0120-06900120-0690S0120-06902007000200008ColombiaColombiaColombia0006200700062007202157173Modelación de funciones de crecimiento aplicadas a la producción animal
Modeling of growth functions applied to animal production
Divier A Agudelo-Gómez1,2,*, Ind Pecuario; Mario F Cerón-Muñoz M3, Zoot, PhD; Luis F Restrepo L2, Est, Esp.
1 Facultad de Ciencias Administrativas y Agropecuarias, Corporación Universitaria Lasallista. Caldas Antioquia, Colombia 2 Grupo GRICA, Facultad de Ciencias Agrarias e Instituto de Biología, Universidad de Antioquia. Medellín, Colombia 3 Grupo de Investigación en Genética y Mejoramiento Animal, Facultad de Ciencias Agrarias e Instituto de Biología, Universidad de Antioquia. Medellín, Colombia ]]>
diagudelo@lasallista.edu.co
El crecimiento animal es uno de los aspectos más importantes al momento de evaluar la productividad en las explotaciones dedicadas a la producción de carne y en algunos casos es usado como criterio de selección, sin embargo, debe tenerse en cuenta que el crecimiento no se debe exclusivamente a factores genéticos sino también, a efectos ambientales. Para medir el crecimiento animal se han usado diferentes modelos matemáticos lineales, no lineales y logarítmicos, entre otros, eligiéndolos por su bondad de ajuste y la facilidad de interpretación biológica de sus parámetros. Recientemente se han usado modelos mixtos en los que sus parámetros están compuestos de efectos fijos y efectos aleatorios, representando los valores esperados y la varianza de los primeros, respectivamente, lo que permite evaluar la variabilidad de las diferentes curvas entre los individuos de una población, así como la covarianza entre los parámetros. Los criterios más usados para elegir la curva que mejor ajusta a los datos son: el coeficiente de determinación, el porcentaje de curvas significativas y atípicas encontradas para cada función; además se pueden aplicar criterios como el criterio de información Akaike y el criterio de información Bayesiano. El objetivo del presente trabajo es indicarle al lector una aplicación de los modelos no lineales y no lineales mixtos en el análisis del crecimiento animal.
Palabras clave:biomodelación, curvas, modelo. ]]>
Summary Animal growth is one of the most important aspect for evaluating animal productivity in beef cattle enterprises and in some cases it is used as a criterion of selection, nevertheless, the fact that animal growth is not exclusively due to genetic factors but also, to environmental effects should be keep in mind. Measurement of animal growth have been performed by several logarithmic, not lineal, lineal, and mathematical models, having as selection criteria its fitness of adjustment and the feasibility for biological interpretation of their parameters. Recently the mixed models have been used in which their parameters are composed of fixed and random effects representing the expected values and variance of the fixed ones, respectively, which permits to evaluate the variability of different curves between individuals of apopulation, as well as the covariance between parameters. The most used criteria for selection of the curve that best fit data are: determination coefficient and the percent of significant and atypical curves found for each function. In addition, other models as the Akaike information criteria and Bayesian information criteria can also be applied. The objective of the present review is to provide the criteria for application of linear and non linear models when analyzing animal growth.
* Autor para el envío de la correspondencia y la solicitud de separatas. Facultad de Ciencias Administrativas y Agropecuarias, Corporación Universitaria Lasallista. Caldas Antioquia, Colombia. E-mail: diagudelo@lasallista.edu.co
El crecimiento animal puede ser descrito por medio de funciones matemáticas que predicen el desempeño de la evolución del peso vivo, dichas funciones permiten realizar evaluaciones sobre el nivel de producción en las empresas ganaderas, pudiendo clasificar de forma sencilla la productividad de una raza específica para una zona determinada (27).
También permiten calcular los valores máximos de los crecimientos medio y corriente, pudiendo determinar las edades de sacrificio que permitan obtener el máximo beneficio económico. Además, proveen información que permite realizar programaciones de alimentación, de capacidad de carga y medir cambios genéticos de una generación a otra que estén relacionados con el nivel de producción.
Las funciones que más se han usado son las propuestas por Gompertz en 1825 (12) Verhulst en 1838 (35) conocida como logística, la de Brody en 1945 (3), la de Von Bertalanffy en 1957 (36), y la de Richards en 1959 (31); además, se ha hecho uso de funciones lineales, y polinómicas entre otras, para tratar de predecir el peso adulto o el grado de madurez sin tener que realizar mediciones repetidas o esperar que el animal alcance su madurez para tomar decisiones relacionadas con la producción (21).
El desconocimiento de las curvas de crecimiento y de parámetros productivos de interés económico, ha limitado la implementación de programas de mejoramiento zootécnico que permitan aumentar la productividad, como lo son la velocidad de crecimiento, la tasa de madurez a diferentes edades y la edad al sacrificio. Estos factores se pueden analizar con base en la información zootécnica de los animales siendo indispensable para ello contar con registros de producción. ]]>
Es necesario realizar trabajos que permitan identificar las funciones matemáticas que mejor se ajusten a las condiciones locales, así como conocer los parámetros genéticos para realizar procesos de selección de los mejores animales.
Crecimiento animal
El crecimiento animal inicia en la etapa prenatal con la fecundación del óvulo y termina cuando el organismo alcanza el peso adulto y la conformación propia de la especie (13), se presenta un aumento cuantitativo de la masa corporal que se define como la ganancia de peso por unidad de tiempo (14). El aumento de peso se produce por tres causas: hiperplasia (multiplicación celular); hipertrofia (aumento del tamaño de las células) y metaplasia (transformación de las células), donde el crecimiento animal es una respuesta celular a diferentes factores que pueden ser inherentes al animal o ajenos a éste (13).
Si el proceso de crecimiento no tiene ningún factor inhibidor, normalmente el organismo sigue un mecanismo de multiplicación constante de las células, y una vez producida la multiplicación suficiente se produce la hipertrofia. Sin embargo, es posible que aparezcan factores inhibidores que detengan el proceso de hiperplasia y así el crecimiento se anula. ]]>
Si bien los diferentes sistemas se desarrollan en forma paralela, la velocidad a la que lo hacen es distinta, existiendo un orden estricto para su desarrollo; el primero en terminar el desarrollo es el nervioso, seguido del esqueleto, luego los músculos y por último se realiza la acumulación de tejido adiposo.
La evolución del aumento de peso vivo a lo largo de la vida de un animal es un fenómeno complejo que depende del genotipo del animal, de factores ambientales como la alimentación, el manejo, el estado de salud y efectos climatológicos, que tienen mayor impacto en las épocas iniciales del crecimiento (13); algunos de estos factores persisten en el tiempo y generan un efecto variable con la edad y el desarrollo del animal; otros por el contrario, pueden afectar sólo en periodos cortos. Los factores genotípicos inciden sobre el desarrollo fetal y se manifiestan desde el nacimiento hasta la adultez; la cría crece en forma lenta durante el primer mes posparto, pero después inicia una fase de un rápido crecimiento hasta alcanzar la pubertad, después de la cual disminuye la velocidad de crecimiento hasta llegar a la estabilización en la edad adulta. Debe tenerse en cuenta al comparar diferentes lotes de animales que las condiciones a las que fueron sometidos sean las mismas, que pertenezcan al mismo grupo genético así mismo se recomienda realizar grupos contemporáneos, las mediciones deben ser tomadas con los mismos intervalos de tiempo, también serecomienda tener lotes testigos; lo anterior está encaminado a la obtención de resultados y análisis más precisos (2, 14).
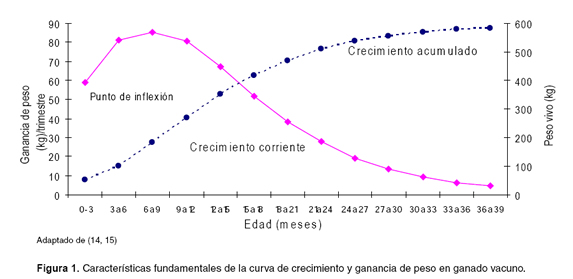
En la figura 1 se puede observar la evolución del peso del animal en el tiempo, así como la velocidad de crecimiento. Inicialmente la ganancia de peso es mayor que en la edad adulta, observándose una curva de evolución cóncava hacia arriba; a medida que se desarrolla el individuo la velocidad de crecimiento disminuye, y se presenta un cambio en la curvatura, identificando un punto de inflexión que corresponde con el valor máximo de la curva de crecimiento corriente. A partir de ese momento, el crecimiento es más lento haciendo que la curva de ganancia de peso disminuya paulatinamente y la curva de la evolución aumenta cada vez más despacio. Esta tendencia continúa hasta que cesa el crecimiento y se estabiliza el peso del individuo, hecho que corresponde matemáticamente con la asíntota horizontal (16).
La edad es uno de los factores más decisivos en el peso vivo de los animales, pues experimenta cambios progresivos desde el nacimiento hasta la adultez. La alimentación es el principal factor exógeno a considerar en el conjunto de particularidades ambientales gravitantes sobre el crecimiento. Para obtener efectos positivos, es necesario garantizar un suministro constante de alimento en la cantidad adecuada y de un valor nutricional que cumpla con las exigencias metabólicas de los animales (16). ]]>
Curvas de crecimiento
El crecimiento a lo largo de la vida de los animales se ha estudiado teniendo en cuenta la relación edad-peso, en los últimos años los estudios han hecho uso de los modelos no lineales, generándose diferentes modelos de crecimiento (1). Los modelos han sido evaluados según su bondad de ajuste, la interpretación biológica de los parámetros, la dificultad de aplicación informática y la evaluación de los parámetros ambientales sobre la curva de crecimiento (4).
Un modelo apropiado de crecimiento debe suministrar información sobre parámetros que pueden ser interpretados biológicamente; además, si se hace un buen uso del modelo se pueden obtener características que son de importancia en el crecimiento animal (18, 26, 29). Las funciones no lineales han sido usadas para describir el crecimiento en peces, aves, y mamíferos (16, 29). A pesar que la mayoría de funciones de crecimiento usadas explican las curvas de crecimiento no logran realizar una explicación clara sobre las tasas medias de crecimiento en la curva, pues en muchos casos el comportamiento del crecimiento varía de acuerdo con la etapa fisiológica en que se encuentre el animal viéndose afectada entre otros aspectos la ganancia diaria de peso, por lo anterior se hace necesario obtener funciones que se ajusten a todos los datos observados (18, 25).
Las funciones que normalmente se usan para evaluar el crecimiento son elegidas en forma empírica y su elección se hace con base a la habilidad de la función de ajustarse a los datos, y en ocasiones los parámetros obtenidos en dichas funciones no tienen una interpretación biológica. Sin embargo, las funciones de crecimiento pueden ser caracterizadas mediante el conocimiento de algunos delineamientos fisiológicos o mecanismos biológicos según lo propuesto por Von Bertalanffy, permitiendo que los parámetros obtenidos puedan ser analizados desde el punto de vista productivo (18, 23, 24). ]]>
Las curvas de crecimiento permiten evaluar parámetros biológicamente importantes: como es el tamaño del animal, evaluado como el peso al alcanzar la madurez sexual; otro es la relación entre la tasa de crecimiento con respecto a la tasa de maduración sexual. Estos parámetros sólo pueden ser evaluados una vez se haya completado el crecimiento. La estimación temprana de los factores puede servir para proponer programas de selección y mejoramiento pues son asociadas a otras características de importancia económica (11, 15, 23).
Funciones de crecimiento más empleadas
Las funciones o ecuaciones de crecimiento describen las variaciones que experimenten el tamaño de un organismo o una población con la edad. El crecimiento biológico es el resultado de un gran número de procesos complejos, puede ser descrito exponiendo la evolución del peso, sin que sea necesario realizar ajustes a las medidas tomadas. Un segundo paso es ajustar un polinomio a los datos, bien sea una ecuación cúbica o de potencias superiores; sin embargo, lo anterior no es aconsejable por tres razones: la primera, porque no siempre los coeficientes tienen una interpretación biológica; la segunda, porque el comportamiento de la ecuación puede ser ilógico al quedar fuera de la nube de puntos. Por ejemplo, la predicción del peso adulto podría disminuir con el tiempo, lo anterior no ocurre con las ecuaciones de las curvas de crecimiento. La tercera razón, es que el ajuste del polinomio es arbitrario; se supone que se va aumentando el grado del polinomio hasta que el ajuste sea satisfactorio, de hecho el polinomio de grado n-1, que describe la línea que une todos los puntos, tiene el ajuste más satisfactorio, aunque sea de poca utilidad para futuras aplicaciones (2, 22).
Otros limitantes que presentan los polinomios ordinarios son los siguientes: exhiben multicolinealidad, desuniformidad a lo largo de la curva, dependencia del comportamiento de la función en las áreas de mayor concentración de los puntos, además, los polinomios de grados elevados son de difícil interpretación biológica (22).
Son múltiples las funciones que han sido usadas para representar los cambios en las medidas de los pesos con respecto a la edad, las funciones más usadas son: ]]>
Donde: "Y" representa el peso; “β0 ” es el peso asintótico cuando "t" tiende a infinito, es interpretado como el peso adulto; “β1” es un parámetro de ajuste cuando “Y ≠0 ” o “t ≠0 ”; “β2” es un índice de madurez expresado como una proporción de porcentaje del máximo crecimiento con respecto al peso adulto del animal, y el parámetro “m” en la función de Richads se refiere al punto de inflexión (21).
Se han hecho estudios comparativos con los modelos matemáticos para describir las relaciones peso-edad en bovinos (4, 5, 7); estos autores observaron que los modelos de Von Bertalanffy, Gompertz y logístico sobrestiman los pesos a edades tempranas, mientras que el logístico subestima el peso a la madurez. El modelo de cuatro parámetros de Richards tiene un ajuste más preciso y requiere más cálculos para su ajuste que los modelos de tres parámetros, mientras que el modelo de Brody tiene un buen ajuste cuando se aplica a datos de bovinos con edades superiores a seis meses, ya que durante los primeros días de vida subestima el peso (30). ]]>
Los modelos de crecimiento deben poseer características para poder ser usados en los sistemas de producción: tener un punto de inflexión, presentar una asíntota horizontal, tener un comportamiento lógico y no permitir valores anormales desde el punto de vista biológico; tener una base biológica, es decir, que la ecuación debe derivarse del conocimiento teórico que se tenga de la variable dependiente que se analiza, lo que permite que al incorporar esta base teórica en el desarrollo de la ecuación se obtengan resultados más exactos (16). Además, otra característica deseable es la flexibilidad, entendida como la capacidad de ajuste con precisión a diferentes conjuntos de datos.
Los modelos no lineales cuando no son elegidos en forma empírica y se incorporan en ellos información sobre los procesos físicos o biológicos, como valores de las asíntotas, permiten una mejor interpretación y asociación de los parámetros obtenidos a procesos o fenómenos físicos. Estos modelos usan menos parámetros que los modelos lineales permitiendo obtener predicciones más fidedignas con mayor facilidad de interpretación de los parámetros que determinan la variable respuesta (29).
En los últimos años se ha implementado el uso de modelos mixtos, los cuales permiten analizar tanto efectos fijos como aleatorios y relacionarlos con la variable respuesta. Estos modelos pueden tomar varias formas, pero la más común es la que indica una distribución condicionada para la variable respuesta dada por los efectos aleatorios. Los primeros reportes que se tienen sobre la teoría y aplicación de los modelos mixtos fueron realizados por Pinheiro y Bates en 1995, generando una buena sustentación teórica sobre su aplicación (29).
La implementación de modelos mixtos brinda la posibilidad de analizar datos con estructuras de dependencia, no balanceados y en ocasiones con falta de normalidad y modelar, de manera flexible, complicadas estructuras de datos (6).
Los modelos mixtos incorporan tanto los parámetros fijos que son asociados a la población y los parámetros aleatorios, que están relacionados con los datos de las unidades experimentales (29). Estos modelos se han empleado para describir las relaciones entre las variables respuesta y algunas covariables que son agrupadas de acuerdo con la estructura de los datos. El número de parámetros aleatorios que se puede analizar en cada modelo puede ser solamente uno o tantos como parámetros fijos tenga el modelo, ya que a cada uno de estos se le puede asociar un efecto aleatorio. ]]>
A modo de ejemplo se presenta el modelo de Gompertz (12): , este mismo modelo con tres efectos aleatorios asociados a cada parámetro fijo, queda convertido en un modelo mixto que se expresa de la siguiente forma:
Donde, β0, β1 y β2 son coeficientes de la función (efectos fijos) que definen los valores esperados de las observaciones y b0i,b1iy b2i son los efectos aleatorios que definen la varianza y la covarianza de las observaciones para cada efecto fijo del i-ésimo animal, y εij representa el residuo asociado i-ésimo animal en el j-ésimo tiempo.
El análisis de medidas repetidas de un mismo individuo a través del tiempo implica una dependencia entre las observaciones tomadas en el individuo, esta situación hace que dichas observaciones estén correlacionadas. Por tanto, el análisis estadístico de estas debe tener en cuenta la variación entre el mismo individuo. Una ventaja de los modelos mixtos es que permiten una aproximación flexible para el modelamiento de los errores correlacionados mediante un modelo de covarianza que registre de forma adecuada la variación dentro de los animales (10, 33, 36).
Tanto el residuo como los efectos aleatorios se asumen como independientes, con distribución normal, media cero y varianza constante: ]]>
Además, la incorporación de efectos aleatorios en los modelos permite contemplar la variabilidad de las diferentes curvas entre los individuos de una misma población (6, 9).
Comparación de modelos
Al aplicar varios modelos de ajuste a una misma base de datos, es necesario decidir cual de todos es el que mejor ajusta y que, además pueda ser empleado para pronosticar resultados en los sistemas de producción. En este sentido, existe una serie de pruebas que pueden aplicarse a los modelos y ser usadas como herramienta de apoyo para decidir cual modelo es el que mejor ajusta. ]]>
Algunas pruebas son: la función de auto correlación, la cual se aplica a los residuos, se pueden analizar los valores estimados y sus varianzas; el coeficiente de determinación; la prueba de lack of fit (bondad de ajuste) que tiene la característica de separar la suma de cuadrados del residuo (SCR) en dos partes; la suma de cuadrados atribuida a la bondad de ajuste (SSF); y la suma de cuadrados atribuida al error puro (SSE). Otra herramienta usada para la selección de modelos es el porcentaje de curvas en que los parámetros de las funciones son significativos y el porcentaje de curvas atípicas para cada función (19).
Otra metodología usada para comparar modelos que evalúan una misma base de datos es la función de máxima verosimilitud (likelihood), que usualmente es empleada para estimar los coeficientes de un modelo matemático de regresión, en el que se calcula la probabilidad de que ocurra un suceso determinado.
Para poder aplicar la función de máxima verosimilitud los datos deben ser tomados en forma aleatoria, e independientes ente sí, además, que la probabilidad que un individuo muestre un suceso o un evento determinado (para nuestro caso que tenga un peso cualquiera) sea independiente de la probabilidad que otro individuo tenga el mismo u otro resultado; por lo que la probabilidad conjunta se calcula como el producto de las probabilidades individuales y de esa forma se obtiene la función de verosimilitud, que tiene en cuenta todos los datos de forma global, y será función únicamente de los coeficientes. De igual manera, se calcula la derivada de la función, se iguala a cero y se obtienen los valores de los coeficientes que maximizan ésa función (20).
Al combinar observaciones independientes en el cálculo de la función de verosimilitud interviene el producto de las probabilidades individuales, por lo que se utiliza la función logarítmica, ya que esta transforma los productos en sumas y los cocientes en restas (20).
En las salidas de los programas de análisis estadístico aparece el término Log−likehood o loglik, que es el logaritmo de la verosimilitud. Al tratarse de productos de probabilidades la función de verosimilitud será siempre menor que 1 y por tanto su logaritmo es negativo, en la práctica para eliminar el signo negativo del loglik se multiplica por -2. ]]>
La función de verosimilitud permite comparar modelos que incluso tengan diferente número de parámetros, por ejemplo si a un modelo se le adiciona una variable adicional con respecto al otro modelo.
Las diferencias en la función de verosimilitud se alteran arbitrariamente con la escala de medida, por lo que la forma adecuada de compararlas es mediante cocientes. Es por eso que cuando se comparan modelos que han sido estimados mediante este procedimiento se hable de razón de verosimilitud (likelihood ratio) (20).
Cuando se trata de la estimación de modelos resulta de utilidad el concepto de modelo saturado. Un modelo se denomina saturado cuando utiliza tantos parámetros como observaciones se hayan efectuado y por tanto se ajusta perfectamente a los datos, pudiendo comparar el modelo actualmente estimado con el modelo teórico perfecto o modelo saturado mediante la expresión:
, o se pueden comparar modelos cuando en uno de ellos se incluye una variable adicional:
, si en el modelo 2 se adiciona una variable más que en el modelo 1, y si el contraste resulta ser no significativo, se asume que la incorporación de la variable en el modelo 2 no mejora sensiblemente la verosimilitud del modelo y por tanto no merece ser tenida en cuenta (20). ]]>
En las salidas de los programas suele aparecer el término likelihood ratio o cociente de verosimilitud para un modelo, sin que se especifique que se esté contrastando con otro diferente. En estos casos el contraste es frente al modelo que sólo incluye el término constante y por tanto no se consideran las variables o los factores de riesgo, y se compara con el modelo que sí incluye las variables, por lo que ahora esa cantidad se distribuye según una χ 2 con grados de libertad igual al número de variables incluidas en el modelo, que es la diferencia frente al modelo con sólo la constante. Al igual que antes, si el contraste resulta no significativo quiere decir que incluir el conocimiento de las variables no mejora significativamente la verosimilitud del modelo y por lo tanto se trata de un modelo sin utilidad (20).
Cada que se adicionan más variables a un modelo la función de verosimilitud mejorará y si la muestra es grande es difícil distinguir mediante el contraste del cociente de verosimilitud entre una mejora “real” y un aporte insignificante. Ya que el modelo perfecto no existe, puesto que todos constituyen simplificaciones de la realidad siempre es preferible trabajar con modelos que tengan menos variables, porque además de ser más sencillos, son más estables y presentan menos sesgo. Por ello se han propuesto otras medidas de contraste entre modelos, que penalizan en alguna medida el hecho de tener más o menos parámetros. Es el caso del criterio de información Akaike (AIC) y el criterio de información Bayesiano (BIC) que incluyen en su fórmula el loglik. (10).
Donde npar es el número de parámetros del modelo y N es el número total de observaciones que se usaron en el modelo. Bajo los conceptos planteados anteriormente es mejor el criterio que presente el menor valor. Esto es, si se usa el BIC para comparar dos o más modelos con los mismos datos, se debe preferir el modelo que presente un menor BIC (29). ]]>
Parámetros para evaluar el crecimiento
Los parámetros que pueden ser empleados para evaluar o describir el crecimiento se describen a continuación:
1. y = y() t . Donde “ y ” es el peso o variable dependiente y “t ” es el tiempo o variable independiente. Esta ecuación es la representación de una o más medidas en el tiempo.
2. dy/dt: La velocidad del crecimiento tiene gran importancia económica, ya que podría ser usada para fijar el momento de sacrificio y poder realizar el aprovechamiento del animal cuando haya alcanzado la máxima velocidad de crecimiento. Después de este momento la ganancia de peso es más lenta, sinembargo, por razones comerciales el sacrificio debe hacerse en otros momentos buscando características especiales en la canal. La información que suministra se limita a la tasa de crecimiento de un momento dado (17). ]]>
3. El punto de inflexión es otro parámetro usado para evaluar el crecimiento, se localiza en los sitios en que se presentan cambios de incremento o decrecimiento del peso con respecto a la edad. La máxima velocidad de crecimiento coincide con uno de los puntos de inflexión que se pueden hallar en las curvas de crecimiento (16, 17).
4. dy2/dt2: La aceleración del crecimiento que indica la tasa absoluta de la ganancia de peso con respecto al tiempo. Cuando el resultado de esta ecuación es cero se dice que allí se localiza el punto de inflexión (18).
5.(1/y) ( dy/dt): La tasa relativa de crecimiento, también llamada tasa específica de crecimiento o velocidad relativa de crecimiento, corrige los efectos de escala debidos al formato del animal. Es útil para comparar líneas o especies. Proporciona datos que no dependen de la dimensión. Indica la variación del crecimiento cuando este se expresa en coordenadas logarítmicas. Decrece con la edad.
6. ,Donde βo es el peso adulto cuando el tiempo "t" tiende a infinito. El peso adulto es un parámetro un poco más difícil de definir, no sólo porque se alcanza a una edad tardía desde el punto de vista comercial, sino también por depender de la cantidad de tejido graso, que es muy variable a lo largo del estado adulto del animal. En la práctica es complicado estimar el peso adulto ya que las hembras varían de peso según el tamaño, el número de partos, y porque la aproximación asintótica al peso adulto final es muy lenta, por lo que no es frecuente encontrar animales en las explotaciones que lo hayan alcanzado. En la práctica el “peso casi-adulto” es suficiente para la mayor parte de usos de este parámetro (2).
7. Otro parámetro que se tiene en cuenta en las explotaciones pecuarias es la tasa de madurez sexual propuesto por Taylor en 1971 (34) con respecto al peso adulto (1, 17). ]]>
8. Donde "v" es el grado de madurez. Esta ecuación hace referencia al grado de madurez absoluta.
El momento óptimo de sacrificio se ha intentado definir, procurando que se produzca antes de que la velocidad de crecimiento descienda demasiado, con intentos de situarlo cerca del punto de inflexión (donde se logra la máxima velocidad de crecimiento). Sin embargo, el momento de sacrificio está determinado por el mercado y no es fácil modificarlo (2).
Una vez que se ha determinado el o los modelos que mejor ajustan se debe analizar los parámetros de crecimiento antes descritos, para determinar el desempeño productivo de los animales, poder realizar ajustes de producción, de alimentación y de programación general del hato, pues se puede calcular la edad a la que alcanzan el punto de inflexión o la edad a la que alcanzan la mayor velocidad de crecimiento, la edad a la que alcanzan el peso adulto y el grado de madurez en los diferentes momentos de la vida del animal.
Procedimientos para el análisis de medidas repetidas (crecimiento animal) ]]>
A continuación se ilustran varios procedimientos para analizar una base de datos que contiene información sobre crecimiento animal, cuyo objetivo es estimar los parámetros y comparar los resultados obtenidos en las diferentes metodologías empleadas. Los datos son procesados con el programa estadístico SAS (32).
Modelación de los datos
Utilizando el modelo de Brody se analiza la información simulada de 500 animales, se tomó como referencia los valores de los parámetros encontrados por Fonseca y Aquino 2001 (8), en ganado Nelore (hembras y machos). Los valores de los parámetros encontrados por los autores se pueden observar en la tabla 1.
]]>
Para desarrollar la modelación es necesario simular una base de datos que contenga información sobre el peso de los animales.
En la programación β0 se define como sem0, β1 como sem1 y correspondiente a sem2 , además se tuvo en cuenta un tiempo (t) total de 2160 días con intervalos de 90 días entre pesajes y Y es la variable respuesta expresada en kg; a continuación se muestra la programación para crear la base de datos con la información de los animales que fue llamada grupo1.
El comando rannor permite generar los valores aleatorios para sem0, sem1 y sem2, con media cero y varianza uno, los cuales son multiplicados por el valor del desvío y se suma la media del parámetro. El valor entre paréntesis después del comando rannor, indica el valor de la semilla de arranque Ej: rannor (5). ]]>
Si se desea observar la base de datos con la información de los animales se debe ejecutar la siguiente programación:
proc print data=grupo1; run
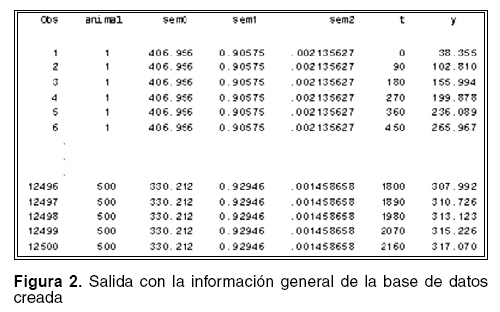
En la figura 2 se puede observar la salida en la que se identifica la siguiente información: Obs (es el número de cada pesaje, 1,2,3,……,12500), animal (número de cada individuo, 1,2,3,….,500), sem0, sem1, sem2 (son los valores aleatorios que se generan para cada parámetro), t (es la edad o el tiempo al que se realiza el pesaje a cada animal, 0,90,180,…….,2160), Y es el peso en kg. que se ha observado en cada observación para cada animal a un tiempo determinado, aplicando la curva de Brody.
]]>
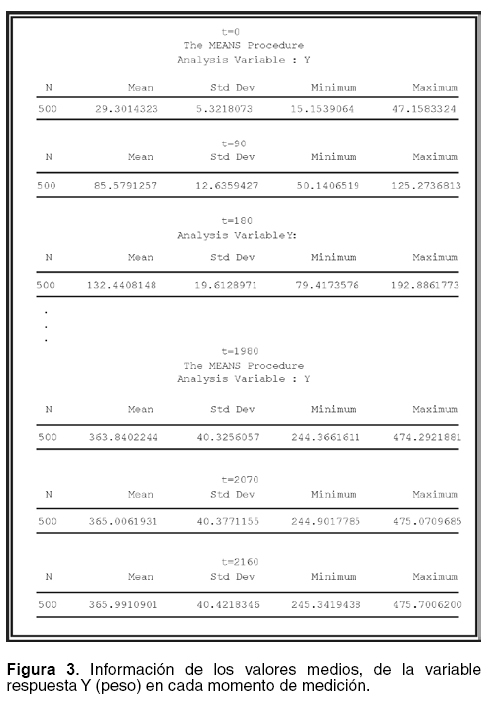
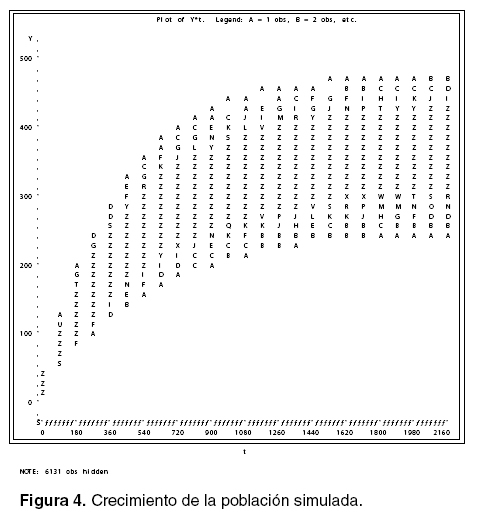
Para realizar la caracterización de la población y las curvas de crecimiento para cada individuo para obtener los valores medios, la desviación estándar y los valores mínimos y máximos de la variable respuesta en un momento determinado, además para observar la figura de crecimiento de los individuos, se realizó la siguiente programación, las salidas se pueden ver en las figuras 3 y 4.
]]>
En la figura 4, se puede observar el aumento de peso de los animales e identificar claramente que con el aumento de la edad aumenta la variación de los pesos, así como la tendencia de los animales a alcanzar la asíntota (cuando llegan al peso adulto).
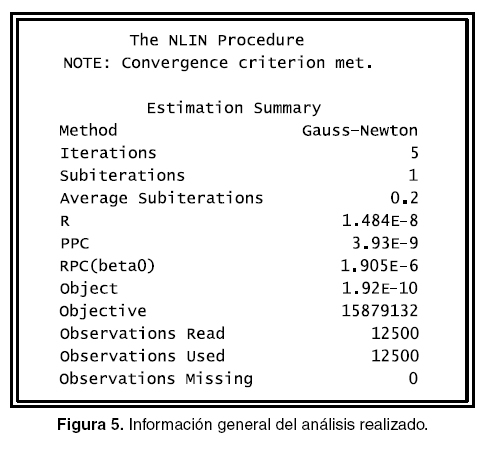
Inicialmente los parámetros de la función son estimados por mínimos cuadrados, este procedimiento se realiza con el comando proc nlin que es usado en regresiones no lineales, el procedimiento primero requiere los valores iniciales o de arranque, para luego evaluar los residuos y la suma de cuadrados para cada combinación de los valores y así determinar los mejores valores para realizar la interacción algorítmica. Para realizar lo anterior existen cuatro métodos: Gauss-Newton, Marquardt, método del gradiente o steepest y secante multivariada o posición falsa. Si en la programación no se específica una de las anteriores metodologías el programa por defecto aplica Gauss-Newton.
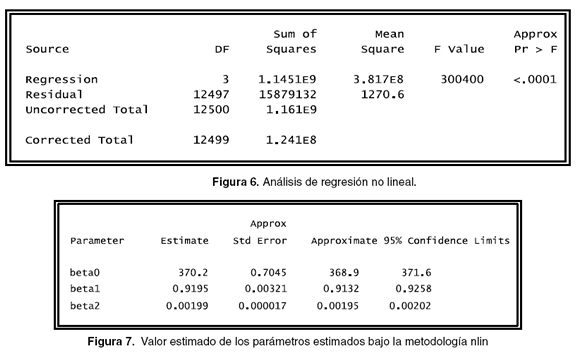
En la programación es necesario incluir un rango en el que se puede ubicar el parámetro, en este caso son: beta0 = 220 to 550 para el peso adulto, beta1 = 0.7 to 0.98 para el factor de ajuste y beta2 = 0.001 to 0.0019 para el índice de precocidad, lo anterior se realiza aplicando el comando parms. Además, es necesario indicar el modelo de la regresión que se utilizará, en este caso es el modelo de Bordy, esta indicación se realiza con el comando model.
]]>
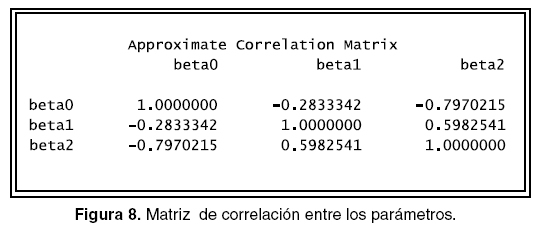
La salida puede observarse en la figuras 5, 6, 7 y 8, en las que se identifica lo siguiente:
El anterior procedimiento realiza el análisis a la población en general, sin embargo, no se tiene en cuenta la relación existente dentro de cada individuo, para ello se aplica el comando by que permite realizar la análisis por individuo, donde se obtendrán las estimativas de los parámetros por individuo. Con el procedimiento means se obtendrán las medias y la varianzas para cada parámetro en todos los individuos. La programación se muestra a continuación y la salida se observa en la figura 9.
]]>
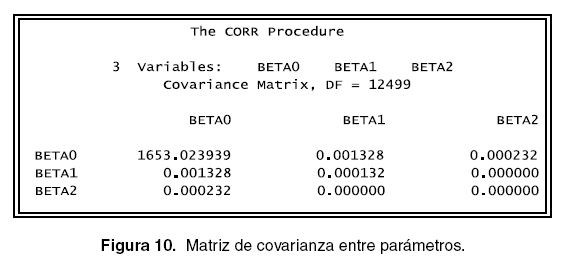
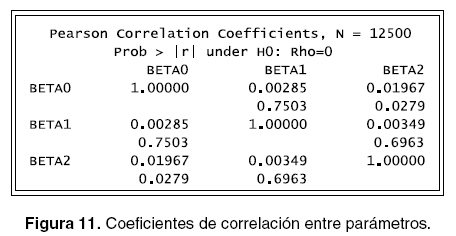
Otro aspecto importante que se debe revisar es si los parámetros están o no correlacionados pues se parte en la simulación de los datos que estos presentan una baja o nula correlación. El comando En la figura 12 se puede observar que no existe proc corr permite realizar el análisis numérico y el ningún tipo de correlación entre los parámetros. comando proc plot permite ver en forma gráfica, la programación que permite hacer lo anterior se Los anteriores procedimientos se realizaron con muestra a continuación y las salidas se pueden ver todos los parámetros fijos. A continuación se realiza en las figuras 10, 11 y 12. ]]>
En la figura 12 se puede observar que no existe ningún tipo de correlación entre los parámetros.
Los anteriores procedimientos se realizaron con todos los parámetros fijos. A continuación se realiza el análisis de la misma base de datos aplicando el modelo mixto, incluyéndose a un sólo parámetro β0 el componente aleatorio b0 con
]]>
El modelo es donderepresenta la j-ésima medida (j=1,…,25) del i-ésimo individuo (i=1,/serial/rccp/v20n2.,500), β1 es un parámetro de ajuste, β2 es un parámetro relacionado a la precocidad o índice de madurez del animal, tij es el tiempo medido en días (tomado desde el nacimiento tij =0 hasta tij = 2160, con mediciones cada 90 días) y εij es el error residual con y considerado independiente de b0. Los efectos fijos del modelo son: β0, β1 y β2. ]]>
Para la estimación de los parámetros del modelo bajo la metodología de modelos mixtos se puede utilizar el procedimiento nlmixed. La programación con un efecto aleatorio es la siguiente:
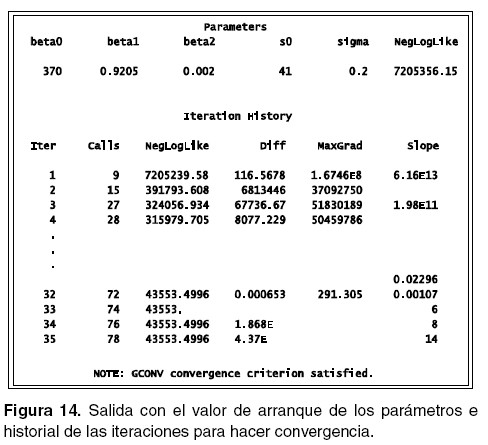
El comando data permite llamar la base de que se ha simulado con la información de los animales del grupo1; gconv se emplea para especificar el criterio que hace referencia al gradiente de convergencia; parms identifica los parámetros que se desean calcular e igualmente permite adjudicarle los valores iniciales o de arranque (beta0= 370 beta1=0.9205 beta2=0.0020 s0=41 y sigma=0.2); bounds se emplea para condicionar a los parámetros a que estén dentro de un rango de valores determinado o como en el caso se la varianza del error s0 y su desvío sigma a que sean positivos.
El modelo que se desea analizar es indicado después del comando model, y la orden random permite definir el efecto aleatorio, que en este caso es b0 asociado al peso adulto y especificar que este tiene una distribución normal, con media 0 y varianza s0**2, subject=animal permite asegurar que se genere un efecto aleatorio asociado a cada medición de los animales. ]]>
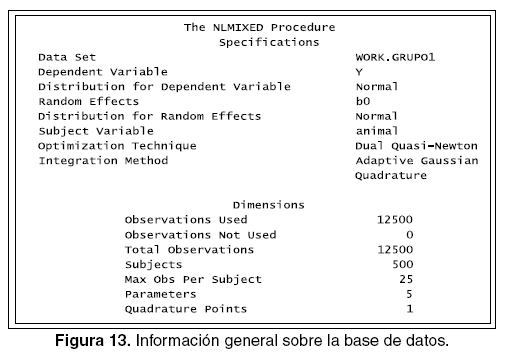
En la figura 13 se observa la lista del modelo mixto que ha sido especificada, incluye la base de datos, la variable dependiente, el efecto aleatorio, las distribuciones como las técnicas de optimización; estas últimas no es necesario programarlas, además, se puede observar las dimensiones de la base de datos: cantidad total y por individuo de observaciones realizadas y cuánta de esta no ha sido usada. Esta salida sirve para chequear que los datos especificados y que el modelo está corriendo apropiadamente.
En la figura 14 se observa la lista de parámetros con los respectivos valores de arranque, así como el Negloglike que evalúa el modelo. Además se puede observar el número de iteraciones que debió realizar el sistema para encontrar la convergencia de los parámetros.
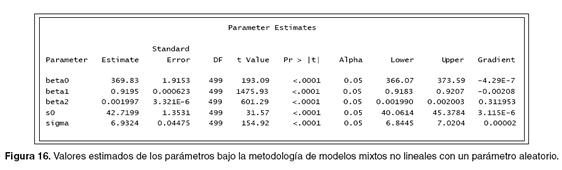
Además, se obtiene el listado de criterios que sirven para evaluar el modelo o compararlo con otros (véase Figura 15), estos valores luego serán tenidos en cuenta para ser comparados con los valores obtenidos cuando la base de datos sea procesada con dos parámetros aleatorios.
]]>
Igualmente, se observan los valores estimados para cada uno de los parámetros y de sus respectivos errores estándar, además se ven los grados de libertad que es igual al número de individuos menos el número de parámetros aleatorios, en la última columna se puede observar el gradiente de optimización, indicando una cifra lo suficientemente pequeña que puede ser interpretada como un punto inmóvil para cada parámetro (véase Figura 16). Al comparar la información obtenida con la información de la figura 7, se puede ver que se obtiene más información (lower, upper), que puede ser utilizada para realizar un mejor análisis sobre la base de datos, además se obtienen los valores de los parámetros (s0 y sigma).
]]>
El mismo modelo de Brody con dos parámetros aleatorios b0 y b2 asociados el primero al peso adulto β0, y el otro al índice de precocidad β2, da lugar al modelo mixto: Al igual que en el caso anterior, debe aplicarse el procedimiento nlmixed, indicar el valor de los parámetros beta0, beta1 y beta2, así como el de “s0” y “s2” que representan las varianzas de β0 y de β2 respectivamente y “sigma” que es el desvío del error, aclarando en la programación que “s0”, “s2” y“sigma” deben ser positivos y que b0 y b2 tienen una distribución normal, con media cero y varianzas s0**2ys2**2, respectivamente y covarianza cero. Se eligió trabajar con éstos dos parámetros aleatorios por que son los que presentan mayor facilidad de interpretación biológica aplicados al crecimiento animal. La programación es la siguiente:
]]>
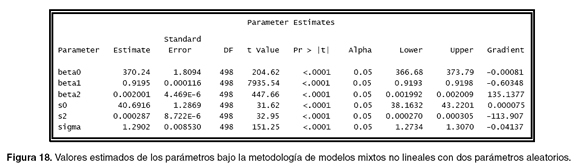
Igual que en la programación del modelo mixto con un parámetro aleatorio, el programa genera la información general que hace referencia a las dimensiones, especificaciones del modelo, el valor de los parámetros de arranque y el histórico de las iteraciones realizadas antes de hacer convergencia. En la figura 17 se identifican los métodos de evaluación del modelo, pudiendo ver que el valor del BIC es mucho menor que el valor calculado en el modelo de un sólo parámetro aleatorio (véase Figura 15), lo que indica que el modelo mixto con dos parámetros aleatorios es un mejor modelo.
En la figura 18, se ve claramente que los valores estimados de los parámetros son similares a los valores que se simularon en la base de datos grupo1, lo que indica que se está realizando un buen ajuste y otorga además, los valores calculados para los parámetros “beta0, beta1 y beta2”; 370.24 , 0.9195 y 0.002001, respectivamente y las varianzas “s0” = 40.6916 y“s2” = 0.000287 asociadas al primer y tercer parámetro.
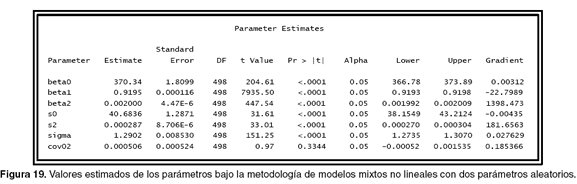
La siguiente programación se realizó para incluir la relación existente entre los dos parámetros (cov02=0.000009) que es el valor inicial de la covarianza entre los dos parámetros.
]]>
En la figura 19 se puede ver que la covarianza entre los dos parámetros fue de 0.000506.
La base de datos grupo1 también podría ser analizada con tres parámetros aleatorios. Sin embargo, consideramos que es suficiente el análisis con los dos parámetros aleatorios que tienen una interpretación biológica. Además con tres o más parámetros aleatorios se presentan dificultades computacionales que dificultan el procesamiento de los datos. ]]>
Consideraciones finales
El avance en las metodologías estadísticas y la mejora de las herramientas computacionales han permitido una mejora sustancial en los sistemas análisis de datos en las empresas pecuarias, pudiendo estas informaciones ser empleadas para realizar proyecciones y programaciones en los sistemas de producción.
El uso de modelos mixtos es una metodología rigurosa, al momento de analizar datos que estén correlacionados, pero su implementación requiere que los datos a analizar tengan una estructura de medidas repetidas. Su uso en los últimos años ha abarcado áreas del conocimiento como lo son la medicina (farmacocinética de droga), la producción agropecuaria (curvas de crecimiento en plantas, curvas de crecimiento animal, curvas de producción láctea y curvas de respuesta fotosintética), entre otros, lo que hace pensar que esta metodología tiene una amplia gama de posibilidades en los sistemas de producción animal, con esta metodología se tienen sistemas para comparar modelos que son más estrictos como el AIC y BIC que son más rigurosos que las metodologías tradicionales (28).
Referencias ]]>
1. Beltran JJ, Butts WT, Oson TA, Koger M, et al. Growth patterns of two lines of angus cattle selected using predicted growth parameters. J Anim Sci 1992; 70:734-741. [ Links ]
2. Blasco A. XIV Curso internacional sobre mejora genética animal. Universidad Politécnica de Valencia; 2004. 21 p. [ Links ]
3. Brody S. Bioenergetics and growth. Reinhold publishing corporation. New York. 1945. 1023 p. [ Links ]
4. Brown J, Fitzhugh, H, Cartwright T. Comparison of nonlinear models for describing weight-age relationships in cattle. J Anim Sci 1976; 42:810-818. [ Links ]
5. Denise RS, Brinks JS. Genetic and environmental aspects of the growth curve parameters in beef cows. J Anim Sci 1985; 61:1431-1440. [ Links ]
6. El Halimi R. Nonlinear mixed-effects models and nonparametric inference. Tesis de doctorado, Departamento de estadística, Universidad de Barcelona, Barcelona, 2005. 246 p. [ Links ]
7. Fittzhugh JH. Analysis of growth curves and strategies for altering their shape. J Anim Sci 1976; 42:1036-1051. [ Links ]
8. Fonseca F, Aquino LH, Gomes AI. Estimativas de parâmetros genéticos de curva de crescimento de gado nelore (Bos indicus). Cienc Agrotec 2002; Suppl 1562-1567. [ Links ]
9. France J, Dijkstra J, Dhanoa M. Growth functions and their application in animal sciences. Ann Zootech 1996; 45:165-174. [ Links ]
10. Gaona B. Matrices de covarianza estructuradas en modelos con medidas repericas. Tesis de maestría, Universidad de Puerto Rico, Mayagüez, 2005. 127 p. [ Links ]
11. Gómez LH. Estadística experimental aplicada a las ciencias Agrícolas. Universidad Nacional de Colombia. Sede Medellín; 1997. 571 p. [ Links ]
12. Gompertz B. On the nature of the function expressive of the law of human mortality, and on a new Mode of determining the value of life Contingencies. Philosophical Transactions of the Royal Society of London 1825; 115:513-585. [ Links ]
13. Hammond J. Avances en fisiología zootécnica: Zaragoza. Acribia; 1959. 1330 p. [ Links ]
14. Helman MB. Cebutecnia. 2nd ed. Buenos Aires: El ateneo; 1984. 538 p. [ Links ]
15. Joander G, Cartwright T. Estimation of efficiency of beef production. J Anim Sci 1969; 29:862-868. [ Links ]
16. Kiviste A, Álvarez G, Rojo A, Ruiz G. Funciones de crecimiento de aplicación en el ámbito forestal. Madrid: Instituto nacional de Investigación y Tecnología Agraria y Alimentaria; 2002. [ Links ]
17. Kratochvilova M, Hyanková L, Knizetová H, Fólder F. Growth curve análisis in cattle from early maturity and mature body size viewpoints. Czech J Anim Sci 2002; 47:125-132. [ Links ]
18. López SJ. France EJ, Gerrits MS, Dhanoa DJ, Humphries, et al. Ageneralized michaelis-menten equation for the analysis of growth. J Anim Sci 2000; 78:1816-1828. [ Links ]
19. Ministry of forests research program. Biometrics information. British Columbia: The institute; 1990. [ Links ]
21. Moore AJ. A mathematical equation for animal growth from embryo to adult. Anim Prod 1985; 40:441-453. [ Links ]
22. Muñoz BM, Tonhihati H. Uso de polinomios segmentados para el estudio de la curva de lactación en búfalos murrah y sus mestizos. Asociación peruana de producción animal 2002; [noviembre, 2005] URL: http://www.appaperu.org/appa2002/invpdf/prod/71Prosocie.pdf [ Links ]
23. Nobre PR, Misztal S, Tsuruta and Bertrand JK. Analysis of growth curves of nellore cattle by multiple-trait and random regression models. J Anim Sci 2003; 81:918-926. [ Links ]
24. Nobre PR, Rosa AN, Euclides FK. Interações reprodutor x estação de nascimento e reprodutor x fazenda sobre o crescimento de bezerros Nelore. Rev Bras Zootec 1988; 17:120-181. [ Links ]
25. Oliveira HN, Lobo RB, Pereira CS. Relationships among growth curve parameters, weights and reproductive traits in guzera beef cows. World congress on genetics applied to livestock production, Guelph; 1994. [ Links ]
26. Oliveira JA, Alemcar MM, Llima R. Causas de variação não genética sobre características de peso de animais de um rebanho nelore. Reunião anual da sociedade brasileira de zootecnia; 1994. 157 p. [ Links ]
27. Parks JA. Theory of feeding and growth of animals. Springger- Verlag, Berlin; 1982. 451 p. [ Links ]
28. Peek SM, Cohen ER, Wait DA, ForsethIn. Physiological response curve analysisi using nonlinear mixed models. Oecologia 2002; 132:175-180. [ Links ]
29. Pinheiro JC, Bates D. Mixed effects models in S and S-PLUS. Springer-Verlag, Berlin; 2000. 528 p. [ Links ]
30. Pereda ME, González SS, Arjona E, Bueno G, Mendoza GD. Ajuste de modelos de crecimiento y cálculo de requerimientos nutricionales para bovinos Brahman en Tamaulipas, Mexico. Agrotecnia 2005; 39:19-27. [ Links ]
31. Richards FJ. A flexible growth function for empirical use. J Exp Bot 1959; 10:290-300. [ Links ]
32. Sherchand L, McNew RW, Kellogg DW, Jonson ZB. Selection of mathematical model to generate lactation curves using daily milk yields of Holstein cows. J Dairy Sci 1995; 78:2507-2513. [ Links ]
33. Statistical Analysis Systems. SAS®, versión 9.1 para Windows,. User´s Guide. Statistics. Statistical Analysis Systems Institute. Inc., Cary, North Carolina; 2005. [ Links ]
34. Taylor St CS. A relation between mature weight and time taken to mature in mammals. Anim Prod 1965; 7:203-220. [ Links ]
35. Verhulst PF. Notice sur la loi que la population pursuit dans son accroissement. Corresp Math Phy 1938; 10:113-121. [ Links ]
36. Von Bertalanffy L. A quantitative theory of organic growth. Hum Biol 1938; 10:181-213. [ Links ] ]]>199270734-741200419451023197642810-8181985611431-14402461976421036-105120021562-1567199645165-17412719975711825115513-5851959133019842538196929862-8682002200247125-1322000781816-1828Ministry of forests research program1990marzo, 2198540441-4532002200381918-926198817120-1811994Guelph 199415719824512002132175-180200052820053919-27195910290-3001995782507-2513Statistical Analysis Systems Institute. Inc.200519657203-220193810113-121193810181-213
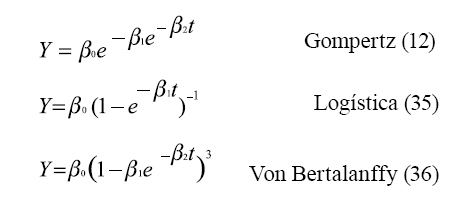
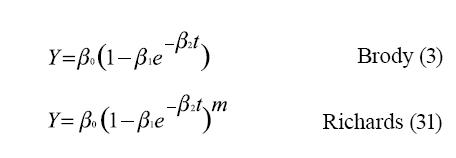
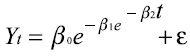 , este mismo modelo con tres efectos aleatorios asociados a cada parámetro fijo, queda convertido en un modelo mixto que se expresa de la siguiente forma:
, este mismo modelo con tres efectos aleatorios asociados a cada parámetro fijo, queda convertido en un modelo mixto que se expresa de la siguiente forma: 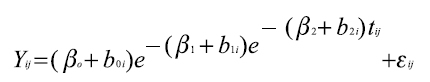 Donde, β0, β1 y β2 son coeficientes de la función (efectos fijos) que definen los valores esperados de las observaciones y b0i,b1i y b2i son los efectos aleatorios que definen la varianza y la covarianza de las observaciones para cada efecto fijo del i-ésimo animal, y εij representa el residuo asociado i-ésimo animal en el j-ésimo tiempo.
Donde, β0, β1 y β2 son coeficientes de la función (efectos fijos) que definen los valores esperados de las observaciones y b0i,b1i y b2i son los efectos aleatorios que definen la varianza y la covarianza de las observaciones para cada efecto fijo del i-ésimo animal, y εij representa el residuo asociado i-ésimo animal en el j-ésimo tiempo. 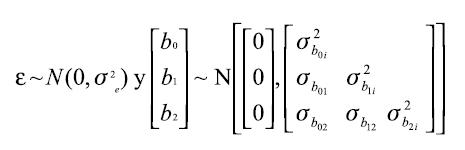
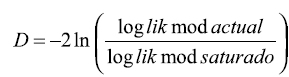 , o se pueden comparar modelos cuando en uno de ellos se incluye una variable adicional:
, o se pueden comparar modelos cuando en uno de ellos se incluye una variable adicional: 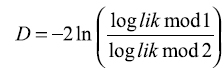 , si en el modelo 2 se adiciona una variable más que en el modelo 1, y si el contraste resulta ser no significativo, se asume que la incorporación de la variable en el modelo 2 no mejora sensiblemente la verosimilitud del modelo y por tanto no merece ser tenida en cuenta (20). ]]>
, si en el modelo 2 se adiciona una variable más que en el modelo 1, y si el contraste resulta ser no significativo, se asume que la incorporación de la variable en el modelo 2 no mejora sensiblemente la verosimilitud del modelo y por tanto no merece ser tenida en cuenta (20). ]]>
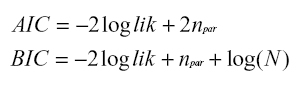
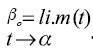 ,Donde βo es el peso adulto cuando el tiempo "t" tiende a infinito. El peso adulto es un parámetro un poco más difícil de definir, no sólo porque se alcanza a una edad tardía desde el punto de vista comercial, sino también por depender de la cantidad de tejido graso, que es muy variable a lo largo del estado adulto del animal. En la práctica es complicado estimar el peso adulto ya que las hembras varían de peso según el tamaño, el número de partos, y porque la aproximación asintótica al peso adulto final es muy lenta, por lo que no es frecuente encontrar animales en las explotaciones que lo hayan alcanzado. En la práctica el “peso casi-adulto” es suficiente para la mayor parte de usos de este parámetro (2).
,Donde βo es el peso adulto cuando el tiempo "t" tiende a infinito. El peso adulto es un parámetro un poco más difícil de definir, no sólo porque se alcanza a una edad tardía desde el punto de vista comercial, sino también por depender de la cantidad de tejido graso, que es muy variable a lo largo del estado adulto del animal. En la práctica es complicado estimar el peso adulto ya que las hembras varían de peso según el tamaño, el número de partos, y porque la aproximación asintótica al peso adulto final es muy lenta, por lo que no es frecuente encontrar animales en las explotaciones que lo hayan alcanzado. En la práctica el “peso casi-adulto” es suficiente para la mayor parte de usos de este parámetro (2). 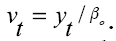 Otro parámetro que se tiene en cuenta en las explotaciones pecuarias es la tasa de madurez sexual propuesto por Taylor en 1971 (34) con respecto al peso adulto (1, 17).
Otro parámetro que se tiene en cuenta en las explotaciones pecuarias es la tasa de madurez sexual propuesto por Taylor en 1971 (34) con respecto al peso adulto (1, 17). 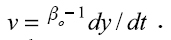 Donde "v" es el grado de madurez. Esta ecuación hace referencia al grado de madurez absoluta.
Donde "v" es el grado de madurez. Esta ecuación hace referencia al grado de madurez absoluta. 





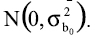
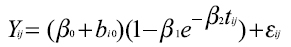 representa la j-ésima medida (j=1,…,25) del i-ésimo individuo (i=1,/serial/rccp/v20n2.,500), β1 es un parámetro de ajuste, β2 es un parámetro relacionado a la precocidad o índice de madurez del animal, tij es el tiempo medido en días (tomado desde el nacimiento tij =0 hasta tij = 2160, con mediciones cada 90 días) y εij es el error residual con y considerado independiente de b0. Los efectos fijos del modelo son: β0, β1 y β2. ]]>
representa la j-ésima medida (j=1,…,25) del i-ésimo individuo (i=1,/serial/rccp/v20n2.,500), β1 es un parámetro de ajuste, β2 es un parámetro relacionado a la precocidad o índice de madurez del animal, tij es el tiempo medido en días (tomado desde el nacimiento tij =0 hasta tij = 2160, con mediciones cada 90 días) y εij es el error residual con y considerado independiente de b0. Los efectos fijos del modelo son: β0, β1 y β2. ]]>


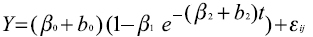 Al igual que en el caso anterior, debe aplicarse el procedimiento nlmixed, indicar el valor de los parámetros beta0, beta1 y beta2, así como el de “s0” y “s2” que representan las varianzas de β0 y de β2 respectivamente y “sigma” que es el desvío del error, aclarando en la programación que “s0”, “s2” y “sigma” deben ser positivos y que b0 y b2 tienen una distribución normal, con media cero y varianzas s0**2ys2**2, respectivamente y covarianza cero. Se eligió trabajar con éstos dos parámetros aleatorios por que son los que presentan mayor facilidad de interpretación biológica aplicados al crecimiento animal. La programación es la siguiente:
Al igual que en el caso anterior, debe aplicarse el procedimiento nlmixed, indicar el valor de los parámetros beta0, beta1 y beta2, así como el de “s0” y “s2” que representan las varianzas de β0 y de β2 respectivamente y “sigma” que es el desvío del error, aclarando en la programación que “s0”, “s2” y “sigma” deben ser positivos y que b0 y b2 tienen una distribución normal, con media cero y varianzas s0**2ys2**2, respectivamente y covarianza cero. Se eligió trabajar con éstos dos parámetros aleatorios por que son los que presentan mayor facilidad de interpretación biológica aplicados al crecimiento animal. La programación es la siguiente: