkay L, Emri M, Trón L. Flow cytometric determination of absolute membrane potential of cell. J Photoch and Photobiol 1995; 28:93-99. [ Links ]
18. Krasznai Z, Márián T, Balkay L, Gáspár Jr.R, Trón L. Potassium chanels regulate hypo-osmotic shock-induced motility of common carp (Cyprinus carpio) sperm. Aquaculture 1995; 129:123-128. [ Links ]
19. Krasznai Z, Márián T, Izumo H, Damjanovich S, Balkay L, et al. Membrane hiperpolarization removes inactivation of Ca2+ channels, leading to Ca influx and subsequent initiation of sperm motility in the common carp. PNAS 2000; 97: 2052- 2057. [ Links ]
20. Lahnsteiner F. Introduction to the special issue on cryopreservation of gametes in acuatic species. Aquac Res 2000; 3: 229. [ Links ]
21. Linhart O, Cosson J, Mims SD, Shelton WL, Rodina M. Effects of ions on the motility of fresh and demembranated paddlefish (Polyodon spathula) spermatozoa. Reprod 2002; 124: 713-719. [ Links ]
22. Márián T, Krasznai Z, Balkay L, Balázs M, Emri M, et al. Hypo-osmotic shock induces an osmolalitydependent permeabilization and structural changes in the membrane of carp sperm. J Histoch and Cytoch 1993; 41: 291-197. [ Links ]
23. Márián T, Krasznai Z, Balkay L, Emri M, Trón L. Role of extracellular and intracellular pH in carp sperm motility and modifications by hyperosmosis of regulation of the Na+ / H+ exchanger. Cytometry 1997; 27: 374-382. [ Links ]
24. Mochida K, Kondo T, Matsubara T, Adachi S, Yamauchi K. A high molecular weight glycoprotein in seminal plasma is a sperm immobilizing factor in the teleost Nile tilapia, Oreochromis niloticus. Dev Growth Differ 1999; 41: 619- 627. [ Links ]
25. Müller K, Labbé C, Zachowski A. Phospholipid transverse asymmetry in trout spermatozoa plasma membrane. Bioch et Bioph Acta 1994; 1192 :21-26. [ Links ]
26. Ohta H, Unuma T, Tsuji M, Yoshioka M, Kashiwagi M. Effects of bicarbonate ions and pH on acquisition and maintenance of potential for motility in ayu, Plecoglossus altivelis Temminck et Schlegel (osmeridae), spermatozoa. Aquac Res 2001; 32: 385-392. [ Links ]
27. Ohta H, Shinriki Y. Changes in osmotic pressure that trigger the initiation of sperm motility in the river sculpin Cottus hangiongensis. Fish Phys and Bioch 1998; 18: 29-35. [ Links ]
28. Sierra RA, Olivera M. Interacción entre gametos: como lo logra el espermatozoide?. Rev Col Cienc Pec 2000; 13: 148-155. [ Links ]
]]>