0.05), pero en mayor medida que la observada con rolipram 25 y 75 µM (31.25, y 28.61%, respectivamente). Los CCOs cultivados en presencia de rolipram 50 µM+Gonadotropinas maduraron en menor proporción (63.74%) comparada con gonadotropinas (p<0.01) o rolipram 50 µM (p<0.05). Los resultados permiten concluir que el porcentaje de maduración nuclear in vitro de oocitos bovinos depende de la dosis de rolipram utilizada, donde la concentración de rolipram 50 µM presentó un comportamiento similar a las gonadotropinas en la maduración del oocito. Además, la presencia del estímulo gonadotrópico y del inhibidor de PDE puede inducir una respuesta de desensibilización de la vía del AMPc, reflejada en una disminución del porcentaje de maduración.]]>
0.05) did, but in a higher way than rolipram 25 µM (31.25%) or 75 µM (28.61%). The COC cultured with rolipram 50 µM+gonadotrophins maturated in a lower proportion (63.74%) than did with gonadotropins (p<0.01) or rolipram 50 µM (p<0.05). A dose-dependent response of percentage of IVNM of bovine oocytes was detected. Thus rolipram 50 µM, exerts a similar effect of gonadotropins on oocyte maturation. In addition, the presence of gonadotropic stimulus and a PDE inhibitor may induce a desensitization response of the cAMP way, as suggested by the reduction of the percentage of maturation.]]>
0.05), porém em maior medida que a observada com rolipram 25 y 75 µM (31.25, y 28.61%, respectivamente). Os CCOs cultivados na presença de rolipram 50 µM+Gonadotrofinas maturaram em menor proporção (63.74%) quando comparado com gonadotrofinas (p<0.01) ou rolipram 50 µM (p<0.05). Os resultados permitem concluir que a percentagem de maturação nuclear in vitro de oocitos bovinos depende da doses de rolipram utilizada, sendo que a concentração de rolipram 50 µM apresentou um comportamento similar às gonadotrofinas na maturação do oocito. Ale disto, a presença do estímulo gonadotrópico e do inibidor de PDE podem induzir uma resposta de diminuição na sensibilização da via do AMPc, refletida numa diminuição da percentagem de maturação.]]>
Efecto del inhibidor de fosfodiesterasa tipo 4 —rolipram, sobre la maduración in vitro de oocitos bovinos¶
Effect of phosphodiesterase type 4-inhibitor rolipram on in vitro maturatrion of bovine oocytes
Efeito do inibidor de fosfodiesterasa tipo 4 —rolipram, sobre a maturação in vitro de oocitos bovinos
Yuddy S López1, Biol; Ana M Mejía1, Biol; Elder E Escobar4, Biol, MS; Bernardo Agudelo Jaramillo5, MD, GO; Neil A Vásquez Araque1,2*, Biol, MS; Hernán Echavarría3, Zoot, MS.
]]> 1Grupo de Biotecnología Animal, 2Facultad de Ciencias, y 3Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia sede Medellín. AA 3840, Medellín, Colombia.4Instituto Antioqueño de Reproduccion, INSER, Medellín, Colombia
5Grupo de Investigación GIE-SHAE, Facultad de Medicina, Universidad de Antioquia. AAA 1226, Medellín, Colombia.
(Recibido: 12 junio, 2007; aceptado: 21 febrero, 2008)
Resumen
Durante el proceso de maduración in vitro de oocitos, las gonadotropinas FSH y LH inducen la producción de AMPc. El AMPc tiene efecto dual, donde los altos niveles de AMPc intraoocitario mantienen su bloqueo meiótico, mientras que en las células de la granulosa inducen la maduración del oocito. Los niveles de AMPc son regulados por hidrólisis mediada por fosfodiesterasas (PDE), las cuales presentan expresión específica en el folículo, el oocito expresa la PDE 3, mientras que las células de la granulosa PDE 4. Con el objetivo de evaluar el efecto del rolipram, un inhibidor específico de PDE 4, sobre el porcentaje de maduración nuclear in vitro (MNIV) de oocitos bovinos, 629 complejos cúmulo oocito (CCO) fueron cultivados a 38.5 °C/5% CO2/24 h, en medio TCM-199 con la adición de pFSH y hrLH, o rolipram. Los grupos experimentales fueron: adición de gonadotropinas, rolipram (25, 50 ó 75 µM), rolipram 50 µM + gonadotropinas, o control sin estímulo. Los oocitos fueron teñidos con DAPI y evaluados bajo fluorescencia para determinar el porcentaje de maduración nuclear por la expulsión del primer cuerpo polar. El rolipram 50µM estimuló la maduración nuclear de oocitos bovinos de una manera similar a la obtenida con las gonadotropinas (76.83 vs 79.46%, p>0.05), pero en mayor medida que la observada con rolipram 25 y 75 µM (31.25, y 28.61%, respectivamente). Los CCOs cultivados en presencia de rolipram 50 µM+Gonadotropinas maduraron en menor proporción (63.74%) comparada con gonadotropinas (p<0.01) o rolipram 50 µM (p<0.05). Los resultados permiten concluir que el porcentaje de maduración nuclear in vitro de oocitos bovinos depende de la dosis de rolipram utilizada, donde la concentración de rolipram 50 µM presentó un comportamiento similar a las gonadotropinas en la maduración del oocito. Además, la presencia del estímulo gonadotrópico y del inhibidor de PDE puede inducir una respuesta de desensibilización de la vía del AMPc, reflejada en una disminución del porcentaje de maduración.
Palabras clave: AMPc intracelular, complejos cúmulo-oocito, enzima fosfodiesterasa, meiosis bovina, metafase II
]]>Summary
Gonadotropic follicle stimulating hormone (FSH) and lutenizing hormone (LH) induce intracellular cyclic AMP (cAMP) production during the in vitro maturation (IVM) of bovine oocytes. Cyclic AMP exerts a dual effect, where high intraoocyte cAMP levels are responsible for oocyte meiotic blockage, while high cAMP levels into the granulose cells induce oocyte maturation. Intracellular cAMP levels are regulated by phosphodiesterases (PDE)-mediated hydrolysis, enzymes having a specific follicle expression pattern. Oocyte expresses typo 3 PDE (PDE 3), while granulose cells expresses type 4 PDE (PDE 4). With the aim to test the effect of the specific PDE 4 inhibitor rolliprom on percentage in vitro nuclear maturation (IVNM) of bovine oocytes, 629 cumulus-oocyte complexes (COC) were cultured at 38.5 °C/CO2 5%/24 h on TCM-199 medium with pFSH and hrLH with or without rolipram. Experimental groups were: gonadotrophins alone, gonadotropins + rolipram (25, 50, or 75 µM), rolipram 50 µM + gonadotrophins, and control (media without stimulus). In order to determinate the nuclear maturation percentage by the first polar body expulsion, oocytes were dyed with DAPI and evaluated by fluorescence. Rolipram 50 µM stimulated bovine oocyte nuclear maturation in a similar way to gonadotrophins stimulus (76.83 vs. 79.46%, p>0.05) did, but in a higher way than rolipram 25 µM (31.25%) or 75 µM (28.61%). The COC cultured with rolipram 50 µM+gonadotrophins maturated in a lower proportion (63.74%) than did with gonadotropins (p<0.01) or rolipram 50 µM (p<0.05). A dose-dependent response of percentage of IVNM of bovine oocytes was detected. Thus rolipram 50 µM, exerts a similar effect of gonadotropins on oocyte maturation. In addition, the presence of gonadotropic stimulus and a PDE inhibitor may induce a desensitization response of the cAMP way, as suggested by the reduction of the percentage of maturation.
Key words: bovine meiosis, cumulus-oocyte complex, intracellular cAMP, metaphase II, phosphodiesterase enzyme
Resumo
Durante o processo de maturação in vitro de oocitos, as gonadotrofinas FSH e LH induzem a produção de AMPc. O AMPc tem duplo efeito, pois os altos níveis de AMPc intraoocitario mantém o bloqueio meiótico, enquanto que nas células da granulosa induzem a maturação do oocito. Os níveis de AMPc são regulados pela hidrólise mediada das fosfodiesterasas (PDE), as quais apresentam expressão específica no folículo, o oocito expressa a PDE 3, enquanto que as células da granulosa PDE 4. Com o objetivo de avaliar o efeito do rolipram, um inibidor específico de PDE 4, sobre a percentagem de maturação nuclear in vitro (MNIV) de oocitos bovinos, 629 complexos cúmulo oocito (CCO) foram cultivados a 38.5 °C/5% CO2/24 h, em meio TCM-199 com a adição de pFSH e hrLH, o rolipram. Os grupos experimentais foram: adição de gonadotrofinas, rolipram (25, 50 ó 75 µM), rolipram 50 µM + gonadotrofinas, ou controle sem estímulo. Os oocitos foram tingidos com DAPI e avaliados sob fluorescência para determinar a percentagem de maturação nuclear pela expulsão do primeiro corpo polar. O rolipram 50µM estimulou a maturação nuclear de oocitos bovinos de maneira similar a obtida com as gonadotrofinas (76.83 vs 79.46%, p>0.05), porém em maior medida que a observada com rolipram 25 y 75 µM (31.25, y 28.61%, respectivamente). Os CCOs cultivados na presença de rolipram 50 µM+Gonadotrofinas maturaram em menor proporção (63.74%) quando comparado com gonadotrofinas (p<0.01) ou rolipram 50 µM (p<0.05). Os resultados permitem concluir que a percentagem de maturação nuclear in vitro de oocitos bovinos depende da doses de rolipram utilizada, sendo que a concentração de rolipram 50 µM apresentou um comportamento similar às gonadotrofinas na maturação do oocito. Ale disto, a presença do estímulo gonadotrópico e do inibidor de PDE podem induzir uma resposta de diminuição na sensibilização da via do AMPc, refletida numa diminuição da percentagem de maturação.
Palavras chave: AMPc intracelular, complexos cúmulo-oocito, enzima fosfodiesterasa, meiosis bovina, metafase II
]]>
Introducción
La principal función de la gónada femenina es la diferenciación y liberación de un oocito maduro para la fertilización y propagación de la especie. En bovinos, este proceso inicia con la formación de las unidades foliculares en la vida fetal, continúa con el reclutamiento de un grupo o cohorte de folículos preantrales, su crecimiento, selección del folículo dominante, hasta llegar a la ovulación (7). Este proceso es regulado por un ambiente de hormonas gonadotrópicas hormona foliculo estimulante (FSH) y hormona luteinizante (LH), hormonas esteroidales y diversos factores de crecimiento endocrinos y paracrinos (19).
La maduración del oocito es un fenómeno complejo que abarca dos procesos bioquímicamente caracterizados: el primero, la maduración citoplásmica que consiste en las modificaciones ultraestructurales de las organelas y la acumulación de macromoléculas necesarias para apoyar la maduración nuclear, la fertilización y el desarrollo embrionario temprano; y el segundo, es la maduración nuclear, en la cual el oocito que se encuentra detenido en profase I continúa hasta el estadío de metafase II, evidenciado por la expulsión del primer cuerpo polar (6). Ambos procesos de maduración, citoplásmica y nuclear, son regulados por las hormonas gonadotrópicas FSH y LH (32).
El pico preovulatorio de LH induce cambios marcados en el folículo y en el complejo cúmulo oocito (CCO). Entre ellos, está el desacoplamiento de las uniones gap en las células del cúmulo, la expulsión del oocito desde el folículo ovárico y la continuación de la meiosis en el oocito hasta metafase II (27, 30), adquiriendo la maduración nuclear y capacitándolo para interactuar con el espermatozoide y formar el nuevo embrión (6, 8).
En los procesos de maduración in vitro (MIV), los complejos cúmulo oocito (CCO) aspirados de folículos con un diámetro entre 3 y 8 mm, completan su maduración en 24 horas, mediante el uso de gonadotropinas FSH y LH (5, 32), cuya acción sobre los complejos oocito-cúmulo es mediada a través de la vía del 3´,5´-adenosín monofosfato cíclico (AMPc). Este AMPc tiene un efecto dual dependiendo del compartimento del CCO: el oocito o las células del cúmulo. Por lo tanto, los altos niveles de AMPc al interior del oocito mantienen su freno meiótico en profase I, mientras que los altos niveles de AMPc en las células de la granulosa inducen la continuación de la meiosis, caracterizada por la disolución de la vesicula germinal (GVBD, Germinal Vesicle BreakDown), condensación cromosómica, formación de las fibras del uso, poliadenilación de RNAm que codifica para la kinasa c-Mos y la expulsión del primer cuerpo polar (13, 15, 28).
En la continuación de la meiosis, los niveles de AMPc intraoocito descienden por dos mecanismos. El primero es la reducción del flujo de AMPc desde las células de la granulosa hacia el oocito debido al desacoplamiento de las uniones gap, y el segundo es la hidrólisis de AMPc a su forma inactiva 5´-AMP, reacción catalizada por fosfodiesterasas (PDE, Phosphodiesterases) (3). La expresión de éstas enzimas es específica de tejido en la unidad folicular, el oocito mamífero expresa la PDE tipo 3, mientras que en las células de la granulosa se expresa la PDE tipo 4 (17).
Debido a la expresión específica de PDE en la unidad folicular, se han evaluado medicamentos que permitan una inhibición selectiva, aumentando la concentración intracelular de AMPc, ya sea en las células de la granulosa o en el oocito, modulando diferentes funciones como la esteroidogénesis (12, 24) y la meiosis (17, 28). En la modulación de la meiosis por estos medicamentos se han planteado dos áreas de investigación, la primera con el objetivo de inhibir la PDE 3 en el oocito, manteniendo el bloqueo meiótico, en donde se han evaluado medicamentos como la milrinona, la cilostamida y el ORG 9935, entre otros (18, 28, 33), y la segunda con el objetivo de inhibir la PDE 4 en células de la granulosa, induciendo la continuación de la meiosis con medicamentos como el rolipram y el Ro171424 (2, 28, 31).
Teniendo en cuenta los planteamientos anteriormente descritos, el objetivo de este trabajo fue determinar si el Rolipram, un inhibidor específico de PDE 4, puede ser utilizado como una alternativa a las gonadotropinas en la inducción de la maduración nuclear in vitro de oocitos bovinos.
]]> Materiales y métodos
Obtención y transporte del material de estudio
Los ovarios de bovino fueron obtenidos de hembras en ciclo reproductivo que fueron sacrificadas en la Central Ganadera de Medellín. Estos ovarios fueron transportados en solución de tampón fosfato salino (PBS, Phosphate buffer saline) estéril a 37 °C, al Laboratorio de Biotecnología Animal de la Universidad Nacional sede Medellín, para su procesamiento.
Obtención de complejos cúmulo oocitos (CCO)
Los folículos entre 3 y 6 mm de diámetro, fueron aspirados con aguja calibre 18 y jeringa de 5 ml, el líquido se recolectó en tubos cónicos de 15 ml/37 °C y se centrifugó a 1500 rpm/5 min. El precipitado se resuspendió en 1 ml de líquido folicular y se depositó en una caja de Petri estéril. Con estereomicroscopio se seleccionaron los CCOs de buena calidad (varias capas de células de la granulosa, cúmulo compacto y citoplasma homogéneo) según los criterios previamente establecidos (9).
Cultivo de los CCOs
Los CCOs seleccionados se lavaron tres veces en medio TCM-199 con sales de Hank´s (Sigma Ref M3274) y fueron cultivados en medio TCM-199 con sales de Hank´s suplementado con 27.9 µg/ml de ácido pirúvico (Sigma Ref p4562), 29.2 µg/ml de glutamina (SIGMA Ref G3126), 1 µg/ml de estradiol (Sigma Ref E2257), suero bovino fetal 10% (Sigma Ref F1051) y cada uno de los diferentes tratamientos (véase Tabla 1). Las condiciones de cultivo fueron de 38.5 °C, 5% de CO2 y 90% de humedad relativa. Al finalizar el tiempo de maduración de 24 horas, los CCOs se evaluaron para el estado de la meiosis (31).
]]> Evaluación de la maduración in vitro
Al terminar el tiempo de maduración, los oocitos fueron desnudados en hialuronidasa (Sigma Ref H4272) al 2% (27). Posteriormente, fueron teñidos con DAPI 5µg/ml (Sigma, Ref D9564). La observación se realizó en un microscopio de fluorescencia (CarlZeiss Oxilab HBO 50/AC, Alemania), donde se determinó la presencia o ausencia del primer cuerpo polar (22, 23).
Análisis estadístico
El experimento se realizó bajo un diseño de bloques al azar, en el que el efecto del rolipram durante la maduración se evaluó a través del porcentaje de oocitos en metafase II (presencia del primer cuerpo polar). Las diferencias de medias entre los tratamientos se evaluaron mediante la prueba de Tukey. Todos los datos fueron procesados en el programa estadístico SAS System, versión 8 2001, y para todas las decisiones se tuvo en cuenta un alfa de 0.05 (p<0.05).
Resultados
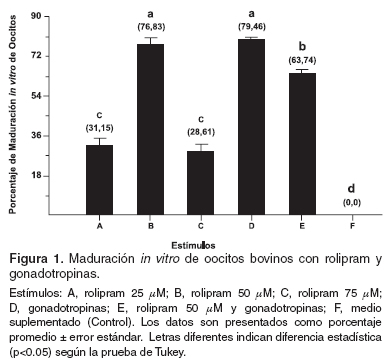
En el estudio se trabajó con un total de 629 CCOs que fueron distribuidos en seis tratamientos de nueve repeticiones cada uno. Al terminar el periodo de maduración de 24 horas y establecer el porcentaje de maduración mediante la presencia del cuerpo polar, los datos fueron sometidos a un análisis de varianza (Anova), cuyos resultados indicaron que existen diferencias significativas entre los tratamientos (p<0.01). Estos resultados permitieron agrupar los datos para cada tratamiento y realizar un análisis de comparación de medias con la prueba de Tukey. Cuando se compararon los grupos cultivados en presencia de rolipram, se encontró que este tuvo efecto sobre la maduración cuando se comparó con el control (véase Figura 1). El rolipram adicionado a una concentración 50 µM (76.83%), presentó un porcentaje significativamente mayor de maduración que a las concetraciones 25 (31.25%) ó 75 µM (28.61%) (p<0.01).
El porcentaje de maduración obtenido con gonadotropinas (79.46%) no presentó diferencia estadística cuando se comparó con el grupo de rolipram 50 µM (76.83%), pero si con 25 (31.25%) y 75 µM (28.61%). Cuando se maduraron los CCOs en presencia de gonadotropinas y rolipram 50 µM, se encontró un porcentaje de maduración significativamente menor (63.74%) que el obtenido sólo con gonadotropinas (79.46%) o rolipram 50 µM (76.83%) (véase Figura 1).
Discusión
]]> En los protocolos de maduración in vitro de CCOs bovinos se han tenido como referencia diferentes estados de la meiosis como indicadores de la maduración nuclear, tal como la disolución de la GVBD (17, 28), metafase I y metafase II (presencia del primer cuerpo polar) (29), siendo este último el mejor criterio de maduración nuclear (26).
Estímulos: A, rolipram 25 µM; B, rolipram 50 µM; C, rolipram 75 µM; D, gonadotropinas; E, rolipram 50 µM y gonadotropinas; F, medio suplementado (Control). Los datos son presentados como porcentaje promedio ± error estándar. Letras diferentes indican diferencia estadística (p<0.05) según la prueba de Tukey.
Debido a la expresión específica de PDEs en la unidad folicular, PDE-3 en el oocito y PDE-4 en las células de la granulosa (31), es posible modular la actividad de la PDE-4 con inhibidores específicos, entre ellos el Rolipram (10), para evaluar su efecto sobre la maduración del oocito. El uso de Rolipram permite aumentar los niveles de AMPc de manera selectiva en las células de la granulosa de los COC (25). Al utilizar una concentración de 50 µM de Rolipram, se induce la acumulación de AMPc en las células de la granulosa de los COC, disminuyen la cantidad de uniones gap (29) y aumenta la expansión del cúmulo. Esto reduce el flujo de AMPc hacia el oocito y permite la continuación de la meiosis, en un porcentaje del 76.83, similar al obtenido con gonadotropinas (79.46%). Estos resultados son similares a los reportados por Thomas et al (28), quienes utilizaron la misma concentración de rolipram (50 µM) y obtuvieron un porcentaje de maduración hasta metafase II del 84%.
La concentración óptima de rolipram para inducir la maduración del oocito puede depende de la especie: mientras en este trabajo se encontró el mayor porcentaje de maduración de oocitos bovinos hasta metafase II con 50 µM, similar al obtenido por Thomas y col (28), en oocitos de rata se obtuvo la mejor respuesta de reanudación de la meiosis (GVBD) con rolipram a 0.5 y 5 µM (31). Esta respuesta dependiente de especie, puede ser debida a varios aspectos como son: la expresión de diferentes subtipos de PDE-4 (A, B, C y D) o de variantes generadas por splicing alternativo, lo cual genera una gran diversidad de isoformas de fosfodiesterasas con diferentes mecanismos de regulación de la actividad catalítica (4, 21); el nivel de expresión, lo cual sugiere que se requiere alta concentración del inhibidor cuando hay mayor expresión de PDE-4; y la presencia de polimorfismos entre las especies, que puede generar una respuesta diferencial con la concentración del inhibidor.
Al utilizar las concentraciones de 25 y 75 µM de rolipram se encontró un porcentaje de maduración menor (31.25 y 28.61%, respectivamente) que el obtenido con 50 µM (76.83%). El resultado obtenido con rolipram 25 µM posiblemente se debe a que es una baja concentración y no es suficiente para inhibir la PDE-4 de las células de la granulosa, como se sugiere por la baja expansión del cúmulo, que a su vez estría explicado porqque las uniones gap y por lo tanto el flujo de AMPc de las células de la granulosa hacia el oocito permanecen y mantienen el bloqueo meiótico (32). Esta respuesta es similar a la descrita por Tsafriri et al (31), quienes al evaluar la concentración de 0.05 µM de rolipram sobre los oocitos de rata, obtuvieron sólo el 20% de reanudación de la meiosis (GVBD). Una baja respuesta también se encontró al cultivar los COCs en presencia de 75 µM de rolipram, cuando se obtuvo 28.61% de maduración de oocitos bovinos hasta metafase II. Este efecto fue similar al observado en los complejos cultivados con rolipram 25 µM, cuando presentaron baja expansión del cúmulo y bajo porcentaje de maduración. Dicha respuesta se debe probablemente a que la concentración 75 µM induzca una desensibilización de la vía del AMPc en las células de la granulosa, mediada por fosforilación de la proteína G (1) o de la adenil ciclasa (16), y adicionalmente a efectos no específicos de inhibición sobre la PDE-3, que aumenten los niveles de AMPc dentro de este y mantengan su bloqueo meiótico (28, 33). El efecto del rolipram 75 µM fue similar al reportado por Tsafriri et al (31), quienes encontraron una disminución significativa en la reanudación de la meiosis de oocitos de rata al incubarlos con rolipram 50 µM.
Cuando se cultivaron los CCOs en presencia de gonadotropinas y rolipram 50 µM, se encontró un porcentaje de maduración significativamente menor (63.74%) que el obtenido con gonadotropinas (79.46%) o rolipram 50 µM (76.83%), lo que sugiere que el rolipram 50 µM inhibe parcialmente la maduración del oocito inducida por gonadotropinas. Esta reducción de la respuesta gonadotrópica puede ser debida a una desensibilización mediada por diferentes mecanismos, tales como: la fosforilación en los aminoácidos serina y treonina del asa 3i citoplasmática y de la cola carboxilo terminal de los receptores de FSH y LH, evitando la interacción de los receptores con la proteína G y la activación de la adenil ciclasa, responsable de la síntesis de AMPc (1, 4); la fosforilación mediada por proteína Gs, impidiendo la activación de la adenil ciclasa (1); la interacción de la proteína arrestina 2 con la lupa 3i citoplasmática del receptor de LH, la cual el bloquea el acoplamiento del receptor de LH a proteína G y por ende la activación de la adenil ciclasa (20); y la internalización de receptores mediada por proteínas adaptadoras y clatrina, lo cual disminuye la presencia de receptores en la membrana celular para las hormonas gonadotrópicas y la activación de la producción de AMPc (11). Resultados de desensibilización fueron reportados por Tsafriri et al (31), quienes encontraron un efecto inhibitorio de la maduración nuclear de CCOs de ratas cuando eran cultivados con LH y rolipram 50 µM. Además, se ha reportado una baja respuesta de maduración cuando se combina el Rolipram y un estimulador de la síntesis de AMPc (forskolina, activador de la adenil ciclasa) (28).
Estos resultados sugieren que la maduración nuclear del oocito, inducida por gonadotropinas, puede ser mimetizada con inhibidores específicos de PDE tipo 4, como el rolipram (50 µM), y confirma la compartamentalización de las fosfodiesterasas (18, 31) y del AMPc en la unidad folicular (3, 18). Además, sugiere que el rolipram (50 µM) no tiene un efecto sinérgico a la acción gonadotrópica, por lo que puede ser utilizado como único estímulo para inducir la maduración nuclear del oocito de bovino. Aunque estos hallazgos son un avance significativo en el conocimiento de la modulación en la señalización hormonal de la maduración nuclear del oocito, se requieren más estudios en los que se determine la expresión de los subtipos de PDE tipo 4 (21) en las células de la granulosa de la unidad folicular bovina y el efecto de inhibidores como el CDP-840 (14), el RO-20-1724 (2), la zardaverina y el rolipram (10), sobre estos subtipos, en los procesos de fertilización y desarrollo embrionario temprano in vitro y su posible uso en el desarrollo de métodos alternativos de producción in vitro de embriones en reproducción asistida en animales y humanos.
]]>Agradecimientos
Los autores expresan sus agradecimientos a la Dirección de Investigación de la Universidad Nacional de Colombia, sede Medellín - DIME (030802736), a la Central Ganadera de Medellín y al Laboratorio de Biotecnología Animal.
Referencias
1. Ámsterdam A, Hanoch T, Dantes A, Tajima K, Strauss III J, et al. Mechanisms of gonadotropin desensitization. Mol Cell Endocrinol 2002; 187:69-74. [ Links ]
2. Bilodeau-Goeseels S. Effects of phosphodiesterase inhibitors on spontaneous nuclear maturation and cAMP concentrations in bovine oocytes. Theriogenology 2003; 60:1679-1690. [ Links ]
3. Conti M, Andersen CB, Richard F, Mehats C, Chun SY, et al. Role of cyclic nucleotide signaling in oocyte maturation. Mol Cell Endocrinol 2002; 187:153-159. [ Links ]
4. Conti M, Nemoz G, Sette C, Vicini E. Recent progress in understanding the hormonal regulation of phosphodiesterases. Endocrine Rev 1995; 16:370-389. [ Links ]
5. Eckert J y Niemann H. In vitro maturation, fertilization and culture to blastocysts of bovine oocyte in protein-free media. Theriogenology 1995; 43:1221-1225. [ Links ]
6. Eppig JJ. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod Fertil Dev 1996; 8:485-489. [ Links ]
7. Gougeon A. Regulation of ovarian follicular development in primates: facts and hypotheses. Endocrine Rev 1996; 17:121-155. [ Links ]
8. Greenwald GS, Roy SK. Follicular development and its control. In: Knobill E, Neill JD (Eds). The physiology of reproduction, Vol. I. 2nd ed. New York: Raven-Press; 1994. p.629-724. [ Links ]
9. Hawk HW, Wall RJ. Improved yields of bovine blastocysts from in vitro-produced oocytes. I. Selection oocytes and zygotes. Theriogenology 1994; 41:1571-1583. [ Links ]
10. Houslay MD, Adams DR. PDE4 cAMP phosphodiesterases: modular enzymes that orchestrate signaling, cross-talk, desensitization and compartmentalization. Biochem J 2003; 370:1-18. [ Links ]
11. Hunzicker-Dunn M, Gurevich V, Casanova JE, Mukherjee S. ARF6: a newly appreciated player in G protein-coupled receptor desensitization. FEBS Lett 2002; 521:3-8. [ Links ]
12. Jensen JT, Schwinof KM, Zelinski-Wooten MB, Conti M, DePaolo LV, et al. Phosphodiesterase 3 inhibitors selectively block the spontaneous resumption of meiosis by macaque oocytes in vitro. Hum Reprod 2002; 17:2079-2080. [ Links ]
13. Jones KT. Turning it on and off: M-phase promoting factor during meiotic maturation and fertilization. Mol Hum Reprod 2004; 10:1-5. [ Links ]
14. Jones TR, McAuliffe M, McFarlane CS, Piechuta H, Macdonald D, et al. Effects of a selective phosphodiesterase IV inhibitor (CDP-840) in a leukotriene-dependent non-human primate model of allergic asthma. Rev Can Physiol Pharmacol 1998; 76:210-217. [ Links ]
15. Josefsberg LB, Dekel N. Translational and post-translational modifications in meiosis of the mammalian oocyte. Mol Cell Endocrinol 2002; 187:161-171. [ Links ]
16. Lai HL, Yang TH, Messing RO, Ching YH, Lin SC, et al. Protein kinase C inhibits adenylyl cyclase type VI activity during desensitization of the A2a-adenosine receptor-mediated cAMP response. J Biol Chem 1997; 272:4970-4977. [ Links ]
17. Mayes M. The meiotic arrest of bovine oocytes. Du grade de Philosophie Doctor (Ph.D.), Université Laval - Québec, Canadá, 2002. 171 p. [ Links ]
18. Mayes MA, Sirard MA. Effect of type 3 and type 4 phosphodiesterase inhibitors on the maintenance of bovine oocytes in meiotic arrest. Biol Reprod 2002; 66:180-184. [ Links ]
19. McGee EA, Hsueh AJW. Initial and cyclic recruitment of ovarian follicles. Endocr Rev 2000; 21:200-214. [ Links ]
20. Mukherjee S, Palczewski K, Gurevich V, Hunzicker-Dunn M. -Arrestin-dependent desensitization of luteinizing hormone/choriogonadotropic receptor is prevented by a synthetic peptide corresponding to the third Intracellular loop of the receptor. J Biol Chem 1999; 274:12984-12989. [ Links ]
21. Müller T, Engels P, Fozard Jr. Subtypes of the type 4 cAMP phosphodiesterases: structure, regulation and selective inhibition. Trends Pharmacol Sci 1996; 17:294-298. [ Links ]
22. Olson SE, Seidel GE Jr. Culture of in vitro-produced bovine embryos with vitamin E improves development in vitro and after transfer to recipients. Biol Reprod 2000; 62:248-252. [ Links ]
23. Pursel VG, Wall RJ, Rexroad CE, Hammer RE y Brinster RL. A rapid whole-mount staining procedure for nuclei of mammalian embryos. Theriogenology 1985; 24:687-692. [ Links ]
24. Reddoch RB, Armstrong DT. Interactions of a phosphodiesterase inhibitor, 3-isobutyl-1-methyl xanthine, with prostaglandin E2, follicle-stimulating hormone, luteinizing hormone, and dibutyryl cyclic 3',5'-adenosine monophosphate (cAMP) in cAMP and steroid production by neonatal rat ovaries in vitro. Endocrine 1984; 115:11-18. [ Links ]
25. Richard FJ, Tsafriri A, Conti M. Role of phosphodiesterase type 3A in rat oocyte maturation. Biol Reprod 2001; 65:1444-1451. [ Links ]
26. Smith GD. In vitro maturation of oocytes. Curr Womens Health Rep 2001; 1:143-151. [ Links ]
27. Suzuki H, Jeong BS, Yang X. Dynamic changes of cumulus-oocyte cell communication during in vitro maturation of porcine oocytes. Biol Reprod 2000; 63:723-729. [ Links ]
28. Thomas RE, Armstrong DT, Gilchrist RB. Differential effects of specific phosphodiesterase isoenzyme inhibitors on bovine oocyte meiotic maturation. Dev Biol 2002; 244:215-225. [ Links ]
29. Thomas RE, Armstrong DT, Gilchrist RB. Bovine cumulus cell-oocyte Gap junctional communication during in vitro maturation in response to manipulation of cell-specific cyclic adenosine 3,5-monophosophate levels. Biol Reprod 2004; 70:548-556. [ Links ]
30. Tsafriri A, Dekel N. Molecular mechanisms in ovulation. In: Molecular Biology of the Female Reproductive System. New York:Academic Press; 1994. p.207-258. [ Links ]
31. Tsafriri A, Chun SY, Zhang R, Hsueh AJ, Conti M. Oocyte maturation involves compartmentalization and opposing changes of cAMP levels in follicular somatic and germ cell: studies using selective phosphodiesterase inhibitors. Dev Biol 1996; 178:392-402. [ Links ]
32. Wassarman PM, Albertini DF. The mammalian ovum. In: Knobill E, Neill JD (Eds.). The physiology of reproduction, Vol. I. 2nd ed. New York: Raven-Press; 1994. p.79-122. [ Links ]
33. Wiersma A, Hirsch B, Tsafriri A, Hanssen RG, van de Kant M, et al. Phosphodiesterase 3 inhibitors suppress oocyte maturation and consequent pregnancy without affecting ovulation and cyclicity in rodents. J Clin Invest 1998; 102:532-537. [ Links ]
¶ Para citar este artículo: López YS, Mejía AM, Biol Escobar EE, Agudelo Jaramillo B, Vásquez Araque NA, Echavarría H. Efecto del inhibidor de fosfodiesterasa tipo 4 —rolipram, sobre la maduración in vitro de oocitos bovinos. Rev Colomb Cienc Pecu 2008; 21:59-65.
]]> * Autor para el envío de la correspondencia y la solicitud de separatas: Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia sede Medellín. AA 3840, Medellín, Colombia. E-mail: nvasquez@unalmed.edu.co ]]>