Genesinvolved in pathogenesis, persistence, and excretion ofSalmonella in animal models¤
Genes involucrados en la patogénesis, persistencia y excreción de Salmonella en modelos animales
Genes envueltos em patogênese, persistencia e excreção de Salmonella em modelos animais
Gabriela Silva1*, MVZ, MSP-E; Héctor S López2, QFB, Dr. Sci
1 Programa Regional del Noroeste. Doctorado en Ciencias. Especialidad Biotecnología. Universidad Autónoma de Sinaloa, México.
2 Laboratorio de Inmunología Molecular, Facultad de Ciencias Químico Biológicas, Universidad Autónoma de Sinaloa. Culiacán, Sinaloa, México.
(Recibido: 6 noviembre, 2010; aceptado: 14 junio, 2011)
Infectious diseases can be studied from different perspectives; from the global epidemiology to the detailed interactions between the pathogen and the host cell. Central to a complete understanding of any disease is the ability to integrate information from different points of view into a coherent model that fully explains the disease process. In vivo infections have been studied mainly in murine models, in which the possibility of genetically manipulating either the bacterium or the host allows variations in the severity of the host–pathogen combination and facilitates the evaluation of individual bacterial and host traits. Gross parameters, such as host mortality, clinical signs, overall bacterial numbers in the tissues, pathological changes, shedding patterns, production of immune mediators, as well as fluctuations and phenotypic profiles of cell populations have been widely used in the descriptive analysis of in vivo infection pathogenesis. However, key factors, such as bacterial location, spatiotemporal patterns of spread and tissue distribution, as well as sites of microbial persistence, have not been sufficiently studied despite their importance in terms of targeted medical intervention. This review aims to give a broad overview of the molecular basis associated with Salmonella infection in murine and other experimental animal models, focusing on organ location, and bacteria spread and distribution. The special conditions that promote bacteria retention and excretion in asymptomatic animals are also highlighted. It aims to gain a better understanding of the infectious process, so that comprehensive reassessment of pathogen behavior allow the establishment of preventive medicine programs in both humans and domestic or wild animals.
Key words: animal models, genes, pathogenesis, Salmonella.
Resumen
Las enfermedades infecciosas pueden ser estudiadas desde diferentes puntos de vista, por ejemplo, el aspecto epidemiológico global o bien el que detalla las interacciones entre el agente patógeno y la célula hospedero. Para tener un entendimiento completo, resulta necesario, por lo tanto, integrar la información de todos los puntos de vista, conformando un modelo coherente que explique completamente el proceso de enfermedad. El seguimiento de las infecciones in vivo ha sido estudiada principalmente en modelos murinos, los cuales ofrecen la posibilidad de manipulación genética tanto en las bacterias como en el hospedero, variando de esta manera la severidad de la enfermedad y facilitando la evaluación de su comportamiento. Los parámetros epidemiológicos o anatomopatológicos, tales como mortalidad, semiología clínica, evidencias bacterianas en tejidos, cambios histopatólogicos, patrones de excreción, producción de mediadores inmunes y fluctuaciones y cambios fenotípicos en poblaciones celulares, han sido ampliamente utilizados para el análisis descriptivo de la patogenia de las infecciones in vivo. De cualquier forma, los puntos clave, como patrones espaciotemporales de diseminación en tejidos, y sitios anatómicos de persistencia microbiana, no han sido profundamente estudiados, a pesar de la importancia que esto implica en el desarrollo de políticas de intervención sanitaria. Esta revisión pretende dar un amplio panorama sobre las bases moleculares asociadas a la infección por Salmonella en modelos murinos, así como en otros modelos animales experimentales, considerado desde el nivel celular, localización, diseminación y distribución de la bacteria en órganos y rescatando aquellas condiciones especiales que promueven su permanencia y excreción en el organismo animal asintomático. Se pretende abordar este proceso infeccioso desde una visión que oriente hacia un mejor entendimiento, de tal manera que, una reevaluación integral del comportamiento del agente patógeno, permitirá el establecimiento de programas de medicina preventiva tanto en seres humanos como en animales domésticos o silvestres.
Palabras clave: genes, modelos animales, patogénesis, Salmonella.
Resumo
As doenças infecciosas podem ser estudadas desde diferentes pontos de vista, como por exemplo: o aspecto epidemiológico global ou as interacções entre o agente patogênico e a célula hospedeira. Para ter um entendimento completo, é necessário, integrar a informação de todos os pontos de vista, formando um modelo coerente e que explique completamente o processo de a doença. Geralmente os modelos das infecções in vivo são estudados principalmente em modelos murinos, os quais oferecem a possibilidade de manipulação genética, tanto nas bactérias como no hospedeiro, variando desta maneira a severidade da doença e facilitando a avaliação de seu comportamento. Os parâmetros epidemiológicos ou anatomopatológicos como: mortalidade, semiología clínica, evidências bacterianas em tecidos, mudanças histopatólogicos, padrões de excrecão, produção de mediadores imunes, flutuações e mudanças fenotípicos em populações celulares foram amplamente utilizados para a análise descritiva da patogenia das infecçõesin vivo. De qualquer forma, os pontos chaves como padrões espaco-temporais de disseminacão em tecidos, e lugares anatômicos de persistencia microbiana não foram profundamente estudados, apesar da importância que isto implica no desenvolvimento de políticas de intervenção sanitária. Esta revisão tem como objetivo dar um amplo panorama sobre as bases moleculares da infecção por Salmonella em modelos murinos, bem como, em outros modelos animais experimentais, considerando desde o nível celular, localização, diseminação e distribuição da bactéria em órgãos e demonstrando as aquelas condições especiais que promovem sua permanência e excreção no organismo animal assintomático. O processo infeccioso foi abordado de maneira que proponha um melhor entedimento do mesmo, uma reavaliação integral do comportamento do agente patogénico que permita o estabelecimento de programas de medicina preventiva, tanto em seres humanos, como em animais domésticos ou silvestres.
]]> Palavras chave: genes, modelos animais, patogênese, Salmonella.§ Para citar este artículo: Silva G, López HS. Genes involucrados en la patogénesis, persistencia y excreción de Salmonella en modelos animales. Rev Colomb Cienc Pecu 2012; 25:107-122
* Autor para correspondencia: Gabriela Silva. Carretera Internacional Sur Km 3.5, Culiacón, Sinaloa, México. E-mail: gaby@uas.uasnet.mx.
Introducción
Salmonella enterica es una bacteria Gram- negativa que causa una amplia variedad de enfermedades tanto en seres humanos como en animales de todo el mundo. Se reportan aproximadamente 28 millones de casos de fiebres entéricas (fiebre tifoidea y paratifoidea) al año (Crump et al., 2004). Las serovariedades no tíficas, pueden causar gastroenteritis en los seres humanos y animales, y pueden ser causa común de septicemias mortales en individuos inmunodeprimidos o en los niños, especialmente en los países en desarrollo (Kankwatira et al., 2004; Maclennan et al., 2008). Actualmente los análisis comparativos entre los mecanismos inmunológicos y genéticos que regulan la interacción de S.enterica en ratones, seres humanos y animales domésticos revelan sorprendentes similitudes. El uso de técnicas moleculares de bioimágen, en combinación con la resolución y el poder predictivo de modelos matemáticos, ha permitido conocer la dinámica espacio-temporal de las distintas poblaciones bacterianas durante la infección. Estos enfoques permiten visualizar el proceso de infección desde un punto de vista exhaustivo e integral (Grenfell et al., 2004).
Patogénesis, persistencia y excreción
Patogénesis
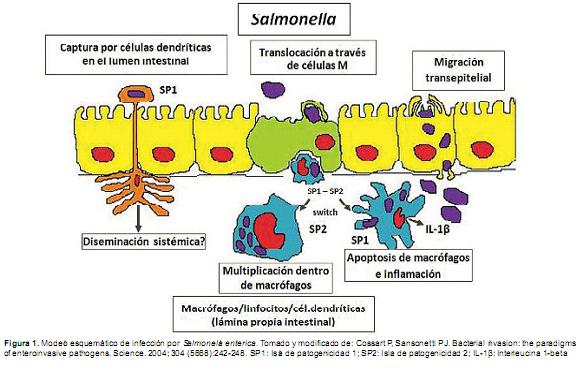
Invasiónintestinalydiseminaciónsistémica.Las infecciones por S.entericase adquieren usualmente a través de la vía oral. La evidencia en modelos murinos de infecciones tíficas sistémicas, indican que algunos de los microorganismos ingeridos invaden los enterocitos y las células M de la mucosa intestinal, y en la submucosa S. enterica es fagocitada por macrófagos fijos. Los macrófagos mueren vía Caspasa 1 y la bacteria, ahora libre de su ambiente intracelular, se disemina en el torrente sanguíneo, entrando directamente a los vasos sanguíneos o bien a través de los vasos linfáticos y de los linfonodos mesentéricos (LnMs) (Figura 1).
]]> En la sangre S.entericaes reconocida como una bacteria extracelular o bien asociada a leucocitos CD18+ (Vázquez et al., 1999). S. enterica también puede alcanzar la submucosa intestinal a través de las células dendríticas, las cuales emiten prolongaciones a través de las uniones celulares de los enterocitos, y de esta manera diseminarse a sitios distantes y ocasionar infección sistémica (Rescigno et al., 2001). En el torrente sanguíneo, la bacteria puede ser opsonizada vía factores del complemento, y rápidamente es alejada de la sangre a una localización intracelular en el hígado, bazo ó médula ósea (Warren et al., 2002).
Mecanismos inmunológicos involucrados en la persistencia y excreción de Salmonella
Persistencia en macrófagos. S. enterica puede permanecer crónicamente en las células del sistema mononuclear fagocitario (SMF) hasta por un año después de la primoinfección. Se ha encontrado la bacteria a menudo secuestrada en macrófagos de LnMs. Se sabe que el interferón (IFN) es esencial para el control de la persistencia de la infección por S. enterica, la cual ha sido co-localizada dentro de macrófagos MOMA-2 (Anticuerpos anti Macrófago para diferenciar subpoblaciones de macrófagos, marcador útil para la detección amplia de monocitos y macrófagos en todas las cepas de ratón que permite una caracterización más precisa de los macrófagos tisulares fijos de varios órganos) en los senos subcapsulares y corteza de los linfonodos. La principal respuesta inmune que rodea a los macrófagos infectados es del tipo de linfocitos B, por lo tanto, Salmonella enterica serovar Typhimurium (S.Typhimurium) persiste intracelularmente en
macrófagos de LnMs de modelos murinos. Algunas otras serovariedades hospedero específicas de S. enterica como S. typhi, S. dublin, y S. pullorum pueden, después de la enfermedad clínica o infección asintomática, persistir en el organismo por largos períodos de tiempo. En el caso de S. typhi y S. dublin, la excreción bacteriana durante largos períodos de tiempo se asocia a una proliferación del agente patógeno, en este caso Salmonella, en vesícula biliar.
Se ha propuesto que el lugar anatómico donde reside S. enterica en animales excretores de bacterias vía fecal, y aún también en el humano, son los LnMs, y que algunas condiciones que promueven inmunosupresión, permiten la diseminación desde el SMF hacia el hígado, vesicular biliar, e intestino, promoviendo nuevamente la excreción bacteriana en heces, tal y como se presenta durante la fase primaria de la infección (Monack et al., 2004).
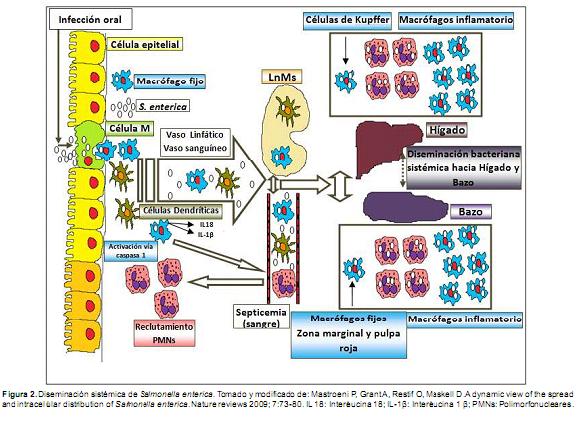
Recientemente se ha investigado el papel que juegan los macrófagos activados (MA) principalmente por interferón-gamma (IFN °) en la patogenia de la salmonelosis. Los MA son macrófagos que pueden fagocitar otros tipos celulares. Se ha propuesto que los MA constituyen un nicho de sobrevivencia intracelular durante las infecciones persistentes por S. Typhimurium. Resultados en investigaciones sugieren que los macrófagos en ratones infectados experimentalmente por S. Typhimurium fagocitan gran cantidad de diferentes tipos de leucocitos, los cuales, al igual que los macrófagos, proveen un excelente nicho de sobrevivencia intracelular (Fink y Cookson, 2005; Mastroeni et al., 1992; Muotiala y Makela, 1990). Se ha observado que los leucocitos fagocitados en los ratones infectados, se lisan por efecto de la fijación durante el procesamiento de los tejidos, y no por efecto de la fagocitosis en sí, lo que sugiere, que S. enterica puede reproducirse intracelularmente también en estos tipos celulares. Se desconoce si este fenómeno puede beneficiar al hospedero bajo ciertas circunstancias o en manifestaciones subclínicas.
La sobrevivencia de S. Typhimurium en macrófagos que han fagocitado células eucarióticas viables de ratón o ser humano sugiere que el fenómeno no se limita solamente al modelo murino, la fagocitosis de células viables y la posterior infección con S. Typhimurium puede inhibir que el macrófago elimine a la bacteria y de esta manera constituir un nicho. Esto resulta interesante, ya que la sobrevivencia de S. Typhimurium en MA puede jugar un papel importante en el establecimiento de infecciones crónicas. Experimentos futuros tendrán que resolver éstas incógnitas. Lo que se reconoce hasta este momento, es la interacción compleja que existe entre los macrófagos con otras células vivas eucarióticas y lo que puede implicar cambios en la activación de este mecanismo (Nix et al., 2007) (Figura 2).
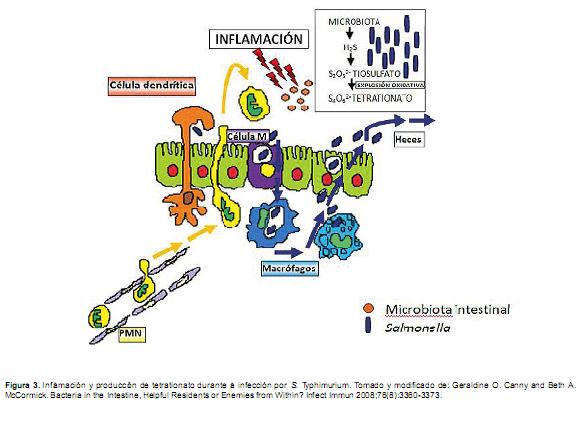
Papel del proceso inflamatorio en la persistenciaintestinal de Salmonella. Se ha encontrado que el proceso inflamatorio modifica el balance entre la microbiota intestinal y el patógeno S. Typhimurium a favor de éste. Este principio puede aplicarse a otros patógenos por lo que constituye un nuevo paradigma en la biología de las enfermedades infecciosas (Stecher et al., 2007). La microbiota residente y el patógeno compiten por el crecimiento. En un intestino sano existen interacciones benéficas entre la microbiota normal y la mucosa intestinal. La respuesta inflamatoria de los hospederos en este caso ocasionada específicamente por los factores de virulencia de S. Typhimurium (sistemas de secreción tipo 3- SST3) altera las condiciones en el lumen intestinal y activan la competencia a favor del patógeno.
]]> Hasta ahora, la infección por S. Typhimurium involucra dos pasos: dispara el proceso inflamatorio y sobrevive y se beneficia de un nicho ecológico alterado. Cómo es que la inflamación intestinal contribuye con el patógeno? El proceso inflamatorio involucra una secreción aumentada de péptidos antibacteriales, lecitinas, mucinas, infiltración y migración fagocitaria, dejando radicales libres de oxígeno y nitrógeno (Dann y Eckmann, 2007). Los factores antibacteriales libres pueden matar ó bien retardar el desarrollo de miembros específicos de la microbiota que normalmente pudieran inhibir el crecimiento de S. Typhimurium en un intestino saludable (Suzuki et al., 2004), es decir, pueden alterarse una ó más especies específicas de microbiota requeridas para el crecimiento eficiente de otras especies de microbiota que disminuyen el desarrollo del patógeno en un intestino saludable (Sonnenburg etal., 2005). Las especies bacterianas de la microbiota pueden ser más susceptibles a estos factores antibacteriales que el patógeno S. Typhimurium.Bajo estas condiciones, la microbiota simplemente se desarrolla más lentamente y por lo tanto el patógeno se multiplica exponencialmente (Stecher et al., 2007). La respuesta inflamatoria causada por el patógeno promueve su propia multiplicación en el lumen intestinal por medio de mecanismos desconocidos.Actualmente se ha observado que derivados reactivos del oxígeno generados durante la inflamación, reaccionan con componentes sulfurados luminales (tiosulfato) para formar un nuevo aceptor de electrones, el tetrationato. Los genes que confieren la capacidad de utilizar el tetrationato como aceptor de electrones hacen que S.Typhimurium crezca con ventaja por encima de la microbiota competidora del lumen del intestino inflamado. Es decir, los factores de virulencia de esta bacteria inducen la producción de este nuevo aceptor de electrones mediante el hospedador, lo que permite al patógeno utilizar la respiración (ciclo respiratorio oxidativo) para competir con los microorganismos de fermentación del intestino. Por tanto, la capacidad de desencadenar la inflamación intestinal es crucial para la biología de este patógeno (Figura 3) (Winter et al., 2010).
Mecanismos genéticos involucrados en la persistencia y excreción de Salmonella
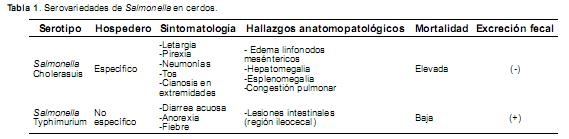
Basesmoleculares de diseminación y persistencia intestinal. Las serovariedades adaptadas a una gran variedad de hospederos (por ejemplo, S. Typhimurium), producen una enteritis aguda, mientras que las serovariedades hospedero-específicas (S.typhi) se asocian a enfermedad sistémica severa y la diarrea puede o no estar presente (Tabla 1) (Paulin etal., 2007; Davies etal., 2004).
Los mecanismos moleculares que expliquen la habilidad de las serovariedades de S. enterica, tanto para colonizar los intestinos de los animales domésticos, destinados a la producción de alimentos para el ser humano, así como, en algunos casos, su diseminación sistémica, son poco entendidos. Dentro de las amplias mutaciones en el genoma del serovar Typhimurium se han identificado grupos de genes involucrados en la colonización intestinal en los bovinos (Paulin et al., 2002), pollos (Morgan et al., 2004), cerdos (Carnell et al., 2007) y ratón (Lawley etal., 2006), y han revelado que el género Salmonellatanto en hospederos específicos como no específicos necesitan factores de colonización. La variación tanto en el repertorio genético, como en la secuencia y en la expresión de algunos factores pueden explicar las diferencias en la virulencia de algunas serovariedades de S. enterica. Se ha visto que las serovariedades hospedero específicas persisten más en LnMs, determinante importante para su virulencia sistémica (Paulin et al., 2002).
Por ejemplo, estudios experimentales han mostrado el comportamiento diferencial entre serovariedades hospedero específico y no específico. En la mucosa de íleon y colon de cerdos inoculados oralmente con cepas de serovar Typhimurium y de serovar choleraesuis, se observó una diferencia entre el número de unidades formadoras de colonias/gramo (UFC/g), en este caso las UFC/g del serovar Typhimurium fueron significativamente mayores a las 36 y 48 horas post-inoculación en comparación con el serovar choleraesuis.
Por otro lado, un elevado número de UFC/g del serovar choleraesuis fue recuperado de LnMs, tanto de íleon como de colon. Por consiguiente, la virulencia enterica del serovar Typhimurium se asocia con la rápida multiplicación en la mucosa intestinal, mientras que la virulencia sistémica del serovar choleraesuis se asocia con la persistencia en LnMs. También se observó una diferencia significativa entre estas dos cepas en cuanto al nivel de excreción fecal de bacterias, el serovar Typhimurium mostró una mayor excreción; las diferencias también pueden constatarse a nivel transcripcional, a las 24 horas post-inoculación, el serovar Typhimurium provocó altos niveles de transcripción de TNF, IL-8 y IL-18 comparado con el serovar choleraesuis, de tal manera que estos elevados niveles de citocinas pro-inflamatorias pueden afectar a la microbiota normal del intestino y promover la persistencia intestinal de la bacteria patógena (Paulin et al., 2007). Puede especularse entonces, que la rápida multiplicación del serovar Typhimurium podría desencadenar una respuesta inflamatoria temprana seguida de una infección de gran magnitud, lo que confinaría localmente el proceso a la mucosa intestinal, mientras que la lenta multiplicación del serovar choleraesuis le permitiría evadir la activación de la respuesta inmune innata, favoreciendo su diseminación disimulada (Uthe et al., 2007).
Por otro lado, genes del serovar Typhimurium codifican Sistemas de Secreción Tipo 3 (SST3), sistemas que juegan un papel importante en la colonización intestinal, así como en hígado y bazo. SST3-1 es un complejo proteico efector que estimula el re-arreglo de la actina en el citoesqueleto celular por lo que facilita la invasión bacteriana, además también contribuye a la multiplicación bacteriana en las células epiteliales (Steele- Mortimer et al., 2002). La invasión asociada al SST3 de S. Typhimurium es necesaria para la proliferación y la biogénesis de la vacuola intracelular en los enterocitos y el efector SopB del complejo proteico SST3-1 estimula la producción de óxido nítrico y permanece activo mucho después de la entrada de la bacteria (Drecktrah et al., 2005).
]]> Proteína ShdA. Los complejos de secreción tipo V (T5S) son la principal vía de transporte proteico a través de la membrana celular en las bacterias Gram negativas (Henderson et al., 2000). Por ejemplo, la proteína de autotransporte ShdA de S. Typhimurium se une a las proteínas de la matriz extracelular fibronectina y colágeno I, específicamente, se une a un grupo de residuos de aminoácidos cargados positivamente ubicados en el dominio Hep-2 de la fibronectina, la cual a su vez está involucrada también en la unión con la heparina (Kingsley et al., 2004). La expresión de la proteína ShdA de S.Typhimurium cepa ATCC14028 reproducida en el Laboratorio, no es detectable por medio de western blot, pero a través de la observación in vivo en intestino de modelo murino, se ha logrado establecer su asociación con la patogenia de la salmonelosis enterica.Actualmente se reconoce que se requiere ShdA para una eficiente y prolongada excreción de S. Typhimurium vía fecal (Kingsley et al., 2002). En los modelos murinos mutados de ShdA, la recuperación de S. Typhimurium tanto en intestino como en heces, se reduce considerablemente, lo que sugiere que la adherencia a la matriz proteica extracelular es un mecanismo de persistencia intestinal. Dicha persistencia intestinal en animales clínicamente sanos, pone en riesgo de contaminación bacteriana las canales, así como las superficies y aditamentos de las plantas procesadoras de alimentos y por consiguiente del alimento destinado para consumo humano. Entender ampliamente los mecanismos involucrados en la persistencia intestinal, permitirá el desarrollo de estrategias innovadoras que salvaguarden la seguridad alimentaria a través de la reducción de la prevalencia de S. Typhimurium entre los animales domésticos destinados para la producción de alimentos (Kingsley et al., 2003).
Otras investigaciones asocian también la unión de S. Typhimurium a colágeno y fibronectina de la matriz extracelular y la colonización intestinal, involucrando de igual manera a la proteína ShdA (Kingsley et al., 2003; Wells et al., 2001). El agregado fimbrial codificado por el operon agf (csg) media la unión a fibronectina que se requiere para la persistencia de S. Typhimurium al epitelio del intestino delgado. La fimbria tipo I de S. Typhimurium se une a laminina y también está involucrada en la colonización intestinal (Kingsley et al., 2004).
Una segunda proteína de autotransporte de S.Typhimurium es codificada por el gen misl localizado en la isla de patogenicidad 3 (SPI3). Este gen se asocia a colonización intestinal crónica en modelo murino, incluso en modelos experimentales de aves se confirmó dicha función (Naughton et al., 2001). Además se ha identificado fibronectina como un ligando para adhesina MisL. Estos hallazgos soportan fuertemente la idea de que la unión a las proteínas de la matriz extracelular puede ser un punto clave durante la colonización intestinal por S. Typhimurium. Este complejo MisL- fibronectina unido a la superficie celular ocasiona un agregado bacteriano in vitro. En resumen, la posibilidad de que la unión de las proteínas de la matriz extracelular a través de MisL a las áreas de erosión epitelial, promovería el agregado bacteriano a estos sitios, y por consiguiente contribuirían a la colonización bacteriana persistente.
Un mecanismo adicional o alternativo es que la unión de fibronectina se dé en la superficie apical de las células epiteliales (Morgan et al., 2004). Los mecanismos que permiten a este patógeno persistir en el aparato digestivo de los animales, son poco conocidos. Las fimbrias codificadas por los genes lpf, bcf, stb, stc, std y operones fimbriales sth pudieran estar involucradas en la colonización por S. Typhimurium, ya que datos de investigaciones en modelos murinos así lo han demostrado (Walia et al., 2004; Weening et al., 2005), incluso se han identificado seis operones fimbriales (lpfabcde, bcfabcdefg, stbabcde, stcabcd, stdabcd y sthabcde) que contribuyen a facilitar la excreción bacteriana en heces y colonización del ciego en modelos experimentales a los 30 días post-infección. De tal manera que, por lo menos en el modelo murino, se requieren por lo menos de 32 genes de S. Typhimurium para la persistencia intestinal. Resulta conveniente considerar también el factor de susceptibilidad de especie. Es de esperarse diferencias sustanciales en la expresión de receptores en animales de diferentes especies en comparación con aquellas pertenecientes a una misma especie animal. Por ejemplo, la fimbria tipo 1 no contribuye a favorecer la persistencia intestinal en el ratón BALB/c o CBA, pero si es importante en la infección de cerdos y ratas (Kingsley etal., 2003).
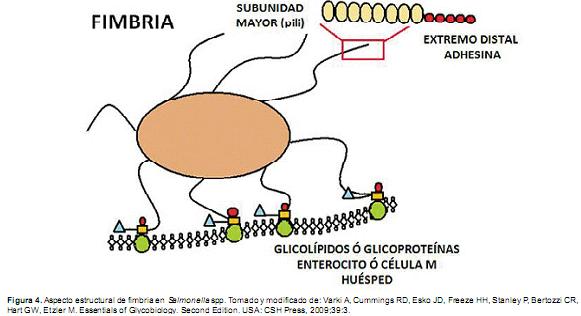
En el caso de las adhesinas, existe una amplia variabilidad requerida para la colonización en diferentes especies, podría especularse que el extenso agregado genético presente en el genoma de S. Typhimurium puede representar una adaptación para colonizar hospederos que difieran en el repertorio de receptores expresados sobre sus superficies mucosas. En el caso de la infección por S. enterica serovar Enteritidis en ratón, la gran mayoría de las bacterias residen en el lumen del ciego. En mutaciones genéticas de shda y ratb, que ocasionan la disminución de la colonización por S.Typhimurium en colón, se ha observado que el número de bacterias recuperadas de heces también se redujo. En contraste, una mutación en el gen sivhreduce la colonización en las placas de Peyer pero no afecta la habilidad de S. Typhimurium de colonizar el ciego ó ser excretada en heces de ratón (Althouse et al., 2003). Estos hallazgos han sugerido que el ciego del modelo murino puede ser considerado como el principal reservorio bacteriano implicado en la excreción fecal de los diferentes serotipos de Salmonella, es decir, puede considerarse este órgano digestivo como un reservorio que brinda la oportunidad de aumentar el número de colonias bacterianas y ser excretadas junto con las heces. En el caso de la proteína de autotransporte ShdA, el mecanismo de colonización del ciego murino se explica de la siguiente manera. El dominio de unión de superficie de ShdA con el tejido cecal se lleva a cabo en áreas ricas en proteínas de matriz extracelular (fibronectina y Colágeno). S. Typhimurium codifica una segunda adhesina que se une a la fibronectina, complejo que se conoce como agregado fimbrial, el cual es codificado por el conjunto de genes fimbriales agf (csg) (Figura 4) (Kingsley et al., 2003).
Regiones zir. Los marcos abiertos de lectura (siglas ORFs del inglés Open reading frames) y designados STM1668 (zirs), están localizados 26 pares de bases río abajo de un "pseudogen- invasivo" denominado stm1669 (zirt). La región zirS fue adquirida principalmente a través de transferencia horizontal de genes durante el proceso evolutivo del género Salmonella, presenta una organización altamente conservada y se manifiesta en todas las secuencias genómicas de las serovariedades de Salmonella, incluyendo S. bongori. Se sabe que el sistema de secreción zirts es funcionalmente similar a otras vías de secreción, sin embargo representa una vía distintiva en el género Salmonella. Los resultados obtenidos en modelosde expresión in vivo sugieren que la expresión de estos genes se da principalmente en el tracto gastrointestinal, más que a nivel sistémico. Por ejemplo, se ha visto diferencia de excreción de Salmonella(medido en unidades formadoras de colonias UFC) en heces asociado a la expresión de zirt, de tal manera que cuando se excretan bajos niveles de bacterias, zirt no se expresa y, existe expresión cuando hay altos niveles de excreción en heces, lo que fuertemente orienta hacia la expresión de zirt en heces de ratones excretores. Análisis de secuencias de aminoácidos en zirs and zirt han mostrado diferencia con otros sistemas de secreción. La mayoría de las investigaciones en la patogénesis bacteriana están principalmente dirigidas al hallazgo de genes que promueven la virulencia en el hospedero. Sin embargo, estudios recientes han demostrado que el "equilibrio" que se presenta durante la infección entre el hospedero y el agente patógeno, se debe a una serie de genes conocidos como "genes antivirulentos" (Kingsley et al., 2004). En resumen, estas investigaciones contribuyen a enfatizar en el tema de patogenia, sobre cómo los agentes patógenos limitan sus efectos sobre las células que infectan y de esta manera, se mantiene un balance con el hospedero. Debemos tener en mentela posibilidad de que el incremento en el número bacteriano en los tejidos del hospedero pueda estar asociado a la activación o inactivación de genes. Pudiera pensarse por ejemplo, que el incremento en la carga bacteriana en el modelo murino, causaría septicemia y por consiguiente un choque séptico, sin embargo, esta letalidad disminuiría por lo tanto la transmisión del patógeno.
En el caso de S. Typhimurium,que primariamente se trasmite a través de la vía oro-fecal, en los animales infectados sucede la excreción bacteriana vía fecal, lo que garantizaría de esta manera tener acceso a otro hospedero y comenzar nuevamente un nuevo ciclo de infección. De tal manera que surge la hipótesis del papel que juega zirts como "modulador de virulencia" en los estadios tempranos de la infección, probablemente contribuya a equilibrar un balance después de la transmisión del patógeno al hospedero. Habrá que considerar por lo tanto el papel que juega zirts durante los estadios tempranos de la infección como mecanismo preventivo de muerte prematura en el hospedero (Formenan-Wykert y Miller, 2003).
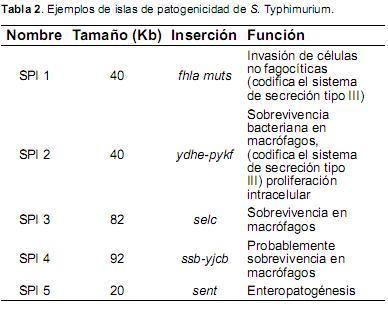
SPIs.Actualmente se han estado aclarando los factores genéticos utilizados por Salmonella tanto para causar alteración digestiva, así como infección sistémica. Para colonizar satisfactoriamente diferentes hospederos requiere una gran variedad genética, misma que se ubica en una parte específica del cromosoma bacteriano denominada isla de patogenicidad de Salmonella (SPIs del inglés Salmonella Pathogenicity Islands). Aunque existen más de 14 diferentes SPIs, la presencia de cada una de ellas varía dependiendo del serovar de Salmonella, sin embargo, cinco de ellas son comunes en el cromosoma de todos los serovares de S. enterica (Karasova et al., 2010). Las SPIs, denominadas con los números ordinales desde el 1 hasta el 14, han mostrado su papel esencial en la virulencia de Salmonella, incluyendo la penetración de enterocitos y la sobrevivencia en macrófagos (Tabla 2) (Gal-Mor et al., 2008).
]]>
Estas SPIs pueden producir un amplio rango de enfermedad en un gran número de especies animales. Otro aspecto importante en la infección por Salmonella es que la persistencia de la infección en animales portadores asintomáticos permite la contaminación de alimentos de origen animal para consumo humano.
Por ejemplo, el ganado porcino es una especie que puede presentar Salmonella, pero en la mayoría de los casos no muestran sintomatología clínica, en algunos casos se han llegado a aislar e identificar hasta más de seis serotipos en animales clínicamente normales en una sola unidad pecuaria (Davies et al., 1998). Para sobrevivir y persistir en los animales, el género Salmonella debe coordinar la expresión genética en respuesta a una gran variedad de ambientes durante el proceso de infección. Muy poco se conoce acerca de los factores genéticos requeridos en animales portadores clínicamente sanos y con excreción de bacterias en heces. De cualquier manera, los factores de colonización deben ser hospedero específico (Chan et al., 2005).
Además de los factores genéticos, una gran variedad de condiciones ambientales presentes en los hospederos animales, han mostrado que proveen señales que controlan la expresión genética. En etapas tempranas de la infección, es necesaria la penetración bacteriana en el epitelio intestinal, muchos de los genes requeridos para esto, se localizan en la SPI1. Se ha demostrado que la regulación de los genes involucrados, responden a una gran variedad de señales ambientales. La baja tensión de oxígeno, la alta osmolaridad y las condiciones en el íleon, han sido asociadas a la inducción de los genes de invasión localizados en la SPI1. La transcripción de los genes de invasividad puede ser también reprimida por la bilis.
Las condiciones ambientales presentes en los animales pueden proveer señales plausibles necesarias para otras funciones requeridas por Salmonella(Carnell et al., 2007). Un factor asociado en portadores asintomáticos ha sido designado como ZraS/ZraR (sensor asociado a la resistencia/ regulación del zinc) (Lawhon et al., 2003). El requerimiento en la dieta de zinc en los cerdos es de 50 a 100 ppm, mucho más que cualquier otra especie doméstica (Berger, 1987). Estas altas concentraciones de zinc son detectadas a través de ZraS/ZraR lo cual favorece la expresión de genes tales como hydh ó zrap tanto en intestino como en tonsilas cecales favoreciendo la vida bacteriana en los hospederos animales. Otro gen identificado es ycir, también designado gcpe (GGDEF dominio- conteniendo proteína E) (Leonhartsberger et al., 2001). Estas proteínas controlan la concentración intracelular del segundo mensajero cdi-GMP y junto con el dominio GGDEF estimulan la producción y degradación del dominio cdi-GMP (García et al., 2004). cdi-GMP ha sido identificado como un responsable regulador global para la motilidad, adhesión, virulencia y formación de biopelículas (Simm et al., 2004). El operón hscba- fdx, importante en la colonización del serovar Typhimurium en portadores sanos y ubicado en la SPI 1, codifica una proteína de 66-kDa homóloga de una proteína DnaK, dichas proteínas favorecen la sobrevivencia de la bacteria en los macrófagos así como la unión con la mucosa intestinal (Mendez- Ortiz et al., 2005)
Por otra parte, aunque los compuestos aromáticos son altamente abundantes en suelo y agua, se ha visto que existen algunas fuentes de estos compuestos en el tracto gastrointestinal, la mayoría derivados de la fermentación de aminoácidos aromáticos provenientes de fuentes vegetales. Se ha visto que el serovar Typhimurium es capaz de degradar ciertos compuestos aromáticos cuando viven en hospederos animales, lo que muestra su habilidad para colonizar el intestino. La vitamina B12 se requiere para la degradación de etanolamina y propanediol, ambos procedentes de fuentes de carbón presentes en el tracto gastrointestinal. Por lo tanto, es posible que Salmonella induzca al operon cob, para que se utilicen estas fuentes nutritivas mientras vivan en el tracto intestinal. Incluso se ha observado que la invasión a los enterocitos se co-regula con el metabolismo de propanediol y etanolamina. Estos son solo algunos ejemplos que muestran puntos ambientales claves que regulan importantes funciones en el hospederos animal, tanto metabólicas como de virulencia (Hoff et al., 2002).
Estas investigaciones aportan una nueva visión genética acerca de la sobrevivencia y colonización de Salmonella en animales portadores que no presentan sintomatología clínica, lo que en un futuro, permitirá establecer nuevas terapias o bien estrategias de prevención que reduzcan la contaminación del alimento de origen animal para consumo humano (Lawhon et al., 2003).
Microbiota intestinal y su papel en la persistencia bacteriana. especies de bacterias controlan la expresión de diversos genes en comunidades de microorganismos a través de la producción, secreción y detección de moléculas de señalización extracelular, conocidas como autoinductores (AIs), que se acumulan en el ambiente en relación a la densidad celular. Este proceso es denominado "quorum sensing" (QS) y se dispara cuando la señal alcanza una concentración umbral estimuladora. Cada una de estas respuestas permite a la comunidad de individuos adaptarse mejor a su entorno y en el caso específico de los patógenos, expresar genes de virulencia para persistir y diseminarse en su hospedero. (Jones et al., 2003; Xavier et al., 2003).
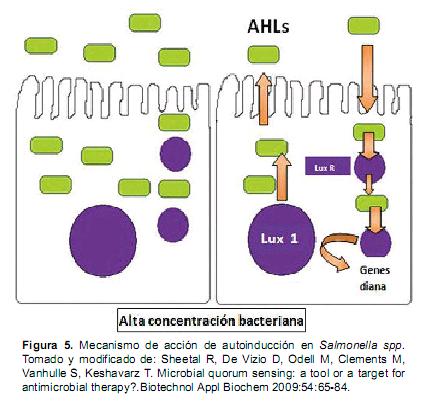
Se han identificado genes de S. Typhimurium que se expresan específicamente ante la presencia de moléculas señal producidas por otros microbios. Entre estas moléculas AIs se encuentran las acil homoserina lactonas (AHLs), su producción depende de las LuxI sintetasas y su detección de activadores transcripcionales del tipo LuxR que unen autoinductores (Figura 5). La función típica de estas proteínas LuxR es "detección de quórum o autoinducción", mecanismo de control de expresiones genéticas dependiente de la densidad celular (Waters y Bassler, 2005). Numerosos patógenos tanto de plantas como de animales utilizan la autoinducción para regular la interacción genética con el hospedero. En el caso de Salmonella esperan a "reunir quorum" antes de liberar las toxinas que atacaran a su huésped. Si la bacteria ataca sola, en vez de hacerlo en conjunto, el sistema inmunológico la neutralizaría rápidamente. Las AHLs son sintetizadas usando enzimas de la familia de LuxI ó LuxM (Milton et al., 2001). Salmonella codifica el homólogo LuxR, SdiA, pero no codifica una sintasa AHL. Por ejemplo, SdiA responde a la producción de AHLs por otras especies bacterianas (Ahmer, 2004).
La composición intestinal de los mamíferos contiene aproximadamente de 800 especies microbianas. Se piensa que esta microflora bacteriana utiliza la autoinducción tipo AHLs y que la presencia de estos AHLs provee a S. Typhimurium una manera efectiva de detectar el ambiente intestinal. En otras investigaciones, se ha visto que S. Typhimurium no detecta AHLs durante el tránsito a través del tracto intestinal de muchas especies animales, lo que sugiere que la microbiota normal de estas especies no produce AHLs del tipo correcto, o bien no en la cantidad suficiente para activar la autoinducción (Backhed et al., 2005). Se ha determinado que SdiA de S. Typhimurium se activa en tortugas, pero no en ratones, cerdos, gallinas, vacas, conejos y cuyes. Parece que la microbiota normal de estos animales no produce AHLs, por lo menos no en la localización ni en la concentración suficiente para activar SdiA. Tomando en cuenta que S. Typhimurium no es un serovar que se asocia comúnmente con tortugas, en estos casos puede ser que S. Typhimurium no posee genes reguladores de SdiA en estas especies animales. Resultaría interesante determinar qué sucedería en tortugas con mutación SdiA asociadas a serovar Muenchen, Arizonae, o Newport, probablemente SdiA confiere una ventaja conveniente en cualquiera de éstas situaciones de coinfección (Smith et al., 2008).
]]>
Medio ambiente y persistencia bacteriana. Las "Thin aggregative fimbriae (Tafi)" son estructuras superficiales de naturaleza proteica consideradas como adhesinas que facilitan la unión de la bacteria a receptores específicos de las células del hospedador (fimbrias). Estas estructuras son producidas junto con celulosa (White et al., 2003), así como también con polisacáridos adicionales (Gibson et al., 2006) y proteínas (BapA) para formar una matriz extracelular que enlaza a las células. El fenotipo asociado con la producción de la matriz se ha denominado morfotipo rdar (red, dry and rough). El Control regulador de rdar se enfoca primariamente al nivel de expresión de AgfD (CsgD), una proteína transcripcional reguladora. AgfD activa la producción de Tafi y polímeros de celulosa, lo que permite a las células responder rápidamente a los desafíos ambientales. La di- guanosina monofosfato cíclico (c-di-GMP), se ha involucrado en la producción de celulosa bacteriana, así como en la modificación de la superficie celular que le permite regular la formación de biopelículas, la motilidad y la virulencia entre otros procesos bacterianos.
Algunos de los efectos del c-di- GMP en el control del morfotipo rdar es específico en Salmonella. En la mayoría de los aislamientos de Salmonella, el morfotipo rdar se expresa bajo condiciones de baja osmolaridad, limitación nutricional y temperatura por debajo de los 30°C (Gerstel et al., 2001). El morfotipo rdar es importante para la persistencia y sobrevivencia de Salmonella en ambientes fuera del hospedero (White et al., 2006), y su producción se asocia directamente a los genes agfd, agfb, adra, y yihu involucrados también en la síntesis de celulosa y antígeno capsular O. Las bacterias con expresión de morfotipo rdar resisten hasta 60 ppm de hipoclorito de sodio, comparadas con aquellas no expresadas, lo que se sugiere que dichos grupos de bacterias producen celulosa, lo que les permite resistir la acción del desinfectante, así como la desecación, y de esta manera la sobrevivencia de Salmonella en medios ambientes naturales puede verse favorecida (White et al., 2006).
En algunos casos de infección, la bacteria no se disemina sistémicamente, lo que pudiera atribuirse a que la fagocitosis es inhibida por la presencia de una matriz extracelular y al no ser fagocitada, se limita al tracto intestinal, además de que la no expresión de agfd y agfb indique la ausencia de señal ó presencia de una condición o factor inhibidor que prevenga la expresión de agfb dentro del hospedero (Weber et al., 2006).
La expresión del morfotipo rdar asegura que más bacterias sobrevivan fuera del hospedero y conserven su capacidad infectiva, lo que explicaría porque este fenotipo se ha conservado a lo largo de la evolución (White et al., 2006). Si el rol primario del enlace ó agregación bacteriana en el ciclo de vida de Salmonella, es con el objetivo de su sobrevivencia en condiciones adversas, la pérdida del morfotipo rdar podría explicar las diferencias observadas en los modos de transmisión de los diferentes serovares (White et al., 2008).
Conclusiones
Las investigaciones sobre los procesos inmunológicos y genéticos de Salmonella, versan generalmente sobre el serovar Typhimurium, ya que el proceso infeccioso desarrollado en modelo murino, representa la salmonelosis tífica en el ser humano, y aunque existe mucho camino recorrido, los mecanismos moleculares de este proceso recién comienzan a ser entendidos. Sin embargo, la diversidad de serovariedades del género Salmonella, exige el desarrollo de investigaciones, desde el punto de vista básico, que analicen las particularidades genómicas en cada una de las serovariedades pertenecientes a este género, para estar en posibilidades de esclarecer el comportamiento bacteriano en los diferentes hospederos animales y, prevenir brotes potenciales en la población humana, ya que se trata de una enfermedad de índole zoonótica. El control de esta zoonosis tiene una alta prioridad ya que la situación epidemiológica gradualmente ha empeorado y el predominio de infección de Salmonellaha aumentado notablemente tanto en gente como en animales domésticos. Aunado a esto, la presentación más frecuente de la enfermedad es en forma de portador asintomático, un reto más que vencer por los profesionistas del área médica, la capacidad de Salmonella para sobrevivir en muchos lugares ambientales sugiere que su diseminación probablemente siga en aumento en el futuro.
Existe aún todo un camino por recorrer y descubrir. Los trabajos de investigación desarrollados hasta ahora, pierden de vista las disimilitudes existentes entre los ratones de experimentación y los seres humanos, si bien, han aportado grandes avances en la dinámica bacteriana, falta considerar las adaptaciones bacterianas de serovares hospedero- específicos a hospederos no específicos, más aún, analizar la dinámica espacio temporal bacteriana de más de un serovar que cohabita en un mismo animal ó bien en el medio ambiente. Estamos en una época de resurgimiento de enfermedades infecciosas emergentes y reemergentes, sin embargo estamos también en una época donde se cuenta con avanzadas técnicas de biología molecular y con modelos matemáticos y simulaciones con ordenador, para estudiar la dinámica de transmisión y control de las enfermedades infecciosas, que impacten en la investigación básica y por consiguiente en la investigación aplicada, y de esta manera estar en condiciones de reducir significativamente los casos de salmonelosis en las diferentes regiones del mundo.
Referencias
]]>1. Ahmer BM. Cell-to-cell signalling in Escherichia coli and Salmonellaenterica. Mol Microbiol 2004; 52:933-945. [ Links ]
2. Althouse C, Patterson S, Fedorka-Cray P, Isaacson RE. Type 1 fimbriae of Salmonella enterica serovar Typhimurium bind to enterocytes and contribute to colonization of swine in vivo. Infect Immun 2003; 71:6446-6452. [ Links ]
3. Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host bacterial mutualism in the human intestine. Science 2005; 307:1915-1920. [ Links ]
4. Berger LL. Salt and trace minerals for livestock, poultry and other anximals. Salt Institute, Alexandria, VA. 1987. [ Links ]
5. Carnell S, Bowen A, Morgan E, Maskell DJ, Wallis TS, Stevens MP. Role in virulence and protective efficacy in pigs of Salmonella enterica serovar Typhimurium secreted components identified by signature tagged mutagenesis. Microbiology 2007; 153:1940-1952. [ Links ]
6. Chan K, Kim CC, Falkow S. Microarray-based detection of Salmonella enterica serovar Typhimurium transposon mutants that cannot survive in macrophages and mice. Infect Immun 2005; 73:5438-5449. [ Links ]
7. Crump JA, Luby SP, Mintz ED. The global burden of typhoid fever. Bull World Health Organ 2004; 82:346-353. [ Links ]
8. Dann SM, Eckmann L. Innate immune defenses in the intestinal tract. Curr Opin Gastroenterol 2007; 23:115-120. [ Links ]
9. Davies PR, Geradus F, Bovee EM, Funk JA, Morrow WEM, Jones FT, Deen J. Isolation of Salmonella serotypes from feces of pigs raised in a multiple-site production system. J Am Vet Med Assoc 1998; 212:1925-1929. [ Links ]
10. Davies RH, Dalziel R, Gibbens JC, Wilesmith JW, Ryan JM, Evans SJ, Byrne C, Paiba GA, Pascoe SJ, Teale CJ. National survey for Salmonella in pigs, cattle and sheep at slaughter in Great Britain (1999-2000). J Appl Microbiol 2004; 96:750-760. [ Links ]
11. Drecktrah D, Knodler LA, Galbraith K, Steele-Mortimer O. The Salmonella SPI1 effector SopB stimulates nitric oxide production long after invasion. Cell Microbiol 2005; 7:105-113. [ Links ]
12. Fink SL, Cookson BT. Apoptosis, pyroptosis, and necrosis: mechanistic description of dead and dying eukaryotic cells. Infect Immun 2005; 73:1907-1916. [ Links ]
13. Foreman-Wykert AK, Miller JF. Hypervirulence and pathogen fitness. Trends Microbiol 2003; 11:105-108. [ Links ]
14. Gal-Mor O, Gibson DL, Baluta D, Vallance BA, Finlay BB. A novel secretion pathway of Salmonella enterica acts as an antivirulence modulator during Salmonellosis. PLoS Pathog 2008; [Agosto 2009] URL: http://dx.plos.org/10.1371/journal.ppat.1000036 [ Links ]
15. García B, Latasa C, Solano C, García-del Portillo F, Gamazo C, Lasa I. Role of the GGDEF protein family in Salmonella cellulose biosynthesis and biofilm formation. Mol Microbiol 2004; 54:264-277. [ Links ]
16. Gerstel U, Romling U. Oxygen tension and nutrient starvation are major signals that regulate agfD promoter activity and expression of the multicellular morphotype in Salmonella Typhimurium. Environ Microbiol 2001; 3:638-648. [ Links ]
17. Gibson DL, White AP, Snyder SD, Martin S, Heiss C, Azadi P, Surette MG, Kay WW. Salmonella produces an O-antigen capsule regulated by AgfD and important for environmental persistence. J Bacteriol 2006; 188:7722-7730. [ Links ]
18. Grenfell BT, Pybus OG, Gog JR, Wood JLN, Daly JM, Mumford JA, Holmes EC. Unifying the epidemiological and evolutionary dynamics of pathogens. Science 2004; 303:327- 332. [ Links ]
19. Henderson IR, Cappello R, Nataro JP. Autotransporter proteins, evolution and redefining protein secretion. Trends Microbiol 2000; 8:529-532. [ Links ]
20. Hoff KG, Ta DT, Tapley TL, Silberg JJ, Vickery LE. Hsc66 substrate specificity is directed toward a discrete region of the iron-sulfur cluster template protein IscU. J Biol Chem 2002; 277:27353-27359. [ Links ]
21. Jones M, Blaser M. Detection of a luxS-signaling molecule in Bacillusanthracis.Infect Immun 2003; 71:3914-3919. [ Links ]
22. Kankwatira AM, Mwafulirwa GA, Gordon MA. Non typhoidal Salmonellabacteremia an underrecognized feature of AIDS in African adults. Trop Doct 2004; 34:198-200. [ Links ]
23. Karasova D, Sebkova A, Havlickova H, Sisak F, Volf J, Faldyna M, Ondrackova P, Kummer V, Rychlik I. Influence of 5 major Salmonellapathogenicity islands on NK cell depletion in mice infected with Salmonella enterica serovar Enteritidis. BMC Microbiology 2010; 10:75. [ Links ]
24. Kingsley RA, Humphries AD, Weening EH, De Zoete MR, Winter S, Papaconstantinopoulou A. Molecular and phenotypic analysis of the CS54 island of Salmonella enterica serotype Typhimurium: identification of intestinal colonization and persistence determinants. Infect Immun 2003; 71:629-640. [ Links ]
25. Kingsley RA, Keestra AM, de Zoete MR, Bäumler AJ. The ShdA adhesin binds to the cationic cradle of the fibronectin 13FnIII repeat module: evidence for molecular mimicry of heparin binding. Mol Microbiol 2004; 52:345-355. [ Links ]
26. Kingsley RA, Santos RL, Keestra AM, Adams LG, Bäumler AJ. Salmonella enterica serotype Typhimurium ShdA is an outer membrane fibronectin-binding protein that is expressed in the intestine. Mol Microbiol 2002; 43:895-905. [ Links ]
27. Lawhon SD, Frye JG, Suyemoto M, Porwollik S, McClelland M, Altier C. Global regulation by CsrA in Salmonella Typhimurium. Mol Microbiol 2003; 48:1633-1645. [ Links ]
28. Lawley TD, Chan K, Thompson LJ, Kim CC, Govoni GR, Monack DM. Genome-wide screen for Salmonella genes required for longterm systemic infection of the mouse. PLoS Pathog 2006; [Agosto 2009] URL: http://dx.plos.org/10.1371/journal.ppat.0020011 [ Links ]
29. Leonhartsberger S, Huber A, Lottspeich F, Bock A. The hydH/G genes from Escherichia coli code for a zinc and lead responsive two-component regulatory system. J Mol Biol 2001; 307:93- 105. [ Links ]
30. Maclennan CA, Gondwe EN, Msefula CL, Kingsley RA, Thomson NR, White SA, Goodall M, Pickard DJ, Graham SM, Dougan G, Hart CA, Molyneux ME, Drayson MT. The neglected role of antibody in protection against bacteremia caused by nontyphoidal strains of Salmonella in African children. J Clin Invest 2008; 118:1553-1562. [ Links ]
31. Mastroeni P, Villarreal-Ramos B, Hormaeche CE. Role of T cells, TNF alpha and IFN gamma in recall of immunity to oral challenge with virulent salmonellae in mice vaccinated with live attenuated aro- Salmonella vaccines. Microb Pathog 1992; 13:477-491. [ Links ]
32. Mendez-Ortiz MM, Membrillo-Hernández J. Proteins with GGDEF and EAL domains: their role in bacterial metabolism. Rev Latinoam Microbiol 2005; 47:130-139. [ Links ]
33. Milton DL, Chalker VJ, Kirke D, Hardman A, Camara M. The LuxM homologue VanM from Vibrio anguillarum directs the synthesis of N-(3- hydroxyhexanoyl) homoserine lactone and N-hexanoylhomoserine lactone. J Bacteriol 2001; 183:3537- 3547. [ Links ]
34. Monack DM, Bouley DM, Falkow S. SalmonellaTyphimurium persists within macrophages in the mesenteric lymph nodes of chronically infected nramp1 mice and can be reactivated by IFN neutralization. J Exp Med 2004; 199:231-241. [ Links ]
35. Morgan E, Campbell JD, Rowe SC, Bispham J, Stevens MP, Bowen AJ, Barrow PA, Maskell DJ, Wallis TS. Identification of host-specific colonization factors of Salmonella enterica serovar Typhimurium. Mol Microbiol 2004; 54:994-1010. [ Links ]
36. Muotiala A, Makela PH. The role of IFN-gamma in murine SalmonellaTyphimurium infection. Microb Pathog 1990; 8:135-141. [ Links ]
37. Naughton PJ, Grant G, Bardocz S, Allen-Vercoe E, Woodward MJ, Pusztai A. Expression of type 1 fimbriae (SEF 21) of Salmonella enterica serotype Enteritidis in the early colonization of the rat intestine. J Med Microbiol 2001; 50:191- 197. [ Links ]
38. Nix RN, Altschuler SE, Henson PM, Detweiler CS. Hemophagocytic macrophages harbor Salmonella enterica during persistent infection. PLoS Pathog 2007; [Julio 2009] URL: http://dx.plos.org/10.1371/journal.ppat.0030193. [ Links ]
39. Paulin SM, Aparna J, Campbell J, Wallis T, Stevens M. Net replication of Salmonella enterica serovars Typhimurium and Choleraesuis in porcine intestinal mucosa and nodes is associated with their differential virulence. Infect Immun 2007; 75:3950-3960. [ Links ]
40. Paulin SM, Watson PR, Benmore AR, Stevens MP, Jones PW, Villarreal-Ramos B, Wallis TS. Analysis of Salmonella enteric serotype-host specificity in calves: avirulence of S. enterica serotype Gallinarum correlates with bacterial dissemination from mesenteric lymph nodes and persistence in vivo. Infect Immun 2002; 70:6788-6797. [ Links ]
41. Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, Granucci F, Kraehenbuhl JP, Ricciardi-Castagnoli P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nature Immunol 2001; 2:361-367. [ Links ]
42. Simm R, Morr M, Kader A, Nimtz M, Romling U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol Microbiol 2004; 53:1123-1134. [ Links ]
43. Smith D, Wang JH, Swatton JE, Davenport P, Price B. Variations on a theme: diverse N-acyl homoserine lactone- mediated quorum sensing mechanisms in gram-negative bacteria. Sci Prog 2006; 89:167-211. [ Links ]
44. Smith JN, Dyszel JL, Soares JA, Ellermeier CD, Altier C. SdiA, an N-acylhomoserine lactone receptor, becomes active during the transit of Salmonella enterica through the gastrointestinal tract of turtles. PLoS ONE 2008; [Agosto 2009] URL: http://dx.doi.org/10.1371/journal.pone.0002826 [ Links ]
45. Sonnenburg JL, Xu J, Leip DD, Chen CH, Westover BP. Glycan foraging in vivo by an intestine-adapted bacterial symbiont. Science 2005; 307:1955-1959. [ Links ]
46. Stecher B, Robbiani R, Walker AW, Westendorf AM, Barthel M, Kremer M, Chaffron S, Macpherson AJ, Buer J, Parkhill J, Dougan G, von Mering C, Hardt WD. Salmonella enterica serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol 2007; [Septiembre 2009] URL: http://dx.doi.org/10.1371/journal.pbio.0050244 [ Links ]
47. Steele-Mortimer O, Brumell JH, Knodler LA, Meresse S, Lopez A, Finlay BB. The invasion-associated type III secretion system of Salmonella enterica serovar Typhimurium is necessary for intracellular proliferation and vacuole biogenesis in epithelial cells. Cell Microbiol 2002; 4:43-54. [ Links ]
48. Suzuki K, Meek B, Doi Y, Muramatsu M, Chiba T. Aberrant expansion of segmented filamentous bacteria in IgA-deficient gut. Proc Natl Acad Sci 2004; 101:1981-1986. [ Links ]
49. Uthe JJ, Royaee A, Lunney JK, Stabel TJ, Zhao SH, Tuggle CK, Bearson SM. Porcine differential gene expression in response to Salmonella enterica serovars Choleraesuis and Typhimurium. Mol Immunol 2007; 44:2900-2914. [ Links ]
50. Vázquez TA, Jones JC, Bäumler AJ, Falkow S, Valdivia R, Brown W, Le M, Berggren R, Parks WT, Fang FC. Extraintestinal dissemination of Salmonella by CD18- expressing phagocytes. Nature 1999; 401:804-808. [ Links ]
51. Walia B, Castaneda FE, Wang L, Kolachala VL, Bajaj R, Roman J, Merlin D, Gewirtz AT, Sitaraman SV. Polarized fibronectin secretion induced by adenosine regulates bacterial- epithelial interaction in human intestinal epithelial cells. Biochem J 2004; 382:589-596. [ Links ]
52. Warren J, Mastroeni P, Dougan G, Noursadeghi M, Cohen J, Walport MJ, Botto M. Increased susceptibility of C1q deficient mice to Salmonella enterica serovar Typhimurium infection. Infect Immun 2002; 70:551-557. [ Links ]
53. Waters CM, Bassler BL. Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol 2005; 21:319-346. [ Links ]
54. Weber H, Pesavento C, Possling A, Tischendorf G, Hengge R. Cyclic-di-GMP-mediated signaling within the sigma network of Escherichia coli. Mol Microbiol 2006; 62:1014-1034. [ Links ]
55. Weening EH, Barker JD, Laarakker MC, Humphries AD, Tsolis R, Ba¸mler AJ. A previous study identified two genes, shdA and ratB, located on the CS54 island of S. enterica serotype Typhimurium that are involved in long-term intestinal persistence in genetically resistant mice. Infect Immun 2005; 73:3358-3366. [ Links ]
56. Wells SJ, Fedorka-Cray PJ, Dargatz DA, Ferris K, Green A. Fecal shedding of Salmonella spp. by dairy cows on farm and at cull cow markets. J Food Protect 2001; 64:3-11. [ Links ]
57. White AP, Gibson DL, Grassl GA, Kay WW, Finlay BB, Vallance BA, Surette MG. Aggregation via the red, dry, and rough morphotype is not a virulence adaptation in Salmonellaentericaserovar Typhimurium. Infect Immun 2008; 76:1048- 1058. [ Links ]
58. White AP, Gibson DL, Collinson SK, Banser PA, Kay WW. Extracellular polysaccharides associated with thin aggregative fimbriae of Salmonella enterica serovar Enteritidis. J Bacteriol 2003; 185:5398-5407. [ Links ]
59. White AP, Gibson DL, Kim W, Kay WW, Surette MG. Thin aggregative fimbriae and cellulose enhance long-term survival and persistence of Salmonella. J Bacteriol 2006; 188:3219- 3227. [ Links ]
60. White AP, Surette MG. Comparative genetics of the rdar morphotype in Salmonella. J Bacteriol 2006; 188:8395-8406. [ Links ]
61. Winter SE, Thiennimitr P, Winter MG, Butler BP, Huseby DL, Crawford RW, Russell JM, Bevins CL, Adams LG, Tsolis RM, Roth JR; Bäumler AJ. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010; 467:426-429. [ Links ]
62. Xavier KB, Bassier K. LuxS quorum sensing: more than just a numbers game. Curr Opin Microbiol 2003; 6:191-197. [ Links ] ]]>