: p < 0,01) y específica (Sjj': p < 0,05) en el dialelo de LS1, explicaron las diferencias para h jj'. Según la metodología de HyM, la acción génica aditiva (HCG: p < 0,01) y no aditiva (HCE: p < 0,05), controlaron la expresión del RFP en el dialelo de LS1, con mayor importancia de la HCG. En el dialelo VAR, sólo la HCG (p < 0,01) explicó tal control. Las variedades Bolo Verde (RFP = 17,8 kg/planta; v j = 3,34**; g i = 1,73*) y Soler (PFP = 17 kg/planta; v j = 2,89*; g i = 175*), resultaron ser los genotipos recomendados para explotar los efectos aditivos y el híbrido LA34 x LSO (RFP = 26 kg/planta; Sjj'= 3,55**) para explotar los efectos aditivos y no aditivos mediante selección recurrente recíproca.]]>
: p < 0,01) and specific heterosis (Sjj': p < 0,05) explained the differences for h jj in the diallelic of LS1. According to the methodology of HyM, the additive (GCA: p < 0,01) and non additive genic action (SCA: p < 0,05) controlled the expression of RFP in the diallele of LS1 with greater importance of the GCA. In the diallele of VAR, only the GCA (p < 0,01) explained such control. The genotypes Bolo Verde (RFP = 17,8 kg/plant; v j = 3,34**; g i = 1,73*) and Soler (PFP = 17 kg/plant; v j = 2,89*; g i = 175*) are the recommended genotypes to exploit the additives effects and the hybrid LA34 x LSO (RFP = 26 kg/plant; Sjj'= 3,55**) to exploit the additive and non additive effects through reciprocal recurrent selection.]]>
EFECTOS HETERÓTICOS Y HABILIDAD COMBINATORIA PARA EL RENDIMIENTO POR PLANTA EN Cucurbita moschata DUCH. Ex Poir
HETEROTIC EFFECTS AND COMBINING ABILITY FOR YIELD PER PLANTIN Cucurbita moschata DUCH. Ex Poir
Miguel Mariano Espitia Camacho1; Franco Alirio Vallejo Cabrera2 y Diosdado Baena García3
1 Profesor Asociado. Universidad de Córdoba. Facultad de Ciencias Agrícolas. Código Postal 354, Montería, Colombia. <mespitia@unicordoba.edu.co > 2 Profesor Titular. Universidad Nacional de Colombia, Sede Palmira. Facultad de Ciencias Agropecuarias. A.A. 0237. Palmira, Colombia. <favallejoc@palmira.unal.edu.co>, 3 Profesor Asociado. Universidad Nacional de Colombia, Sede Palmira. Facultad de Ciencias Agropecuarias. A.A. 0237. Palmira, Colombia. <dbaenag@palmira.unal.edu.co>
Recibido: Septiembre 6 de 2005; aceptado: Enero 27 de 2006.
]]>
RESUMEN
El estudio tuvo como objetivos estimar los efectos heteróticos y de habilidad combinatoria (HCG) y específica (HCE), para el rendimiento/planta (RFP), en dos dialelos [uno entre cinco variedades (VAR) y el otro entre cinco líneas endogámicas S1 (LS1), derivadas de éstas], mediante las metodologías propuestas por Gardner y Eberhart (GyE: 1966: Análisis II) y Hallauer y Miranda (HyM:1981). Los genotipos se evaluaron en Candelaria (Valle del Cauca, Colombia), en un diseño de bloques completos al azar con cinco repeticiones. Las diferencias entre genotipos en el dialelo de LS1 por GyE, fueron explicadas por los efectos de variedades (vj: p <0,01) y heterosis (hjj: p < 0,01) y el dialelo de VAR sólo, por los efectos de (vj: p < 0,01). La heterosis media (: p < 0,01) y específica (Sjj: p < 0,05) en el dialelo de LS1, explicaron las diferencias para hjj. Según la metodología de HyM, la acción génica aditiva (HCG: p < 0,01) y no aditiva (HCE: p < 0,05), controlaron la expresión del RFP en el dialelo de LS1, con mayor importancia de la HCG. En el dialelo VAR, sólo la HCG (p < 0,01) explicó tal control. Las variedades Bolo Verde (RFP = 17,8 kg/planta; vj = 3,34**; gi = 1,73*) y Soler (PFP = 17 kg/planta; vj = 2,89*; gi = 175*), resultaron ser los genotipos recomendados para explotar los efectos aditivos y el híbrido LA34 x LSO (RFP = 26 kg/planta; Sjj= 3,55**) para explotar los efectos aditivos y no aditivos mediante selección recurrente recíproca.
The purpose of this study was to estimate the heterotic effects, general combining ability (GCA), and specific (SCA) for fruit yield per plant (RFP) in two diallelic [one among five cross pollinated varieties (VAR) and the other among five self-pollinated lines S1 (LS1), derived from these], according to the methodologies designed by Gardner y Eberhart (GyE: 1966: Analysis II) and Hallauer and Miranda (HyM: 1981) using a complete randomized design with five replicates in Candelaria (Valle del Cauca, Colombia). The differences between genotypes observed in the diallelic of LS1 by GyE, were explained by the effects of varieties (vj: p <0,01) and heterosis(hjj: p < 0,01), and in the diallelic of VAR only by the effects of varieties (vj : p < 0,01). The mean heterosis (: p < 0,01) and specific heterosis (Sjj: p < 0,05) explained the differences for hjj in the diallelic of LS1. According to the methodology of HyM, the additive (GCA: p < 0,01) and non additive genic action (SCA: p < 0,05) controlled the expression of RFP in the diallele of LS1 with greater importance of the GCA. In the diallele of VAR, only the GCA (p < 0,01) explained such control. The genotypes Bolo Verde (RFP = 17,8 kg/plant; vj = 3,34**; gi = 1,73*) and Soler (PFP = 17 kg/plant; vj = 2,89*; gi = 175*) are the recommended genotypes to exploit the additives effects and the hybrid LA34 x LSO (RFP = 26 kg/plant; Sjj= 3,55**) to exploit the additive and non additive effects through reciprocal recurrent selection.
MATERIALES Y MÉTODOS RESULTADOS Y DISCUSIÓN DISCUSIÓN CONCLUSIONES ]]>
BIBLIOGRAFÍA
Los estudios de efectos heteróticos y habilidad combinatoria de razas o variedades en alógamas con germoplasma mejorado, han sido utilizados desde hace mucho tiempo en el mejoramiento de muchas especies y ha demostrado que constituye una buena alternativa para un mejor aprovechamiento de genes deseables, que puedan ser integrados en poblaciones mejoradas y aumentar su base genética (Pérez et al. 1995 y Machado y Miranda 2003).
El análisis de la habilidad combinatoria general (HCG), permite identificar adecuadamente los progenitores con capacidad para transmitir sus caracteres deseables a la descendencia, y la habilidad combinatoria específica (HCE) posibilita conocer aquellas combinaciones híbridas F1 sobresalientes, originadas de cruzamientos entre variedades, línea o líneas por variedad. Igualmente este tipo de análisis facilita información sobre el tipo de acción génica que condiciona la expresión de un carácter, lo cual es básico en la escogencia del método de mejoramiento a seguir. Según Sprague y Tatum 1942, la HCG designa el comportamiento promedio de una línea en sus combinaciones híbridas y está asociada con genes de efectos principalmente aditivos (acción génica aditiva). La HCE designa aquellos casos en que ciertos cruzamientos tienen un comportamiento mejor o peor del que se puede esperar sobre la base del comportamiento promedio de sus progenitores y está asociada con genes de efectos dominantes, epistáticos y otros tipos de interacciones génicas (acción génica noaditiva).
En un programa de mejoramiento de cualquier especie, debe darse gran importancia a los estudios genéticos de los caracteres relacionados con el rendimiento y la calidad (muchos de los cuales son de tipo poligénico), como una de las mejores formas de evaluar el potencial genético del germoplasma, así como también para aumentar la eficiencia de los métodos de mejoramiento.
Los cruzamientos dialélicos constituyen uno de los sistemas de apareamiento más utilizados en la actualidad para la estimación confiable y segura de HCG y HCE lo cual es de considerable valor para entender la naturaleza de la acción génica involucrada en la determinación de rasgos cuantitativos.
Entre las metodologías de análisis para un dialélico se encuentran las propuestas por: Hayman 1954, 1959, Griffing 1956, Gardner y Eberhart 1966 y Hallauer y Miranda 1981. Vallejo y Estrada 2002, señalan que en plantas autógamas, el análisis de los cruzamientos dialélicos se fundamenta en dos metodologías básicas: a) La metodología propuesta por Hayman 1954, 1958 y b) la propuesta por Griffing 1956, basada en un modelo más amplio. De igual forma en alógamas son usados los métodos propuestos por Gardner y Eberhart 1966 y Hallauer y Miranda 1981.
La metodología de Griffing 1956, fue desarrollada para evaluar la HCG y la HCE de los progenitores que intervienen en el cruzamiento dialélico. El métodos de análisis dialélico propuesto por Hayman 1954, 1958, fue desarrollado para ser utilizado en cruzamientos dialélicos que incluyen líneas homocigotas únicamente como progenitores, siendo por lo tanto, usado ampliamente en plantas autógamas.
Gardner y Eberhart 1966, propusieron varios métodos de análisis, siendo el más utilizado el análisis tipo II. Esta metodología puede aplicarse desde grupos parentales totalmente homocigotos (F = 1) hasta aquellos sin ningún grado de endogamia (F=0). Típicamente requiere de la evaluación de los padres y los cruzamientos F1 directos. Este método considera sólo modelos fijos, y por lo tanto, no tiene sentido aplicarlo para la estimación de componentes de varianza genética (modelos aleatorios).
En su trabajo Gardner y Eberhart 1966, señalan que esta metodología permite un análisis más preciso y detallado de los efectos heteróticos que se manifiestan en los cruzamientos. En general este método se ha usado con más frecuencia en el análisis de cruzamientos dialélicos, en la que los padres son variedades de polinización abierta (Pérez et al. 1995 y Machado y Miranda 2003), igualmente ha sido empleado en algunas autógamas como la soya (Pandini, Vello y Celis 2002).
]]>
El método de análisis propuesto por Hallauer y Miranda 1981, es una combinación de distintas propuestas e involucra los principios de las metodologías de Gardner y Eberhart 1966 (método III) y la de Griffing 1956 (método 4), siendo por ello uno de los métodos más utilizados en maíz (Hallauer y Miranda 1981 y Ceballos 2003).
La literatura reporta que la variación genética del rendimiento/planta en C. moschata, ha estado algunas veces bajo el control de los efectos de acción génica aditiva (Vinasco, Baena y García 1998) no aditiva (Korzeniewska y Niemirowicz 1993, Mohanty, Mohanty y Mishra 1999, Mohanty y Mishra 1998, 2000, Sirohi y Behera 2000, Mohanty 2000b, 2000a, 2001a, 2001b, y 2001c) y otras veces bajo los efectos de acción génica aditiva y no aditiva de manera conjunta (Doijode y Sulladmath 1985, Gwanama, Botha y Labuschagne 2001 y Mohanty y Prusti 2002), predominando dentro de la acción génica no aditiva, los efectos de dominancia y epístasis y en menor proporción la sobredominancia.
La importancia del zapallo (C. moschata) ha sido reconocida en Colombia por su área de siembra, producción, versatilidad en consumo directo, alimento saludable, materia prima para la agroindustria, artesanías, decoración, calidad nutricional, cultivo rústico, potencial de exportación, cultivos propios y ligados a pequeños agricultores (Espitia 2004). Sin embargo, son muy escasos los estudios realizados sobre efectos heteróticos y habilidad combinatoria para el rendimiento. Lo anterior señala un vacío en la investigación en este aspecto, sobre todo, si se tiene en cuenta que la Universidad Nacional de Colombia, Sede Palmira conformó una colección de germoplasma de esta especie, producto de recientes misiones de colectas en gran parte del territorio nacional, lo cual hizo necesario conocer el potencial per se o en cruzamientos de tales accesiones, junto con otras introducciones de diferentes regiones del mundo.
El estudio tuvo como objetivos específicos: 1) cuantificar y analizar los efectos heteróticos de los progenitores y los cruzamientos F1, en dos dialelos de C. moschata de cinco progenitores cada uno (uno entre cinco variedades y otro entre cinco líneas endocridas S1, derivadas de éstas), mediante la metodología propuesta por Gardner y Eberhart 1966 (Análisis II) y 2 ) determinar la importancia de los efectos de habilidad combinatoria general (HCG) y específica (HCE) en cada uno de los dos dialélicos anteriores, mediante la metodología propuesta por Griffing 1956 (Modelo I Método 4), modificada por Hallauer y Miranda 1981, para el rendimiento/planta.
MATERIALES Y MÉTODOS
El trabajo se realizó durante los años 2002 y 2003, en el Centro Experimental de la Universidad Nacional de Colombia, Sede Palmira (CEUNP), ubicado en el municipio de Candelaria (Valle del Cauca, Colombia), según la clasificación de Holdridge, la zona pertenece a la formación Bosque Seco Tropical (BS-T) (Rodríguez 1999).
Los datos utilizados en el presente estudio se obtuvieron de la evaluación agronómica de dos estudios dialélicos de zapallo (C. moschata) de cinco progenitores cada uno. Uno entre cinco variedades de libre polinizacion: Bolo Verde (BV); Soler (SO); Accesión 88 (A88); Accesión 34 (A34) y Accesión 126 (A126) y otro entre cinco líneas endogámicas S1: LBV; LSO; LA88; LA34 y LA126, obtenidas de las anteriores variedades. En cada dialélico se evaluaron 15 genotipos (5 progenitores + 10 cruzamientos directos).
Se evaluaron los siguientes caracteres de interés agronómico: días a floración masculina y femenina, número de frutos por planta, peso promedio de fruto, rendimiento por planta, grosor de la pulpa, porcentaje de pulpa por fruto, diámetro de la cavidad interna del fruto, peso de 100 semillas, peso de semillas por fruto y número de semillas por fruto. Sin embargo, aquí solo se presentan los resultados obtenidos para el rendimiento por planta, por considerar a este carácter como uno de los más importantes para el agricultor.
La evaluación agronómica de los 15 genotipos de cada dialélico, se realizó mediante el uso del diseño de bloques completos al azar con cinco repeticiones. las distancia de siembra utilizada fue de 3 x 3 metros en cuadro. La unidad experimental efectiva consistió de un surco de seis plantas (54 m2). A pesar de que los datos se tomaron sobre plantas individuales, los análisis se realizaron con base en los promedios por unidad experimental.
]]>
Los análisis genético-estadísticos para estimar los efectos heteróticos y la habilidad combinatoria de los diferentes genotipos, se realizaron de manera individual para cada dialélico, empleando las metodologías propuestas por Gardner y Eberhart 1966: análisis II y Griffing 1956: Modelo 1- método 4, modificada por Hallauer y Miranda 1981. Por ser estas metodologías ampliamente conocidas en el fitomejoramiento, sólo se describirán los aspectos más generales.
Método de Gardner y Eberhart 1966. Gardner y Eberhart 1966 desarrollaron varios tipos de análisis, siendo el más comúnmente utilizado el análisis II, el cual se usó en este estudio. En este modelo, la expresión del valor fenotípico promedio de cualquier padre o híbrido F1, está dada por la siguiente ecuación: Donde: Yjj: es el valor promedio esperado para una variedad (j = j) o para un cruzamiento (j ¹ j). Si j = j, entonces q = 0 y sí j ¹ j, entonces q = 1. mv: es la media de las n variedades parentales. Vj: es el efecto varietal, de la j-ésima variedad. hjj: es el efecto de heterosis que resulta cuando la variedad j se cruza con la variedad j. Este efecto ocurre solamente en los cruzamientos F1. El efecto de la heterosis a su vez puede dividirse así: Donde: : es la heterosis promedio de todos los cruzamientos. hj: es la heterosis varietal producida por la j-ésima variedad y Sjj: es la heterosis específica para el cruzamiento entre las variedades j y j; es una desviación de la media esperada basada en los efectos + hj.
De acuerdo con Gardner 1967, el efecto de la variedad (Vj), es la diferencia entre la media de un padre per se y la media de todos los padres. La heterosis promedio (), del grupo de progenitores usados en el estudio es la diferencia entre la media de todos los cruzamientos F1 y la media de todos los progenitores. La heterosis varietal (hj), es la contribución de heterosis por la variedad j en sus cruzamientos F1, medida como una desviación del promedio de heterosis. La heterosis específica (Sjj) entre las variedades j y j, mide la desviación entre el comportamiento observado de un cruzamiento específico y su comportamiento esperado, basado en el efecto varietal (Vj), la heterosis promedio () y la heterosis varietal (hj).
Método 4 modelo 1 de Griffing 1956, modificado por Hallauer y Miranda 1981. Esta metodología propone un análisis que combina las propuestas de Gardner y Eberhart 1966 (análisis III) y Griffing (método 4 modelo 1: 1956). El modelo que explica la variación de las variables, bajo el diseño de bloques completos al azar para cada dialélico, considerando los datos de los progenitores y sus cruzamientos F1, es el siguiente:
Donde Yijk = valor fenotípico del genotipo (ij) dentro del bloque (k), m = efecto de la media general del experimento, gij = efecto del genotipo ij ésimo, bk = efecto del bloque k ésimo y eijk = error experimental. Con el objeto de establecer la significancia para las fuentes de variación propuestas por Gardner y Eberhart 1966 (análisis III): la fuente de variación: genotipos, se descompone en: padres (P), cruzamientos F1(C) y el contraste P versus C.
La metodología de Griffing 1956 (modelo 1 método 4) permite una estimación más precisa de los efectos de HCG y HCE, dado que no considera en los cálculos la habilidad combinatoria de los padres consigo mismo, aún cuando éstos se hallan incluidos conjuntamente con los cruzamientos F1 en las evaluaciones de campo. El análisis descompone la variación de los cruzamientos en términos de efectos de HCG y HCE de acuerdo al siguiente modelo genético y estadístico: Yij = m + gi + gj + Sij + åå eijk. Donde: m = media poblacional de todos los cruzamientos; gi = efecto de habilidad combinatoria general del progenitor i; gj = efecto de habilidad combinatoria general del progenitor j; Sij = efecto de habilidad combinatoria específica del híbrido entre los progenitores i x j, con Sij = Sji, eijk = error ambiental asociado con la observación Yijk; Yij = valor promedio fenotípico observado de la variable en estudio para el cruzamiento (ij). Los supuestos del modelo son: ågi = 0, porque los efectos gi son desviaciones con respecto a la media general del experimento m; åSij = 0, porque los efectos de Sij miden las desviaciones respecto a las expectativas basadas en el promedio de HCG de las variedades o líneas progenitoras de cada híbrido.
Los cálculos para la estimación de los análisis de varianzas y los efectos genéticos para el carácter de interés, en cada una de las metodologías, se realizaron mediante el uso de los programas computacionales SAS (SAS Institute Inc., 2000) y GENES (Cruz, 2004).
RESULTADOS Y DISCUSIÓN
]]>
Análisis de varianza (ANDEVA). Para las dos metodologías de análisis se observaron diferencias altamente significativas para genotipos. Lo anterior permite rechazar la hipótesis nula de igualdad de medias entre los genotipos (progenitores y cruzamientos) y justifica continuar con el análisis e interpretación de los resultados (Tabla 1).
Tabla 1. Cuadrados medios (CM) en el análisis dialélico entre cinco variedades (VAR) y cinco líneas S1 (LS1) de zapallo (C. moschata), para el rendimiento/planta (RFP), según el análisis II de Gardner y Eberhart (GyE: 1966) y el métodod de Hallauer y Miranda (HyM: 1981).
Los cuadrados medios asociados a los efectos de variedades (vj) y heterosis (hjj) resultaron significativos, en el dialelo de líneas S1 (LS1), evidenciando que las líneas no constituyen un grupo genéticamente homogéneo y que hay manifestación de heterosis en sus cruzamientos.
A pesar de lo anterior los efectos de heterosis resultaron ser el componente más importante, ya que explicaron el 76 % de la variación de la suma de cuadrados para genotipos. Situación contraria ocurrió en el dialelo varietal (VAR), ya que sólo las variedades (vj) fueron un componente estadísticamente importante (p < 0,01) en el desglose de la variación de los genotipos, respondiendo por el 71 % de la variación de la suma de cuadrados de éstos. Ello señala una mayor importancia de los efectos de acción génica aditiva en relación con los no aditivos, en el dialelo de variedades.
La descomposición de los efectos de heterosis total (hjj) en el dialelo de LS1, permite verificar la significancia estadística de la heteroris media () y la heterosis específica (Sjj), indicando que cada cruzamiento tuvo un efecto heterótico propio. La heterosis media fue la componente más importante, en razón a que explicó el 81 % de la variación de la suma de cuadrados de la heterosis. Por lo anterior se espera que el uso del modelo 4: de acuerdo a GyE, explique, aún cuando no totalmente, la variación observada en las medias de la tabla dialélica de los genotipos para el rendimiento/planta, ya que la heterosis varietal (hj), no resultó significativa.
En el dialelo VAR, a pesar de que la heterosis total (hjj) no fue significativa, la heterosis media () si presentó diferencias estadísticas (p< 0,01), explicando el 67,5 % de la variación de la suma de cuadrados de la heterosis. Por lo tanto constituye una evidencia para señalar que las medias de los progenitores y las de los cruzamientos varietales son diferentes estadísticamente; por ello se admite la existencia de heterosis, más sin embargo, ella es la misma para todos los cruzamientos varietales. De acuerdo con ello se supone el modelo 2: Yij = m + (vi + vj)/2 + + eij de GyE, como la opción más probable para explicar la variación existente en las medias de los genotipos (variedades e híbridos varietales) en la tabla dialélica varietal.
Los resultados obtenidos en la metodología propuesta por HyM (Tabla 1), permitieron detectar diferencias estadísticas significativas (p < 0,05 o p < 0,01) para todas las fuentes de variación, excepto para padres (P) en el dialelo de LS1 y habilidad combinatoria específica (HCE) en el dialelo VAR. Estos resultados confirman las diferencias estadísticas obtenidas a nivel de los cuadrados medios de genotipos, heterosis media () y heterosis específica (Sjj) en el análisis de GyE en los dos dialelos, los cuales son idénticos con genotipos, padres versus cruzamientos (P vs C) y HCE, respectivamente.
Los niveles de significancia estadística obtenidos en los dos dialelos, en el cuadrado medio de Padres (P), sugiere que existen diferencias de origen genético en al menos una de las variedades, mientras que en las líneas S1 no existe suficiente evidencia estadística para afirmar que alguna de ellas difiere genéticamente de las demás. La existencia de diferencias estadísticas altamente significativas para el contraste P vs C, indican que el comportamiento promedio de todos los cruzamientos F1 (varietales o de líneas S1) fue superior al comportamiento promedio de los progenitores en conjunto, señalando efecto heteróticos significativos en los dos dialelos para el rendimiento/planta. De manera similar, la significancia estadística para cruzamientos (C), demuestran que al menos uno de los cruzamientos intervarietales o entre líneas S1, presentó comportamiento promedio diferente del resto.
No obstante que las tres componentes de genotipos resultaron significativos estadísticamente, la descomposición de su suma de cuadrados, permite detectar que en el dialelo VAR el orden de mayor a menor explicación de dicha variación, fue: cruzamientos (C) con el 51 %, padres (P) con el 30 % y P vs C con el 19 %; mientras que en el dialelo de LS1, fue P vs C con el 61 %, cruzamientos (C) con el 32 % y padres (P) con el 7 %.
]]>
La significancia estadística de los efectos de habilidad combinatoria general (HCG: p < 0,01) y específica (HCE: p < 0,05) en el dialélico de LS1, indica la importancia de la acción génica aditiva y no aditiva en la expresión y control genético del rendimiento/planta en los híbridos entre líneas S1. Los efectos genéticos aditivos (HCG) fueron la componente de variación más importante de los cruzamientos, ya que representaron el 65 % de su suma de cuadrados. En el dialelo de VAR igualmente los efectos aditivos fueron los más importantes, en razón a que sólo la HCG, resultó estadísticamente significativa al nivel de probabilidad del 1 %, explicando el 95 % de la variación de la suma de cuadrados de los cruzamientos. Lo anterior permite deducir que en el dialelo de variedades sólo los efectos genéticos aditivos son los importantes, mientras que en el dialelo de líneas S1, ambos efectos resultaron estadísticamente importantes, pero con diferente contribución.
Los resultados obtenidos en el dialelo de LS1, están de acuerdo con los encontrados por Doijode y Sulladmath 1985, Chattopadhyay, Dana y Mukhopadhyay 1986, Mohanty y Mishra 1998, Mohanty, Mhanty y Mishra 1999, Mohanty y Mishra 2000, Mohanty 2000a, 2000b, 2001b, Gwanama, Botha y Labuschagne 2001, Mohanty 2001c y Mohanty y Prusti 2002; quienes encontraron que los efectos aditivos y no aditivos de los genes, explicaron la variación del rendimiento/planta en un dialélico de líneas endogámicas.
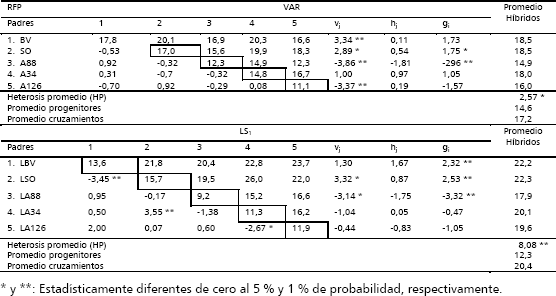
Efectos heteróticos y de habilidad combinatoria general individual. La Tabla 2 permite deducir que para explotar los efectos de acción génica aditiva los genotipos más recomendables son las variedades Bolo Verde (BV) y Soler (SO), y las líneas S1 LBV y LSO, por haber presentado los mayores promedios/planta, altos efectos de variedades (vj), y /o habilidad combinatoria general (gi). Ello indica que estos genotipos presentaron la más alta concentración de genes favorables para incrementar el rendimiento/planta. Por lo tanto, son los más indicados para formar nuevas poblaciones mejoradas, debido a que tienen mayor capacidad de transmitir genes favorables a sus progenies.
Tabla 2. Rendimiento/planta (RFP) para padres (diagonal), híbridos (arriba la diagonal), heterosis específica (Sjj’) (debajo la diagonal), efectos de variedades (vj), heterosis varietal (hj), habilidad combinatoria general (gi) y promedio de los híbridos de cinco progenitores de zapallo (C. moschata), en los dialelos varietal (VAR) y entre líneas S1 (LS1).
Esto favorece la selección de nuevas líneas homocigotas o progenitores superiores, dado que son los genes con efectos aditivos los que responden a la selección. La superioridad del valor como progenitor de los anteriores progenitores se puede corroborar al observar que fueron los que presentaron los mayores promedios en sus híbridos, con valores de 18,5 y 22,3 kg/planta. El comportamiento diferencial en los progenitores en sus valores como progenitor, confirman las diferencias altamente significativas detectadas para HCG en el ANDEVA en ambos dialelos (Tabla 1).
Los resultados permiten deducir alta asociación de los valores como progenitor (vj y gi) entre las variedades y sus respectivas líneas S1, en razón a que las variedades que resultaron deseables o indeseables como progenitor en el dialelo VAR, sus líneas S1 respectivas, también mostraron tendencia en su comportamiento como progenitor muy similar.
Para los efectos de Sjj en los híbridos varietales no se presentaron valores significativos lo que corrobora los niveles de significancia estadística detectados en las dos metodologías de análisis, en el sentido que ninguno de los híbridos varietales presentó valores de heterosis específica significativos. En el dialelo de LS1 tres cruzamientos presentaron efectos de Sjj significativos, son ellos LSO x LBV (-3,45 **), LA126 x LA34(-2,67*) y LA34 x LSO (3,55**). Los valores de heterosis específica negativa y significativas, para los dos primeros sugieren que su promedio fue inferior, al promedio esperado con base en la gi de sus progenitores y la media general. Ello igualmente permite deducir que no existe divergencia y complementación genética entre sus progenitores para el carácter rendimiento/planta, a pesar de que las variedades de donde se originaron tienen origen geográfico diferentes (SO: Puerto Rico; BV: Valle del Cauca y A126: Barranquilla).
El valor de Sjj (3,55**) alto, positivo y significativos (p < 0,01), para el híbrido entre LA34 x LSO, indica que su rendimiento, fue superior al promedio esperado con base en la gi de sus progenitores y la media general. Considerándose por ello como una alternativa genética viable para incrementar la competitividad del zapallo en Colombia, ya que además presentó el mayor rendimiento/planta (26 kg), superando al promedio de todas las líneas progenitoras (12,3 kg), al promedio de todos los cruzamientos F1 entre líneas (20,4 kg) y a la variedad testigo comercial Bolo Verde (BV = 17,8 kg), en 111 %; 27 % y 46 %, respectivamente. De la misma manera sugiere además la presencia de un patrón heterótico muy importante entre estas dos líneas S1.
El híbrido LA34 x LSO se originó del cruzamiento entre un progenitor con altos efectos varietales y habilidad combinatoria general como LSO (vj = 3,32 y gi = 2,53) y otro de bajos efectos para esos estimadores, como: LA34 (vj = -1,04 y gi = -0,47). Las líneas S1 progenitoras de este híbrido: LA34 y LSO, provienen en su orden de la accesión 34 (A34) colectada en la ciudad de Popayán y la variedad mejorada Soler (SO) de Puerto Rico, lo cual permite deducir que ese origen geográfico diferente y una generación de autofecundación en las poblaciones originales, conllevó a diferenciar genéticamente las líneas S1 de las variedades de donde se derivaron, permitiendo la expresión del alto nivel de heterosis específica.
]]>
En la Tabla 2, se puede percibir la significativa expresión de la heterosis promedio, la cual resultó mayor en el dialelo de LS1 (8,08**), en comparación con el dialelo de variedades (2,57*), lo cual respalda lo observado en el análisis de varianza.
DISCUSIÓN
La mayor expresión de la heterosis promedio detectada en este estudio, en el dialelo de LS1 en comparación con el dialelo VAR, está respaldada por lo reportado en la literatura (Hallauer y Miranda 1981, Falconer y Makay 1996, Vasal 2000 y Vallejo y Estrada 2002), en el sentido que en las progenies de líneas endocriadas, se expresa una mayor heterosis en relación con las progenies de poblaciones de base genética amplia (variedades de polinización abierta), siempre y cuando se presenten niveles importantes de divergencia genética entre los progenitores y haya niveles de dominancia unidireccional en la mayor cantidad de loci que controlan el carácter de interés en los progenitores.
Los efectos no aditivos (Sjj y HCE) de los genes fueron más importantes estadísticamente en el dialelo de LS1 en comparación con el dialelo de VAR, situación similar ha sido detectada y reportada en maíz por varios autores (Sprague y Tatum 1942, Miranda y Vencovsky 1984 citado por Machado y Miranda 2003, Beck, Vasal y Crossa 1990, Crossa, Vasal y Beck 1990, Vasal et al. 1992, Miranda 1999, Rezende y Souza Junior 2000). La explicación que dan a tal situación es la mayor divergencia genética (diferencia en frecuencias alélicas) que puede presentarse en los cruzamientos de progenitores de base genética estrecha (líneas) en comparación con los cruzamientos entre genotipos de base genética amplia (variedades), ello debido a que muchos de los loci que controlan el carácter en las poblaciones de base genética amplia, tienen frecuencias alélicas intermedia con baja proporción de alelos fijados en frecuencias 0 ó 1, en comparación con las líneas. Como la heterosis es una función de la diferencia entre frecuencias alélicas entre los progenitores, muchos de los loci en las variedades contribuyen poco a la expresión de la heterosis, aún bajo alta expresión de acción de genes dominantes.
La importancia estadística del tipo de acción génica aditiva (HCG) en los dos dialelos y genes con efectos no aditivos en el dialelo de LS1, controlando la expresión del rendimiento, permite esperar que la explotación de los efectos genéticos aditivos y no aditivos, pueden facilitar un progreso significativo en el mejoramiento genético para incrementar el carácter en estudio a través de las siguientes estrategias: 1) selección recurrente intrapoblacional en los progenitores (variedades y/o líneas S1) con altos efectos varietales y de habilidad combinatoria general (aprovechamiento de la acción génica aditiva); 2) identificación de variedades y/o obtención de líneas superiores, aprovechando la aparición de individuos transgresivos, en las diferentes generaciones segregantes originadas a partir de los híbridos F1 (aprovechamiento de la acción génica aditiva) y 3) aprovechar la acción génica no aditiva, mediante la identificación y evaluación de aquellos híbridos entre líneas con mayores niveles de heterosis útil, heterosis específica, habilidad combinatoria específica y promedios en los caracteres antes mencionados.
La tendencia de alta asociación entre los valores como progenitor (efecto varietal: vj y habilidad combinatoria general: gi) de las variedades y sus líneas endogámicas S1, está de acuerdo con lo reportado en maíz por Hallauer y Miranda 1981, Márquez 1988 y Bauman 1981 citado por Vasal 2000, quienes encontraron alta correlación para HCG, entre las plantas S0 o líneas S1, con sus correspondientes líneas puras en generaciones S5, S6, S7 ó S8, concluyendo que las líneas puras adquieren su individualidad como progenitoras al principio del proceso de autofecundación y tienden a que su HCG permanezca bastante estable en lo sucesivo.
La superioridad de los cultivares Bolo Verde (BV) y Soler (SO) en los valores deseables en sus efectos varietales (vj), habilidad combinatoria general (gi) y en sus rendimientos promedios está explicado por el aporte del mejoramiento genetico, en razón a que estos dos genotipos fueron liberados comercialmente para su siembra en Colombia y Puerto Rico, respectivamente. Por ello constituyen los genotipos más recomendados para explotar los efectos aditivos por selección recurrente con el objeto de mejorar genéticamente tal caracter. No obstante, aunque las líneas S1 de los anteriores genotipos LBV y LSO exhibieron altos efectos de vj y gi, su depresión por endogamia, detectada en sus bajos rendimientos promedios, los torna indeseables, para el presente propósito.
Es de resaltar el cruzamiento formado por LA34 x LSO, el cual constituyó la mejor porcion por sus efectos no aditivos (Sjj = 3,55**) y mayor rendimiento/planta (RFP = 26 kg). Adicionalmente, aún cuando no se reporta en este artículo, este híbrido presentó el mayor número de frutos/planta (5,9 frutos/planta), el menor peso/100 semillas (PCS = 11 g), peso/fruto de 4,9 kg, grosos de pulpa/fruto de 4,9 cm y 515 semillas/fruto. Lo anterior junto con su forma esférica, superficie lisa y color verde de fruto, lo convierten en una excelente alternativa en el corto plazo, para mejorar genéticamente la productividad de zapallo en el país, mediante la explotación de heterosis. Al igual que señala la existencia de un potencial patrón heterótico entre estas dos líneas S1, para explotar en el largo plazo los dos tipos de acción génica, mediante selección recurrente recíproca.
]]>
CONCLUSIONES
En las dos metodologías de estudio (EyG y HyM), la acción génica aditiva y no aditiva, controlaron la expresión de la RFP en el dialelo de LS1, con mayor importancia de los efectos genéticos aditivos.
En el dialelo de VAR, sólo los efectos de acción génica aditiva explicaron el control de la variación de RFP, en las dos metodologías (EyG y HyM).
Los efectos heteróticos fueron importantes solamente en el dialelo de LS1.
Las variedades Bolo Verde y Soler, resultaron ser los genotipos recomendados para mejorar genéticamente el RFP, mediante la explotación de los efectos aditivos por selección recurrente, mientras que el híbrido LA34 x LSO, resultó ideal para explotar ambos efectos genéticos por selección recurrente recíproca.
BIBLIOGRAFÍA
Beck, D. ., Vasal, S. K. and Crossa, J. L. 1990. Heterosis and combining ability of CIMMYTs tropical early and intermediate mature maize germoplasm. En: Maydica. Vol. 35; p.279-285. [ Links ]
Ceballos, H. 2003. Genética cuantitativa y fitomejoramiento. Palmira: Diseños Genéticos, Impreso Universitario. Universidad Nacional de Colombia. 524 p. [ Links ]
Chattopadhyay, P., Dana, S. and Mukhopadhyay, S. 1986. Diallel analysis of five fruit characters in pumpkin (Cucurbita moschata Poir). En: Experimental Genetics. Vol. 2, no. 1/2; p. 33-40. [ Links ]
Crossa, J., Vasal, S. K. and Beck, D. L. 1990. Combining ability in diallel crosses of CIMMYTs tropical late yellow maize germoplasm. En: Maydica. Vol. 35; p. 273-278. [ Links ]
Cruz, C. D. 2004. Programa Genes. Versao Windows. Aplicativo Computacional em Genética e Estatística. Universidade Federal de Vicosa. Disponible en Internet: www.ufv.br/dbg/genes/genes.htm Consultada: 9 Jul. 2005]. [ Links ]
Doijode, S. D. and Sulladmath, U. V. 1985. Genetics of certain fruit characters in pumpkin (Cucurbita moschata Poir.). En: Egyptian Journal of Genetics and Cytology. Vol. 4, no.1; p. 35-40. [ Links ]
Espitia, M. 2004. Estimación y análisis de parámetros genéticos en cruzamientos dialélicos de zapallo (Cucurbita moschata Duch. Exp Poir). Palmira. 206 h. Tesis Doctorado en Ciencias Agropecuarias. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias [ Links ]
Falconer, D. S. and Mackay, T. 1996. Introduction to quantitative genetics. 4 ed. New Jersey: Prentice Hall. 464 p. [ Links ]
Gardner, C. O. 1967. Simplified methods por estimating constants and computing sums of squares for a diallel cross analysis. En: Fitotecnia Latinoamericana. Vol. 4; p. 1-12. [ Links ]
_________ and Eberhart, S. A. 1966. Analysis and interpretation of the variety cross diallel and related populations. En: Biometrics. Vol. 22, no. 3; p. 439-452. [ Links ]
Griffing, B. 1956. Concept of general and specific combining ability in relation to diallelic crossing systems. En: Australian Journal of Biological Science. Vol. 9, no. 4; p. 463-493. [ Links ]
Gwanama, C., Botha, A. Am. and Labuschagne, M.T. 2001. Genetic effects and heterosis of flowering and fruit characteristics of tropical pumpkin. En: Plant Breeding. Vol. 120, no. 3; p. 271-272. [ Links ]
Hallauer, A. R. and Miranda, J. B. 1981. Quantitative genetics in maize breeding. Iowa: The Iowa State University. 468 p. [ Links ]
Hayman, B.I. 1958. The theory and analysis of variance of diallel crosses. Part 2. En: Genetics. Vol. 43, no. 1; p.63-85. [ Links ]
Hayman, B.I. 1954. The theory and analysis of variance of diallel crosses. En: Genetics. Vol. 39, no. 5; p. 789-809. [ Links ]
Korzeniewska, A. and Niemirowicz-szczytt, K. 1993. Combining ability and heterosis effect in winter squash (Cucurbita maxima Duch.). En: Genetica Polonica. Vol. 34, no. 3; p. 259-272. [ Links ]
Machado, R. and Miranda, J. B. 2003. Heterosis expression in crosses between maize populations. year yield. En: Scientia Agricola. Vol. 60, no. 3; p. 519-524. [ Links ]
Márquez, S. F. 1988. Genotecnia vegetal: métodos teoría resultados. México: AGT Editor. v. 2, 665 p. Citado por: Vasal, S. K. 2000. The quality protein maize story. En: Food and Nutritional Bulletin. Vol. 21, no.4; p. 445-450. [ Links ]
Miranda Filho, J. B. 1999. Inbreeding and heterosis. p. 69-80. En: International Symposium, CIMMYT (1997: México) The genetics and exploitation of heterosis in crops. Proceedings.. Madison: Crop Science Society of America . [ Links ]
Mohanty, B. K. 2001a. Genetic analysis of yield and its components in pumpkin. En: Madras Agricultural Journal. Vol. 88, no. 1/3; p. 53-56. [ Links ]
Mohanty, B. K. 2001 b. Genetics of economic traits in pumpkin. En: Indian Agriculturist. Vol. 45, no. 3/4; p. 135-140. [ Links ]
________. 2001 c. Polygenic in-heritance in pumpkin a diallel analysis. En: Horticultural Journal. Vol. 14, no. 1; p. 55-59. [ Links ]
________. 2000 a. Combining ability for yield and its components in pumpkin. En: Indian Journal of Genetics and Plant Breeding. Vol. 60, no. 3; p. 373-379. [ Links ]
________. 2000 b. Gene action for quantitative characters in pumpkin. En: Indian Agriculturist. Vol. 44, no. 3/4; p. 157-163. [ Links ]
________ and Mishra, R. S. 2000. Studies on combining ability for flowering traits in pumpkin. En: Haryana Journal of Horticultural Sciences. Vol. 29, no. 3/4; p. 220-222. [ Links ]
________ and ________. 1998. Genetic studies in pumpkin (Cucurbita moschata Duch. Ex. Poir.). En: Orissa Journal of Horticulture. Vol. 26, no.1; p. 40-43. [ Links ]
________, Mohanty, S. K. and Mishra, R. S. 1999. Genetics of yield and yield components in pumpkin (Cucurbita moschata Duch. Ex Poir) En: Indian Journal of Agriculture Sciences. Vol. 69, no. 11 ; p. 781-783. [ Links ]
________ and Prusti, A. M. 2002. Heterosis and combining ability for polygenic characters in pumpkin. En: Indian Agriculturist. Vol. 46, no.1/2; p. 27-36. [ Links ]
Pandini, F., Vello, A. N. and Celis, D. A. 2002. Heterosis in soybeans for seed yield components and associated traits. En: Brazilian Archives of Biology an Technology. Vol. 45, no. 4; p. 401-412. [ Links ]
Pérez, J. C., Ceballos, H., Pandey, S. and Días, C. 1995. Analysis of diallel crosses among Colombian landraces and improved populations of maize. En: Crop Science. Vol. 35; p. 572-578. [ Links ]
Rezende, G. S. P. and Souza Junior, C. L. 2000. A reciprocal recurrent selection procedure outlined to integrate hybrid breeding program in maize. En: Journal of Genetics and Breeding. Vol. 54; p. 57-66. [ Links ]
Rodríguez, C.A. 1999. Efecto de cuatro métodos de labranza sobre las propiedades físicas de un suelo vertisol ústico y sobre la producción de Sorghum bicolor en el Valle del Cauca. Palmira. 147 h. Trabajo de grado Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
SAS Institute. 2000. SAS/STAT Guide for personal computers. Version 8. Cary, NC: SAS. 378 p. [ Links ]
Sirohi, P. S. and Behera, T. K. 2000. Inheritance of yield and its attributing characters in pumpkin (Cucurbita moschata Duch. ex. Poir). En: Journal of Applied Horticulture ( Lucknow). Vol. 2, no. 2; p. 117-118. [ Links ]
Sprague, G. F. and Tatum, L. A. 1942. General vs specific combining ability in single crosses of corn. En: Journal of the American Society of Agronomy. Vol. 34; p. 923-932. [ Links ]
Vasal, S.K. 2000. The quality protein maize story. En: Food and Nutritional Bulletin. Vol. 21, no. 4; p. 445-450. [ Links ]
Vasal, S. K., Srinivasan, G., Beck, D. L., Crossa, J., Pandey, S. and Leon D. E., S. 1992. Heterosis and combining ability of CIMMYTs tropical late white maize germplasm. En: Maydica, Vol. 37; p. 217-223. [ Links ]
Vallejo, F. A. y Estrada, S. E. 2002. Mejoramiento genético de plantas. Palmira: Feriva. 402 p. [ Links ]
Vinasco, L. E., Baena, G. D. y García, M. 1998. Análisis genético de caracteres que afectan el hábito de crecimiento de zapallo Cucurbita maxima (Duch. Ex Lam). En: Acta Agronómica. Vol. 48, no. 3/4; p. 12-18. [ Links ] ]]>199035279-285Universidad Nacional de Colombia2003198621/21/233-40199035273-2789 Jul. 2198541135-40200419964 ed196741-1219662233439-4521956944463-493200112033271-272The Iowa State University19811958431163-8519543955789-80919933433259-27220036033519-52419882144445-450199969-802001a881/353-562001453/43/4135-1402001141155-5920006033373-3792000 b443/43/4157-1632000293/43/4220-2221998261140-431999691111781-7832002461/21/227-3620024544401-412199535572-57820005457-661999SAS Institute20002000222117-118194234923-93220002144445-450199237217-22320021998483/43/412-18
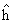 : p < 0,01) y específica (Sjj: p < 0,05) en el dialelo de LS1, explicaron las diferencias para hjj. Según la metodología de HyM, la acción génica aditiva (HCG: p < 0,01) y no aditiva (HCE: p < 0,05), controlaron la expresión del RFP en el dialelo de LS1, con mayor importancia de la HCG. En el dialelo VAR, sólo la HCG (p < 0,01) explicó tal control. Las variedades Bolo Verde (RFP = 17,8 kg/planta; vj = 3,34**; gi = 1,73*) y Soler (PFP = 17 kg/planta; vj = 2,89*; gi = 175*), resultaron ser los genotipos recomendados para explotar los efectos aditivos y el híbrido LA34 x LSO (RFP = 26 kg/planta; Sjj= 3,55**) para explotar los efectos aditivos y no aditivos mediante selección recurrente recíproca.
: p < 0,01) y específica (Sjj: p < 0,05) en el dialelo de LS1, explicaron las diferencias para hjj. Según la metodología de HyM, la acción génica aditiva (HCG: p < 0,01) y no aditiva (HCE: p < 0,05), controlaron la expresión del RFP en el dialelo de LS1, con mayor importancia de la HCG. En el dialelo VAR, sólo la HCG (p < 0,01) explicó tal control. Las variedades Bolo Verde (RFP = 17,8 kg/planta; vj = 3,34**; gi = 1,73*) y Soler (PFP = 17 kg/planta; vj = 2,89*; gi = 175*), resultaron ser los genotipos recomendados para explotar los efectos aditivos y el híbrido LA34 x LSO (RFP = 26 kg/planta; Sjj= 3,55**) para explotar los efectos aditivos y no aditivos mediante selección recurrente recíproca.
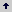
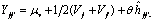 Donde: Yjj: es el valor promedio esperado para una variedad (j = j) o para un cruzamiento (j ¹ j). Si j = j, entonces q = 0 y sí j ¹ j, entonces q = 1. mv: es la media de las n variedades parentales. Vj: es el efecto varietal, de la j-ésima variedad. hjj: es el efecto de heterosis que resulta cuando la variedad j se cruza con la variedad j. Este efecto ocurre solamente en los cruzamientos F1. El efecto de la heterosis a su vez puede dividirse así:
Donde: Yjj: es el valor promedio esperado para una variedad (j = j) o para un cruzamiento (j ¹ j). Si j = j, entonces q = 0 y sí j ¹ j, entonces q = 1. mv: es la media de las n variedades parentales. Vj: es el efecto varietal, de la j-ésima variedad. hjj: es el efecto de heterosis que resulta cuando la variedad j se cruza con la variedad j. Este efecto ocurre solamente en los cruzamientos F1. El efecto de la heterosis a su vez puede dividirse así: 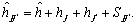 Donde:
Donde: 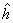 : es la heterosis promedio de todos los cruzamientos. hj: es la heterosis varietal producida por la j-ésima variedad y Sjj: es la heterosis específica para el cruzamiento entre las variedades j y j; es una desviación de la media esperada basada en los efectos
: es la heterosis promedio de todos los cruzamientos. hj: es la heterosis varietal producida por la j-ésima variedad y Sjj: es la heterosis específica para el cruzamiento entre las variedades j y j; es una desviación de la media esperada basada en los efectos 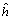 + hj.
+ hj.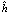 ), del grupo de progenitores usados en el estudio es la diferencia entre la media de todos los cruzamientos F1 y la media de todos los progenitores. La heterosis varietal (hj), es la contribución de heterosis por la variedad j en sus cruzamientos F1, medida como una desviación del promedio de heterosis. La heterosis específica (Sjj) entre las variedades j y j, mide la desviación entre el comportamiento observado de un cruzamiento específico y su comportamiento esperado, basado en el efecto varietal (Vj), la heterosis promedio (
), del grupo de progenitores usados en el estudio es la diferencia entre la media de todos los cruzamientos F1 y la media de todos los progenitores. La heterosis varietal (hj), es la contribución de heterosis por la variedad j en sus cruzamientos F1, medida como una desviación del promedio de heterosis. La heterosis específica (Sjj) entre las variedades j y j, mide la desviación entre el comportamiento observado de un cruzamiento específico y su comportamiento esperado, basado en el efecto varietal (Vj), la heterosis promedio (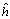 ) y la heterosis varietal (hj).
) y la heterosis varietal (hj).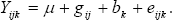

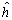 ) y la heterosis específica (Sjj), indicando que cada cruzamiento tuvo un efecto heterótico propio. La heterosis media fue la componente más importante, en razón a que explicó el 81 % de la variación de la suma de cuadrados de la heterosis. Por lo anterior se espera que el uso del modelo 4:
) y la heterosis específica (Sjj), indicando que cada cruzamiento tuvo un efecto heterótico propio. La heterosis media fue la componente más importante, en razón a que explicó el 81 % de la variación de la suma de cuadrados de la heterosis. Por lo anterior se espera que el uso del modelo 4: 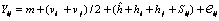 de acuerdo a GyE, explique, aún cuando no totalmente, la variación observada en las medias de la tabla dialélica de los genotipos para el rendimiento/planta, ya que la heterosis varietal (hj), no resultó significativa.
de acuerdo a GyE, explique, aún cuando no totalmente, la variación observada en las medias de la tabla dialélica de los genotipos para el rendimiento/planta, ya que la heterosis varietal (hj), no resultó significativa.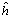 ) si presentó diferencias estadísticas (p< 0,01), explicando el 67,5 % de la variación de la suma de cuadrados de la heterosis. Por lo tanto constituye una evidencia para señalar que las medias de los progenitores y las de los cruzamientos varietales son diferentes estadísticamente; por ello se admite la existencia de heterosis, más sin embargo, ella es la misma para todos los cruzamientos varietales. De acuerdo con ello se supone el modelo 2: Yij = m + (vi + vj)/2 +
) si presentó diferencias estadísticas (p< 0,01), explicando el 67,5 % de la variación de la suma de cuadrados de la heterosis. Por lo tanto constituye una evidencia para señalar que las medias de los progenitores y las de los cruzamientos varietales son diferentes estadísticamente; por ello se admite la existencia de heterosis, más sin embargo, ella es la misma para todos los cruzamientos varietales. De acuerdo con ello se supone el modelo 2: Yij = m + (vi + vj)/2 + 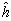 + eij de GyE, como la opción más probable para explicar la variación existente en las medias de los genotipos (variedades e híbridos varietales) en la tabla dialélica varietal.
+ eij de GyE, como la opción más probable para explicar la variación existente en las medias de los genotipos (variedades e híbridos varietales) en la tabla dialélica varietal.