EVALUACIÓN DE DOS MEDIOS DE CULTIVO Y HEREDABILIDAD DE PRODUCTIVIDAD Y TIEMPO DE DESARROLLO PARA TRES MUTANTES DE Drosophila melanogaster (DROSOPHILIDAE)
FERNANDO DÍAZ-GONZÁLEZ1, MAYRA PIZARRO-LOAIZA1, MAURICIO RAMÍREZ-CASTRILLÓN1, YHERSON MOLINA-HENAO1, DIEGO SOLARTE-GARCÍA1, DANIELA BRAVO-GUERRERO1, ALEJANDRA HURTADO-GIRALDO1, HEIBER CÁRDENAS-HENAO2 , Biólogo M.Sc.
1Grupo de Estudio y Trabajo En Genética GETEG y estudiantes de pregrado en el programa de Biología, Universidad del Valle. Calle 13 Nº 100-00, Cali-Colombia.
2Profesor Asociado, Universidad del Valle, Cali, Valle, Colombia. geteg@univalle.edu.co - ferdiazfer@gmail.com
Presentado 22 de julio de 2007, aceptado 25 de enero de 2008, correcciones 15 de febrero de 2008.
RESUMEN
Con el objetivo de investigar el efecto del medio de cultivo en la productividad y tiempo de desarrollo huevo-adulto de una cepa silvestre y tres cepas mutantes (CyLv, vg, w) de Drosophila melanogaster, se examinaron dos tipos de medios: banano y naranja. Para esto se empleó un diseño con dos factores, medio de cultivo y tipo de cepa, para un total de ocho tratamientos con cinco repeticiones cada uno. Se obtuvo que la productividad y el tiempo de desarrollo dependen del medio de cultivo y el tipo de cepa, encontrándose mayor productividad en el medio de naranja. La cepa silvestre presentó la mayor productividad y el menor tiempo de desarrollo en los dos medios (α=0,05). El análisis genético evidenció una heredabilidad baja y una variación fenotípica debida en su mayor parte al componente de interacción genotipo-ambiente, lo que explica la diferencia en el patrón de productividad y tiempo de desarrollo entre medios de cultivo.
]]> Palabras clave: Drosophila melanogaster, heredabilidad, tiempo de desarrollo, productividad, mutantes.ABSTRACT
With the objective of investigating the effect of the culture media in the productivity and development time egg-adult of Drosophila melanogaster in +/+ and three mutants (CyLv, vg, w), two culture media: banana and orange, were evaluated. An experimental design with two factors: culture media and kind of flies-stock, were tested, for a total of eight treatments with five replicas each one. The productivity and development time depend on culture media and kind of flies-stock, and the biggest productivity was in the orange culture media. The +/+ presented the biggest productivity and lowest development time in both culture media (α=0.05). The genetic analysis showed a low heritability and the phenotypic variation was due, in a mayor part, to the component of the interaction genotype-environment that explains the difference in the patron of productivity and development time between culture media.
Key words: Drosophila melanogaster, heritability, development time, productivity, mutants.
INTRODUCCIÓN
Drosophila melanogaster es considerada una de las especies existentes más importantes para el estudio de la biología. Esto se debe principalmente a su facilidad de cultivo, corto tiempo de generación, grandes cantidades de descendencia, tamaño pequeño, bajo costo en su manejo y mantenimiento (Mora et al., 2000). Esta especie pertenece al orden Díptera, tiene una distribución cosmopolita, y al igual que todos los organismos holometábolos, el desarrollo ontogénico presenta varios estadios: huevo, larva, pupa, adulto (Alvarado, 2000). Se ha evidenciado que factores ambientales como la temperatura, la humedad relativa y el tipo de medio de cultivo, ejercen una influencia en la productividad y desarrollo de D. melanogaster (Alvarado, 2000). La influencia que ejerce el medio de cultivo se debe principalmente a sus características físico-químicas y microbiológicas, tales como el porcentaje de nutrientes, el pH y la interacción existente entre microorganismos como bacterias y levaduras (Balbín et al., 2000). Otro factor de gran influencia en la productividad es el genotipo de los individuos. Se han reportado diferencias marcadas, en términos tanto de productividad como de la viabilidad entre los diferentes mutantes utilizados en D. melanogaster (Bonnier y Jonson, 1957). Esto se debe a que en muchos mutantes se ven afectadas características estructurales y funcionales como el comportamiento de cortejo, la capacidad de cópula, la oviposición y la viabilidad tanto del esperma como de los diferentes estadios de la metamorfosis (Mora et al., 2000), que influencian la tasa de mortalidad, la viabilidad y/o el tiempo de desarrollo de la especie (Gluecksohn-Waelsch, 1953; Shearn et al., 1971).
Mutantes como white (w//w), vestigial (vg//vg) y Curly-Lobe-vermillion (CyL//+L v//v) se encuentran entre los más utilizados en la experimentación genética, por lo que son de suprema importancia los conocimientos en términos de productividad para estas mutaciones (Ramos, 1993). La mutación w fue la primera descubierta en la especie; presenta herencia ligada al sexo (Morgan, 1914) y supresión de un gen involucrado en la producción de pigmentos como los homocromos y pteridinas, lo que ocasiona el color blanco en los ojos (Polaczyk et al., 1998). La mutación vg exhibe una herencia autosómica recesiva y se caracteriza por mostrar alas reducidas a vestigios, por lo general, dispuestas en ángulo recto respecto al cuerpo. En cuanto al triple mutante, la mutación v, presenta ojos color bermellón y herencia recesiva ligada al sexo. La mutación L, ojos de forma lobulada, y la mutación Cy son autosómicas dominantes, ambas letales en homocigosis (Ramos, 1993; Wallace, 1965). El presente trabajo determinó la productividad de los mutantes white (w//w), vestigial (vg//vg) y Curly-Lobe-vermillion (CyL//+L v//v) de D. melanogaster en dos medios de cultivo diferentes (banano y naranja) a temperatura ambiente. Se estimó el número de adultos desarrollados para cada uno de los mutantes evaluados y la cepa silvestre, en cada uno de los dos medios de cultivo, con el fin de asociar la productividad con el tiempo de desarrollo (días) huevo-adulto. En este estudio se encontraron diferencias significativas respecto a la productividad de las cepas silvestres entre los dos tipos de medios de cultivo. Esto sugiere la posibilidad de que exista una interacción entre algunos genotipos mutantes de D. melanogaster y algunos de los medios ya evaluados como naranja y banano (Díaz et al., 2005).
MATERIALES Y MÉTODOS
COLONIA FUENTE
Para el montaje del experimento se utilizaron moscas provenientes de la colonia de Drosophila del grupo de estudio GETEG, ubicado en las instalaciones de la Universidad del Valle, sede Meléndez.
]]> PPREPARACIÓN DEL MEDIO (PROTOCOLO PARA 10 FRASCOS)Para la preparación del medio, inicialmente se esterilizaron todos los frascos. Posteriormente, se licuaron 379 g de fruta en 333 mL de agua destilada. Simultáneamente, se diluyeron en otro recipiente 2,667 g de ágar en 66 mL de agua destilada. Luego, se calentó el licuado en una estufa, mezclando constantemente durante el proceso; una vez en ebullición, se adicionó y mezcló el agar diluído, luego se retiró la mezcla del calor y se dejó enfriar durante algunos minutos. Después de esto se adicionaron 1 mL de nistatina y 2,667 mL de ácido propiónico, mezclando durante el proceso. Final-mente, se adicionaron 0,1 g de levadura, se mezclaron y se sirvieron 50 mL de medio en cada frasco hasta la solidificación del medio. Por último, el montaje del experimento se realizó a temperatura ambiente en una cámara previamente esterilizada ubicada en el espacio del grupo de estudio GETEG.
DISEÑO ESTADÍSTICO
Para el análisis estadístico de los datos se empleó un diseño de dos factores (Tabla 1). El primer factor estaba constituido por los tres tipos de mutantes y la cepa silvestre, y el segundo factor por el tipo de medio de cultivo, banano y naranja. La variable respuesta fue la productividad de la especie expresada como el número de adultos generado en un tiempo de quince días y el tiempo de desarrollo en días. Se realizaron ocho tratamientos y cada tratamiento constó de cinco repeticiones, cada una de éstas correspondía a un frasco con medio de cultivo, tres machos y tres hembras vírgenes de tres días de fase adulta. En primer lugar, se evaluaron los supuestos de normalidad por medio de la prueba de Kolmogorov-Smirnov, y la homogeneidad de varianzas con la prueba Q de Bartlett. Con base en las pruebas anteriores se utilizó un análisis de varianza paramétrico. Por último, se hizo una comparación de medias post-ANDEVA de Tukey. Todos estos análisis se realizaron con los programasestadísticos STATISTICA® 6.0 (2003) y SPSS 11.5 para Windows (2002).
ANÁLISIS GENÉTICO
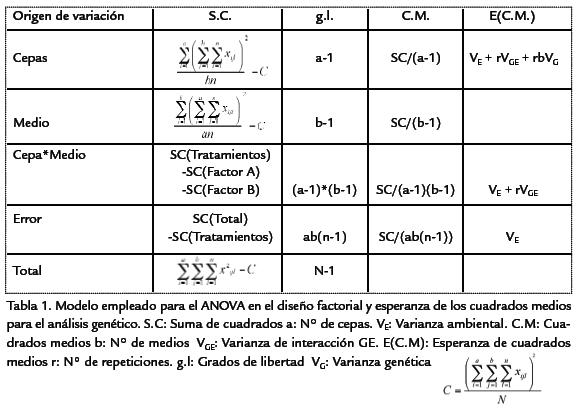
A partir de los cuadrados medios obtenidos en el análisis de varianza (ANDEVA; Tabla 1) se estimaron cada uno de los componentes de la varianza fenotípica obtenida parala productividad y el tiempo de desarrollo, discriminando la varianza genética, varian-za ambiental y la varianza de interacción genotipo-ambiente (Molina, 1992). De estaforma (Tabla 1),


Por último, se realizó un análisis de correlación lineal simple entre la productividad y el tiempo de desarrollo para determinar si la regulación génica y la interacción con el ambiente para estos dos caracteres de historia de vida se comparten de manera significativa.
TOMA DE DATOS
La toma de datos se inició ocho días después de la siembra de los parentales y su posterior sacrificio. Se realizó dos veces al día a las: 7:00 y 19:00 horas, registrando en cada una el estado del medio y el número de adultos emergidos que posteriormente se sacrificaron.
RESULTADOS
La productividad en los dos medios de cultivo, independientemente de las cepas evaluadas, presentó grandes diferencias (Figura 1), siendo el medio de banano el menos productivo con una media de 48.6 individuos respecto al de naranja con 125,5 individuos (Tabla 2; Figura 1). De igual manera ocurrió para el tiempo de desarrollo, un mayor tiempo para el medio de banano, con un tiempo medio de 9,055 días, mientras que el medio de naranja mostró una media de 6,996 días (Figura 2). Por otro lado, cada una de las cepas independientemente de los medios evaluados, presentaron grandes diferencias, encontrándose en el siguiente orden (Figura 1): la cepa w, de menor productividad con una media de 64,6 individuos adultos, seguida de CyLv con una productividad media de 67,4, después vg con 86,9 y el mayor número se observó en la cepa silvestre con una media de 127,7 individuos (Tabla 2; Fig. 1).
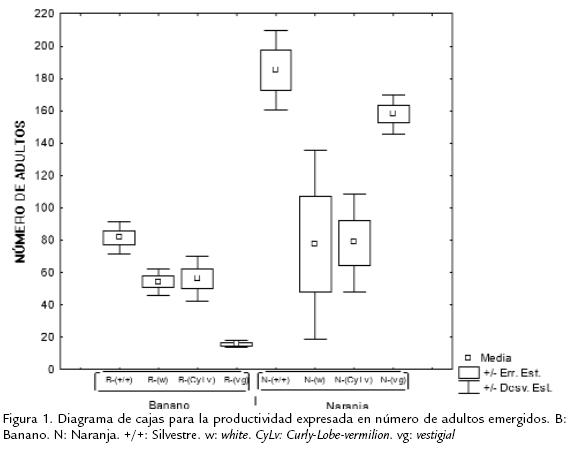
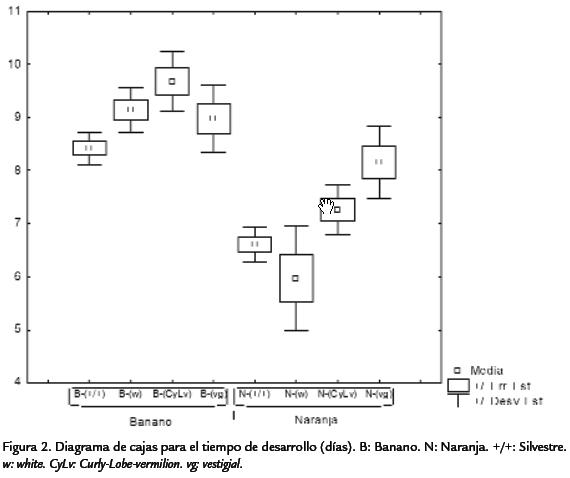
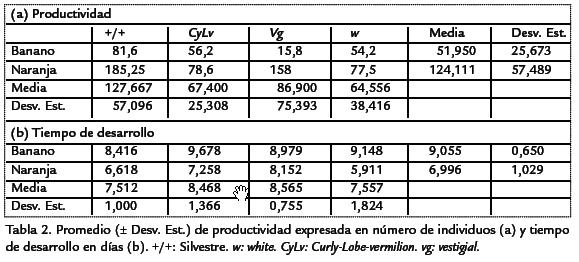
Ahora, los resultados para el tiempo de desarrollo no presentaron el mismo patrón (a excepción de la cepa silvestre; Figura 2). La cepa vg mostró un tiempo de desarrollo medio de 8,66 días, seguido de la cepa CyLv con 8,47, después la cepa w con 7,56, y por último, la cepa silvestre, con una valor medio de 7.5 días (Tabla 2; Figura 2). Es evidente que el medio de naranja muestra el mayor número medio de individuosadultos y menor tiempo medio de desarrollo (Figura 1; Figura 2). Mientras que las cepas,independientemente del medio de cultivo, no presentaron el mismo patrón. En térmi-nos de la variación entre repeticiones, la productividad expresada como el número deadultos presentó una variación mucho más alta (Fig. 1) que la del tiempo de desa-rrollo (Fig. 2). Sin embargo, ésta pueden ser evidencia de los grandes efectos de inte-racción de uno de los dos factores sobre el otro. Como ocurrió en el caso de la cepavestigialque fue diez veces más productiva en naranja que en banano. En general, lascurvas de los tiempos de desarrollo fueron similares. Así, la cantidad de individuos enambos medios mostraron varios picos ubicados entre el día tres y el 11 culminandocon un descenso posterior a este pico (Figura 3; Figura 4). Sin embargo, en el medio naran-ja la producción decrece a partir del día 12 y culmina en el día 14 (Figura 4). A diferenciade la producción en el medio de banano que no terminó en un descenso de indi-viduos, sino que continúo con una producción de individuos entre 0-30 casi constante hasta el fin del muestreo (Figura 4). Estas curvas de productividad evidenciaron las mismas diferencias obtenidas para los valores medios de productividad (Figura 1). Así, el área bajo cada una de las curvas, que representa la productividad para las cuatro cepas, es mayor en el medio de naranja (Figura 3). Lo mismo ocurre con los picos de máxima productividad, de tal manera que en el medio de banano, el pico de máxima productividad se observó en el día siete para la cepa silvestre, con un valor de 60 individuos, mucho menos incluso comparado con el pico obtenido para la cepa menos productiva (CyLv) del medio de naranja con 80 individuos (Fig. 3).
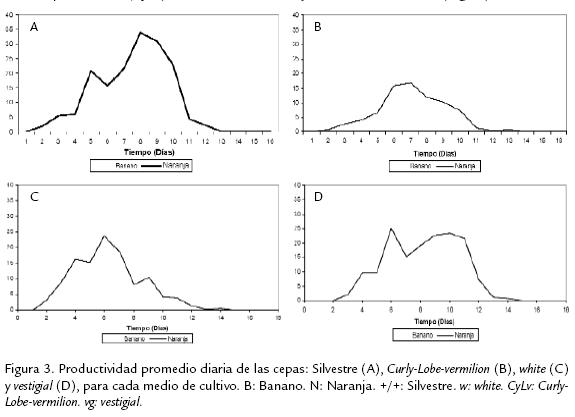
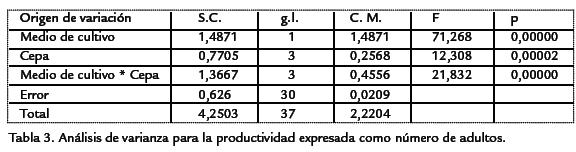
Los análisis de la productividad en el tiempo para cada una de las cepas evaluadas, evidenciaron el mismo patrón encontrado para los valores medios, teniendo la cepa silvestre una mayor área bajo la curva en ambos medios de cultivo, al igual que el mayor pico de máxima productividad (Fig. 4). Las siguientes cepas en orden descendente de productividad fueron w y CyLv con curvas muy similares (Fig. 4). De igual manera se evidencia una vez más la gran diferencia que presentó la cepa vg en términos de valores medios, es decir de evidencia la gran diferencia en productividad para cada uno de los medios de cultivo, siendo diez veces más productiva en naranja que en banano (Fig. 4; Tabla 2). Los datos de tiempo de desarrollo cumplieron con los supuestos del análisis de varianza, mientras que los de productividad no, razón por la cual fue necesario la eliminación de dos datos, una repetición del tratamiento Naran-ja-white y una de Naranja-(+/+). Además, estos datos se transformaron mediante la formula: Log(x) para el cumplimiento del supuesto de normalidad (p < 0,05), conforme a la prueba de Kolmogorov-Smirnov y homogeneidad (Q) de varianzas de Barlett (p< 0,05). El análisis de varianza bifactorial mostró diferencias significativasentre medios de cultivo y cepas, para la productividad y el tiempo de desarrollo (a=0,05), al igual que se encontraron efectos significativos de interacción (Tabla 3; Tabla 4). Evidentemente, el medio de cultivo es un factor que muestra diferencias signi-ficativas en términos de la productividad y el tiempo de desarrollo en D. melanogaster,siendo el medio de naranja el más productivo (Tabla 5; Tabla 6). Para identificar lasdiferencias entre cepas dentro de medios y entre medios para el criterio de número deadultos, se realizó la post-ANDEVA Tukey (a= 0,05). De este modo, la cepa silvestrefue la más productiva independientemente del medio de cultivo.
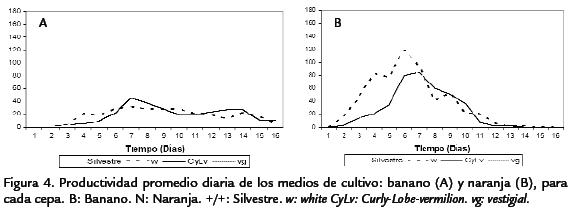
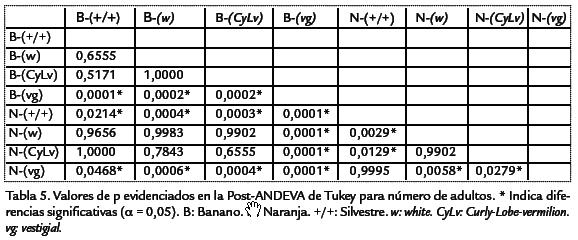
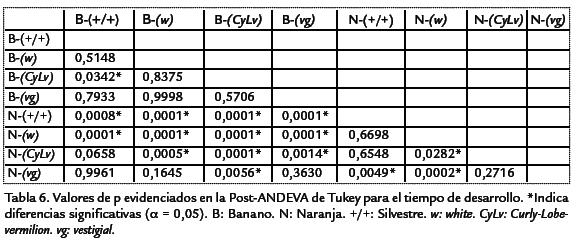
Por otro lado, para el medio de banano se encontraron diferencias significativas entre la cepa vg con la menor productividad respecto al resto (Tabla 5), siendo las cepas sil-vestre, wy CyLviguales en términos de productividad (Tabla 5). Al comparar las cepasdentro del medio de naranja el patrón de productividad fue diferente, lo que evidenciael efecto de interacción hallado a partir del análisis de varianza. Así, se encontró que lamayor productividad fue para las cepas vg y silvestre, presentando diferencias respectoa wy CyLv(Tabla 5). Ahora, al comparar las cepas entre los dos medios de cultivo se en-contraron diferencias significativas al comparar los tratamientos de naranja-silvestre,naranja-vgy banano-vgrespecto al resto de cepas. Así, los tratamientos de naranja-silvestre y naranja-vgpresentaron la mayor productividad, mientras que el tratamientode banano-silvestre evidenció la menor productividad. No obstante, al comparar lamisma cepa entre los medios de cultivo, se evidenció que el medio de cultivo afectó significativamente la productividad de las cepas silvestre y vg (siendo mayor su efecto enel medio de naranja), pero no la de wy CyLv(Tabla 5). Al realizar estas comparaciones para el tiempo de desarrollo, nuevamente silvestre fue la cepa con el menor tiempo dedesarrollo independientemente del medio de cultivo. En general, el tiempo de desarrollopresentó patrones de diferencias un poco distintos a los de productividad.
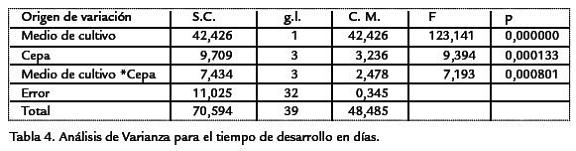
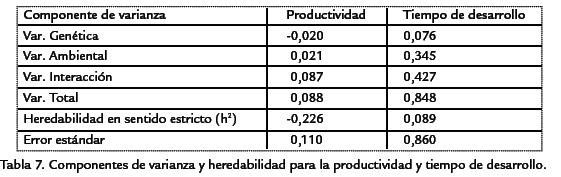
Así, dentro del medio de banano, solo se observaron diferencias significativas al compa-rar la cepa CyLvcon la cepa silvestre (Tabla 6. Dentro del medio de naranja la relaciónfue mucho más compleja, evidenciándose el efecto de interacción entre los dos factores,presentándose diferencias al comparar las cepas CyLvy vg,con las cepas wy silvestre, conmenor tiempo de desarrollo para estas dos últimas (Tabla 6). Por último, al comparartodas las cepas entre ambos medios de cultivo se observaron diferencias significativasentre la mayoría de parejas de comparaciones (Tabla 6). El análisis genético evidencióuna mayor varianza ambiental en comparación con la varianza genética. La productivi-dad presentó una varianza ambiental dos veces mayor a la genética, mientras que parael tiempo de desarrollo la varianza ambiental fue más de cuatro veces superior (Tabla 7). En ambos casos, la variación debida a la interacción genotipo-ambiente fue superiora la debida al genotipo o al ambiente por separado. Para la productividad, la varianzade interacción fue más de cuatro veces superior a las varianzas genética y ambiental,mientras que en el tiempo de desarrollo dicha varianza fue más de cinco veces superiora la varianza genética pero muy similar a la varianza ambiental. Debido a que las cepasutilizadas han sido mantenidas por cientos de generaciones en apareamientos endo-gámicos, cada una de ellas puede considerarse como una línea pura y por lo tanto, lavarianza genética hallada es enteramente la varianza aditiva. Por esta razón, la estima-ción de la heredabilidad corresponde a la heredabilidad en sentido estricto. Así, la pro-ductividad presentó una heredabilidad muy baja, cercana a cero, mientras que el tiempode desarrollo mostró un valor mayor, pero aun así en el rango considerado bajo (h2< 0,3). A pesar de valor negativo de la heredabilidad para la productividad, como el errorestándar es bajo (Tabla 7), se puede considerar que el valor es muy cercano a cero. Parael tiempo de desarrollo, el error estándar de la heredabilidad fue alto (Tabla 7), así paraeste carácter la estimación de heredabilidad fue poco significativa. El análisis de corre-lación entre la productividad y el tiempo de desarrollo mostró un valor de R2=0,0808,dicha correlación no fue significativamente diferente de cero (p > 0,0837).
]]>
DISCUSION
El efecto del medio encontrado en la productividad y tiempo de desarrollo de D.melanogasterindicó diferencias en los requerimientos nutritivos y características físico-químicas de los dos medios utilizados (Balbín et al., 2000). Estas diferencias causan un efecto significativo en estos caracteres y es claro que actúan en la totalidad del ciclo devida de D. melanogaster, como en la oviposición y principalmente en el desarrollo de lostres estadios larvales. Esto se debe a que es en la fase larvaria en la que ocurre la mayorinteracción de los individuos con el medio (Godoy, 2001). Al comparar la larva con eladulto, ésta se encuentra más restringida en sus movimientos por el sustrato donde sedesarrolla. Las larvas permanecen constantemente excavando el sustrato, ingiriendo eneste proceso el medio de cultivo y construyendo galerías (Godoy, 2001). Además, lamovilización de la larva en el medio de cultivo tiene un costo energético enorme, supe-rior a caminar, volar o nadar. Cuando la larva se desplaza por el medio se oponen fuer-zas de fricción que se manifiestan en un gran gasto de energía (Godoy, 2001). Entre las24 y las 96 horas de desarrollo, la larva de D. melanogasteringiere alimento con unagran voracidad. En este periodo la larva debe alcanzar un peso mínimo teórico que lepermita realizar la pupación y dar origen al adulto. La incapacidad para alcanzar estepeso mínimo, prolonga la etapa de alimentación y la expone a desaparecer porinanición, contaminación del lugar donde crece por excretas que ella misma produce,y/o parasitismo. Del mismo modo, la pupación puede llegar a iniciarse, pero por faltade reservas suficientes la transición pupa-adulto puede no tener lugar en muchasocasiones (Godoy, 2001). Debido a esto, los medios de cultivo que proporcionenmejores características tanto nutritivas como físico-químicas producen un mayornúmero de larvas que logran el desarrollo del correspondiente adulto, permitiendo deesta manera explicar las diferencias encontradas en el análisis de la varianza de laproductividad de los medios de cultivo.
Ahora, para determinar las razones por las cuales un medio resultó mejor que otro,es preciso aclarar más a fondo cuales son las características nutritivas que hacen a unmedio influir en el desarrollo de D. melanogaster. Es decir, exactamente de que com-ponentes del medio se alimentan tanto larvas como adultos. En general, las larvas sealimentan del medio en grandes cantidades. Sin embargo, la levadura es particular-mente nutritiva para su crecimiento y desarrollo, debido principalmente a que es unagran fuente de proteína (Ramos, 1993). Por otro lado, las moscas adultas, sealimentan principalmente de ácido acético (vinagre; Ramos, 1993). No obstante, lacantidad de este ácido no va disminuyendo con el tiempo a medida que las moscaslo consumen. Este se produce continuamente en el medio debido a la acción de microorganismos que se desarrollan en él. En primer lugar, la levadura metaboliza lashexosas por fermentación produciendo metabolitos secundarios como alcohol y gascarbónico (Mathews et al., 2002). En segundo lugar, este alcohol es metabolizado porvarios tipos de bacterias, las cuales producen ácido acético, alimento principal de lasmoscas como metabolito secundario (Madigan et al., 2004). De esta manera, unbuen medio debe presentar las características físico-químicas necesarias para unadecuado desarrollo de estos microorganismos. Lo que permitiría que tanto losprocesos de ovogénesis como los de espermatogénesis y oviposición conlleven a unamayor productividad. Las principales características físico-químicas que afectan aldesarrollo de estos microorganismos en el medio de cultivo son: porcentaje de hexo-sas (alimento de la levadura), nutrientes minerales necesarios para su formación, ypH (estos nutrientes son absorbidos del medio por diferencias de concentración ióni-ca entre el medio y el citoplasma; Madigan et al., 2004). De estas tres características,parece ser que tuvo mayor trascendencia en términos de productividad en D.melanogasterla acidez, ya que precisamente el medio preparado con la fruta másácida, la naranja, fue el más productivo. Sin embargo, con el porcentaje de hexosasocurre lo contrario, siendo la fruta de mayor porcentaje, el banano. Posiblemente,estos resultados evidencian que no es suficiente con tener altos niveles de nutrientespara constituir un buen medio de cultivo, parece ser mucho más importante carac-terísticas físico-químicas como pH que permite captar la poca cantidad de nutrientesque hay en el medio. Sin embargo, esta afirmación constituye simplemente una hipó-tesis de trabajo que puede ser puesta a prueba en un trabajo posterior. Por otro lado,el hecho de haber encontrado diferencias significativas entre las cepas evaluadas,evidencia el efecto del tipo de cepa de D. melanogasteren la productividad y tiempo dedesarrollo (Bonnier y Jonson, 1957). Esto se debe a que en muchos mutantes se venafectadas características estructurales y funcionales que influencian la tasa de mor-talidad (Gluecksohn-Waelsch, 1953; Shearn et al., 1971), la viabilidad y/o el tiempode desarrollo de la especie, tales como el comportamiento de cortejo, la capacidadde cópula, la oviposición y la viabilidad tanto del esperma como de los diferentes estadios de la metamorfosis (Mora et al., 2000). De este modo, para los dos medios evaluados, la cepa silvestre fue la más productiva con el menor tiempo de desarrollo independientemente del medio de cultivo, debido a que dicha cepa no presenta en absoluto ninguna de las desventajas descritas anteriormente para la mayoría de losmutantes. Vg,al mostrar una disminución en el tamaño del área del ala y un cambioen la posición de la línea a lo largo de la cual el ala se despliega del disco imagal(Ramos, 1993), tiene dificultades en el cortejo, ya que el éxito de apareamientodepende de las vibraciones de las alas durante el mismo (Geer y Green, 1962; Scout,1994). La cepa CyLvpuede tener disminuida la productividad y/o el tiempo de desa-rrollo principalmente debido a que la mutación Curlyse comporta como un gen letalen homocigosis (Wallace, 1965), y solo en algunas ocasiones pueden emerger comoadultos enanos (Truman, 1947). Además, al igual que vestigialla deficiencia en susalas provocada por la contracción desigual de los epitelios superior e inferior duranteel periodo de secado después que la mosca emerge de la pupa, provoca que el cortejosea deficiente y por tanto, la productividad de adultos sea menor (Ramos, 1993). Porotra parte, wpresenta ausencia de pigmentación en sus ojos, lo que le otorgainsuficiencias visuales que afectan el cortejo, de tal forma que se diferencia de lassilvestres y de las otras cepas investigadas porque mientras éstas realizan cortejoscoordinados en el día y aleatorios en la noche, w siempre realiza cortejos aleatorios.Es decir, no diferencia entre cortejos macho-macho y macho-hembra. Además, laausencia de visión hace que al igual que las mutaciones dependientes de alas, no hayauna vibración alar apropiada para el apareamiento, ya que la última estimulación delcortejo es visual con las vibraciones de las alas (Geer y Green, 1962). Estas defi-ciencias para cada una de las cepas evaluadas explican su baja productividad y largotiempo de desarrollo en comparación con la cepa silvestre. Sin embargo, no explicanel hecho de que el orden en que se encontraron las diferencias entre cepas, no semantuviera para la productividad y el tiempo de desarrollo, ni de un medio a otro.Esto puede ser explicado por el efecto de interacción evidenciado en el análisis devarianza para productividad y tiempo de desarrollo. Esta interacción cepa-mediopuede ocurrir debido a que existen reportes que indican que el efecto deletéreo de unadeterminada mutación puede ser modificado por la influencia del medio ambiente.Así, una cepa puede expresar totalmente su carácter deficiente en un ambiente comoel medio de cultivo, pero puede hacerlo parcialmente en otro tipo de medio. Inclusocuando es posible ajustar el medio ambiente a las necesidades y requerimientos de unorganismo, la mutación puede perder totalmente su carácter deficiente (Gluecksohn-Waelsch, 1953). Esto explica el hecho de que algunas de las cepas hallan resultadosignificativamente iguales a la silvestre en algunas situaciones, lo que ocurrió para laproductividad con wy CyLven el medio de banano, vgen naranja, y para el tiempo de desarrollo con w y vg en el medio de banano. Igualmente, la interacción encontrada explica el hecho de que el medio de naranja presentara mayor productividad para la totalidad de las cepas excepto para w y CyLv. En el análisis genético solo fue posible tener una estimación significativa de la heredabilidad para la productividad. El error estándar de la heredabilidad para el tiempo de desarrollo fue demasiado alto, por lo que se recomienda aumentar el número de cepas, medios o repeticiones para una mejor estimación. Para la productividad, la heredabilidad presentó un valor dentro del rango considerado bajo (h2 < 0,3; Mitchell-Oldd, 1986), lo que evidencia que la variación de éste carácter en la población de laboratorio evaluada es debida en una muy baja proporción a los efectos de las diferencias genotípicas. Esto concuerda con los valores reportados en la literatura en cuanto a la heredabilidad para caracteres asociados con adecuación biológica, fitness, poligénicos y se encuentran bajo la influencia de una alta cantidad de factores ambientales como por ejemplo la viabilidad y altura (Molina, 1992). Así, estos caracteres presentan alto componente ambiental pero bajo componente genético (Mitchell-Oldd, 1986). Sin embargo, en la productividad, al igual que el componente genético, la varianza ambiental fue muy baja, por lo que la mayor parte de la variación fenotípica se debió a las diferencias ocasionadas por la interacción genotipo-ambiente (Gordon et al., 1972). Esto explica nuevamente el hecho de que las cepas no presentarán el mismo patrón de productividad y tiempo de desarrollo en los dos medios de cultivo. El hecho de que la productividad no presentara correlación respecto al tiempo de desarrollo sugiere que la mayor parte de los genes que regulan estos caracteres difieren y/o interaccionan de manera distinta en los ambientes evaluados. Estas evidencias sugieren que cualquier tipo de experimentación en D. melanogaster que necesite un manejo de su productividad deberá tener en cuenta, además, del tipo de mutante a utilizar, también las diferencias de expresión en un determinado medio.
AGRADECIMIENTOS
A los integrantes del Grupo de Estudio y Trabajo en Genética -GETEG- que participaron en la toma de datos del proyecto. Al Departamento de Biología por su aporte económico, a la Sección de Genética por la infraestructura facilitada y colaboración de sus integrantes, y por último, a los empleados del Laboratorio de Biología de la Universidad del Valle, quienes intervinieron en la preparación de los reactivos utilizados durante el proyecto.
BIBLIOGRAFÍA
ALVARADO MC. Estimación de los componentes genéticos y ambientales de los tiempos de desarrollo huevo-pupa y pupa-adulto de algunos mutantes del cromosoma II de Drosophila melanogaster [trabajo de grado]. Santiago de Cali: Departamento de Biología, Facultad de ciencias, Universidad del Valle; 2000. [ Links ]
BALBÍN A, ROJAS Y, CHICA C, CAMPOS A. Efecto de la temperatura y del medio de cultivo de dos generaciones hijas de un cruce dihíbrido en Drosophila melanogaster. Acta biol Colomb. 2000;1(5):47-57. [ Links ]
BONNIER G, JONSSON U. Rate of Development of Viability Mutants in Drosophila melanogaster. Evolution. 1957;3(11):271-279. [ Links ]
DÍAZ F, MAYOR V, GONZÁLEZ F, AGUIRRE D, PIZARRO M, ARIAS F, VILLACÍS M, CÁRDENAS H. Efecto del medio de cultivo en la productividad de una cepa silvestre de Drosophila melanogaster en: XL Congreso de la Asociación Colombiana de Ciencias Biológicas. 11 al 14 de octubre del 2005. Cali, Universidad Autónoma de Occidente; 2005. [ Links ]
GEER B, GREEN M. Genotype, Phenotype and Mating Behaviour of Drosophila melanogaster. Am Nat. 1962;888(96):175-181. [ Links ]
GLUECKSOHN-WAELSCH S. Lethal Factors in Development. Q Rev Biol. 1953;2(28):115-135. [ Links ]
GODOY-HERRERA R. La conducta de larvas de Drosophila (Diptera; Drosophilidae): Su etología, desarrollo, genética y evolución. Rev Chil Hist Nat. 2001; 1(74):55-64. [ Links ]
GORDON LL, BYTH DE, BALAM LN. Variance of Heritability Ratios Estimated From Phenotypic Variance. Biometrics. 1972;2(28):401-415. [ Links ]
MADIGAN M, MARTINKO JM, PARKER J. Brock biología de los microorganismos. 10ª ed. Madrid: Editorial Prentice Hall; 2004. [ Links ]
MATHEWS CK, VAN-HOLDE KE, AHERN KG. Bioquímica. 3ª ed. Madrid: Editorial Pearson Education S.A.; 2002. [ Links ]
MITCHELL-OLDS T. Quantitative Genetics of Survival and Growth in Impatiens Capensis. Evolution. 1986;1(40):107-116. [ Links ]
MOLINA JD. Introducción a la genética de poblaciones y cuantitativa. 1ª ed. Ciudad de México: Editorial AGT Editor S.A.; 1992. [ Links ]
MORA F, SANTOS F, CAMPOS H. Efecto del doble mutante e//e w//w y el medio del cultivo en la productividad de Drosophila melanogaster. Acta biol Colomb. 2000;1(5):39-46. [ Links ]
MORGAN T. Sex-Limited and Sex-Linked Inheritance. Am Nat. 1914;574(48):577-583. [ Links ]
POLACZYK PJ, GASPERINI R, GIBSON G. Naturally Occurring Genetic Variation Affects Drosophila photoreceptor Determination. Dev Genes Evol. 1998;207:462-470. [ Links ]
RAMOS PM. Manual de genética para Drosophila melanogaster. 1ª ed. Ciudad de México: Editorial McGraw Hill; 1993. [ Links ]
SHEARN A, RICE T, GAREN A, GEHRING W. Imaginal Disc Abnormalities in Lethal Mutants of Drosophila. Proc Nat Acad Sci USA. 1971;10(68):2594-2598. [ Links ]
TRUMAN P. Second Chromosome Inversions in Wild Populations of Drosophila melanogaster. Evolution. 1947;172(1):42-47. [ Links ]
WALLACE B. The Viabily Effects of Spontaneous Mutations in Drosophila melanogaster. Am Nat. 1965;908(99):335-348. [ Links ] ]]>