BEES VISITING Aspilia tenella (KUNTH) S .F. BLAKE (ASTERACEAE): FORAGING BEHAVIOR AND POLLEN LOADS
Clara Isabel Aguilar Sierra1 y Allan Henry Smith Pardo2
1 Profesora Asociada. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias. Escuela de Biociencias. A.A. 3840, Medellín, Colombia. <ciaguila@unalmed.edu.co>
2 Profesor Asistente. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. Departamento de Ciencias Agronómicas. A.A. 1779, Medellín, Colombia. <ahsmithp@unal.edu.co>
Recibido: Marzo 27 de 2008; Aceptado: Septiembre 26 de 2008.
Resumen. En cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), se observó el comportamiento de 23 taxones de abejas durante sus visitas a Aspilia tenella (Kunth) S. F. Blake; se registró el número de individuos y el tiempo total de visita y se midieron la temperatura y la humedad relativa. Los análisis de varianza indican que hubo una relación estadísticamente significativa en cuanto a los atributos considerados para las especies de abejas y una covariación significativa con las variables ambientales. Las pruebas de rangos múltiples muestran que Trigona nigerrima es la especie con los mayores valores promedio para el número de individuos y el tiempo de visita. En el estudio de las cargas polínicas se encontraron 30 tipos polínicos, dentro de los cuales A. tenella presentó el mayor porcentaje; Piper aduncum, Miconia minuti flora, Psidium guajava, Rapanea sp., Mimosa pudica y Psychotria sp., en su orden, son otras fuentes de polen importantes para varias de las especies de abejas. El análisis de agrupamiento para las abejas capturadas permitió diferenciar dos grupos: el primero incluye a 18 especies cuyas cargas polínicas que poseían una abundancia relativa de granos de polen de A. tenella por encima de 77%; el segundo grupo estaba conformado por cinco especies de abejas que recolectaron porcentajes mayores de otros tipos polínicos: Exomalopsis sp. 1 (90,4% de M. minuti flora); Lasioglossum sp. 1 y Coelioxys aff. mexicana (77,4% y 64,1% de P. aduncum, respectivamente); Exomalopsis sp. 2 (67,1% de P. guajava) y Auglochloropsis vesta (55,5% de Rapanea sp.). Trigona fulviventris y Apis mellifera presentaron los valores más altos de riqueza de tipos polínicos en las cargas de polen; ello evidencia sus nichos tróficos amplios y su comportamiento generalista de visita.
]]> Palabras claves: Análisis palinológico, tipos polínicos, abejas, cargas polínicas, polinización.Abstract. We observed the behavior of 23 species of wild bees visiting Aspilia tenella (Kunth) S .F. Blake in four different localities of the Porce II dam influence zone ( Antioquia, Colombia ). We recorded number of individuals and total time spent per visit and we measured the temperature and relative humidity. The analyses of the pollen loads showed that out of the thirty pollen types, A. tenella represented the most common of all pollen types, followed by pollen types of Piper aduncum, Miconia minuti flora, Psidium guajava, Rapanea sp., Mimosa pudica y Psychotria sp., in that order. Grouping analyses for the collected bees produced two groups: first, a group of 18 species of bees whose pollen loads had more than 77% of A. tenella pollen grains; and the second group with five species of wild bees whose pollen loads were more abundant in other types of pollen: Exomalopsis sp. 2 (90,4% of M. minuti flora); Lasioglossum sp. 1 and Coelioxys aff. mexicana (77,4% and 64,1% of P. aduncum, respectively); Exomalopsis sp. 2 (67,1% of P. guajava) and Auglochloropsis vesta (55,5% of Rapanea sp.). Trigona fulviventris and Apis mellifera showed the highest richness of pollen types in their pollen loads; this is an evidence of their broad trophic niches and overall generalist feeding habits.
Key words: Palynological analysis, pollen types, bees, pollen loads, pollination.
Son numerosos los insectos que se pueden encontrar en las flores de las angiospermas; el orden Hymenoptera es el más importante, principalmente el suborden Apocrita, el cual incluye visitantes florales frecuentes y algunas relaciones especializadas como el parasitismo, la pseudocópula y la ovoposición (Smith, 1999).
Dentro del suborden Apocrita, muchas abejas (Superfamilia Apoidea) adquieren pasivamente el polen cuando visitan a las flores; pero también pueden colectarlo activamente gracias a varias adaptaciones estructurales y de comportamiento. Aunque algunas llevan el polen internamente en sus canales alimenticios, la mayoría posee estructuras externas especiales para reunir los granos de polen en cargas y transportarlas hasta el nido: pelos en forma de gancho sobre las partes bucales o patas delanteras para facilitar la recolección en flores cuyas anteras están ocultas; grupos de pelos, generalmente en la cabeza, que ayudan a acumular los granos de polen; tibia posterior modificada en corbícula. Otras abejas realizan movimientos especiales en su comportamiento de pecoreo, para liberar el polen mediante la vibración de las flores, particularmente en plantas con anteras poricidas (Thorp, 2000).
De acuerdo con Rezende et al. (1996), la importancia de las abejas en la polinización se ha estudiado desde los tiempos de Mendel (1865) y de Darwin (1872) y, actualmente, el valor económico de la polinización es decenas de veces mayor que la producción de miel y cera. Kerr (1978) afirma que las plantas y las abejas que las polinizan poseen una interacción ecológica tan estrecha que, sin las plantas, la mayoría de ellas desaparecería en pocos meses y, sin abejas, muchas fanerógamas también lo harían en pocas generaciones.
Los estudios de abejas como visitantes florales en las regiones neotropicales son escasos, con objetivos limitados y metodologías variables (Ramalho et al., 1990). La flora apícola se ha caracterizado principalmente con base en observaciones de las visitas de las abejas melíferas africanizada y europea; ello ha contribuido significativamente al conocimiento de las plantas que les son útiles, pero existe un gran vacío con relación a las miles de especies silvestres que existen en el mundo, aunque se sabe que las abejas eusociales de la familia Apidae han desarrollado hábitos alimenticios generalistas (Ramalho et al., 1989; Marques-Souza et al., 1993; Rezende et al., 1996). La mayoría de las abejas nativas alimentan a sus crías con polen y néctar que colectan de las flores o, en pocos casos, de los nidos de otras abejas; a veces también lo hacen utilizando aceites vegetales (Vogel, 1969; Wcislo y Cane, 1996; Smith, 1999).
Para comprender las relaciones existentes entre abejas y plantas es preciso estudiar la dinámica de las poblaciones involucradas; realizar observaciones directas y trabajo experimental que permitan establecer el efecto de las abejas como visitantes florales. Existen especies vegetales generalistas, visitadas por gremios de insectos; otras son especialistas, como ocurre con varias orquídeas y la subfamilia Euglossinae (Feinsinger, 1987; Roubik, 1992; Vogel y Westerkamp, 1991, Marques-Souza et al., 1995). En muchos casos, el éxito reproductivo de las plantas está influenciado por las abejas, bien sea mediante la polinización, o de manera adversa cuando, al visitarlas, destruyen algunas flores o se roban las recompensas atractivas para los verdaderos polinizadores (Roubik, 1992). De todas formas, la alteración o ruptura de estos mutualismos pueden tener efectos en cascada, que afectan a otros organismos aunque no participen directamente de dichas relaciones (Aizen et al., 2002).
Debido a la importancia de las abejas no sólo en los procesos reproductivos de las plantas con flores, sino también en la conservación del equilibrio ecológico, se propuso realizar el estudio palinológico de las cargas polínicas colectadas por abejas visitantes de Aspilia tenella (Kunth) S. F. Blake, una especie muy abundante en fases sucesionales iniciales en la zona de influencia del embalse Porce II (Antioquia, Colombia). El análisis estadístico de las observaciones directas sobre el comportamiento de pecoreo de las abejas, combinado con el de los tipos polínicos encontrados en sus cargas de polen, permite comprender, al menos parcialmente, la importancia de las abejas en la relación plantapolinizador. Los resultados de este trabajo contribuyen al conocimiento de los hábitos alimenticios de las abejas en la zona de estudio, y pueden ser útiles para programas de recuperación de áreas degradadas en la región, mediante el uso de especies vegetales pioneras que sirven como fuente de alimento a muchos insectos.
]]>MATERIALES Y MÉTODOS
Esta investigación se realizó durante el primer semestre de 1998, en la zona de influencia del proyecto hidroeléctrico Porce II, la cual se sitúa al noroeste del departamento de Antioquia, Colombia, a 120 km de la ciudad de Medellín (Smith, 1999).
Según el sistema de clasificación propuesto por Holdridge, en el área del proyecto se encuentran tres zonas de vida: el bosque húmedo tropical (bh-T), el cual ocupa gran parte de los alrededores del embalse; la transición del bosque húmedo tropical al bosque muy húmedo premontano (bh-Tbmh-PM); y el bosque muy húmedo premontano (bmh-PM), localizado en las partes más altas de las vertientes del río Porce (Cano, 1995). La temperatura promedio anual es de 22,5 °C, la humedad relativa promedio mensual es del orden de 83% y la pluviosidad media anual de 3050 mm. El régimen de lluvias es bimodal: un período seco desde noviembre hasta marzo y uno lluvioso de abril a octubre, con una época seca entre julio y agosto (Smith, 1999).
La cobertura vegetal de la zona está compuesta principalmente por pastos que comienzan a ser reemplazados por rastrojos bajos; así mismo, se observan algunas áreas con rastrojos más altos y fragmentos de bosque localizados en la parte superior de colinas y vertientes y en las riberas del río Porce y algunos de sus afluentes (Smith, 1999).
Se estableció una parcela de 50 m x 100 m, en fase sucesional de pastizal a rastrojo bajo, en cuatro localidades: Santa Lucía (1100 msnm, N 06º, W 75º), Picardía (1037 msnm, N 06º, W 75°), Fosforito (945 msnm, N 06º, W 75º) y Cancana (941 msnm, N 06º, W 75°). En cada una de ellas se escogieron 24 individuos de A. tenella. Esta especie, conocida comúnmente como Botón de oro, crece en potreros con rastrojos y es abundante en bordes de fragmentos de bosque secundario en la región de Porce; es una hierba rastrera, con inflorescencias en capítulos de color amarillo. Florece a lo largo del año y es visitada por abejas en busca de polen y néctar (Smith, 1999). Los granos de polen son prolato-esferoidales, de ámbito circular, con espinas; 3-colporados, colpos largos; sexina espinosa (Aguilar-Sierra obs. pers.)
El comportamiento de pecoreo de las abejas se observó desde las 8:00 hasta las 16:00, por espacio de 15 min por planta, en cada una de las parcelas estudiadas. Este procedimiento se repitió durante cuatro días consecutivos para cubrir todas las parcelas. Se registraron las especies de abejas visitantes, número de individuos y tiempo total de visita. Como factores ambientales se midieron la temperatura y la humedad relativa a intervalos de dos horas. Estas variables se sometieron a análisis de covarianza factoriales para establecer si existían relaciones estadísticamente significativas entre ellas, mediante el programa Statgraphics Plus® v. 3.0 (Manugistics).
De cada una de las especies de abejas visitantes se capturaron entre dos y cuatro especímenes, los cuales se colocaron en tubos de vidrio con ácido acético glacial, debidamente tapados y rotulados. Mediante una jeringa los ejemplares colectados se lavaron con el mismo ácido y el material polínico obtenido, se sometió al proceso de acetólisis de Erdtman (1960) con las modificaciones descritas por Fonnegra (1989). Para el análisis polínico se prepararon cuatro placas permanentes con gelatina glicerinada, selladas con parafina, se marcaron y se incluyeron en la palinoteca del Laboratorio de Ecología y Conservación Ambiental (LECA) de la Universidad Nacional de Colombia, Sede Medellín.
Las especies de abejas fueron identificadas utilizando claves taxonómicas especializadas como las de Michener, McGinley y Danforth (1994) y Michener (2000) para los géneros y mediante la comparación con especímenes de la Colección de Abejas del Departamento de Agricultura de los Estados Unidos (USDA) en la Universidad del Estado de Utah, EEUU. Dichos ejemplares se encuentran depositados en el Museo Entomológico Francisco Luis Gallego (MEFLG) de la Universidad Nacional de Colombia, Sede Medellín.
El análisis palinológico comprendió la identificación de los tipos polínicos mediante la comparación con la colección de referencia de la zona de estudio y consulta de literatura especializada. Las preparaciones se observaron bajo un microscopio óptico OLYMPUS CH-30, con un aumento de 400X. Sobre las cuatro placas preparadas, se contabilizaron entre 1000 y 1500 granos de polen, según las recomendaciones de Vergeron (1964) para análisis de miel; los resultados se expresaron como porcentajes para estimar la abundancia relativa de cada tipo polínico.
]]> El paquete estadístico MVSP Plus® v. 3.0 se utilizó para aplicar el análisis de agrupamiento a los tipos polínicos encontrados en las cargas de las distintas especies de abejas, a las abejas mismas y a la riqueza de tipos polínicos colectados por ellas.
RESULTADOS
El anexo 1 presenta la abundancia relativa de los tipos polínicos hallados en las cargas polínicas de los 23 taxones de abejas capturados visitando a A. tenella. Dichas cargas muestran, en su totalidad (100%), la presencia de granos de polen de esta planta. No obstante, el tipo polínico de Piper aduncum fue encontrado en 21 de las cargas analizadas (91,3% del total).
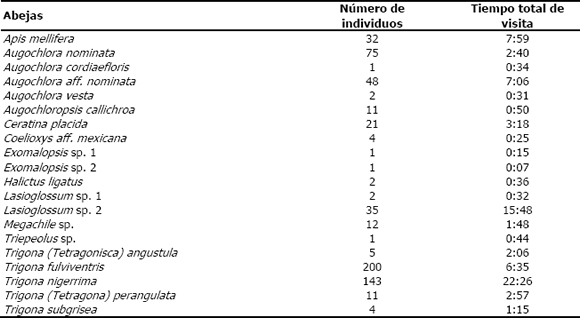
En la Tabla 1 se registra el número de individuos y la duración de la visita de las 20 especies de abejas observadas en A. tenella. Se pueden destacar dos meliponinos: Trigona fulviventris, la especie más abundante (200 individuos) y Trigona nigerrima, con el mayor tiempo total de visita (22:26).
Tabla 1. Especies de abejas observadas en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), visitando a Aspilia tenella, número de individuos y tiempo total de visita.
Las Tablas 2-4 contienen los resultados de los análisis de covarianza factoriales y de las pruebas de rangos múltiples; es importante señalar que Augochlora cordiae floris, Exomalopsis sp. 1, Exomalopsis sp. 2 y Triepeolus sp., representadas por un solo individuo (Tabla 1), se excluyeron de estos análisis.
Tabla 2. Nivel de significancia de las variables ambientales y las diferentes especies de abejas observadas en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), en relación a: 1 número de individuos; 2 tiempo total de visita.

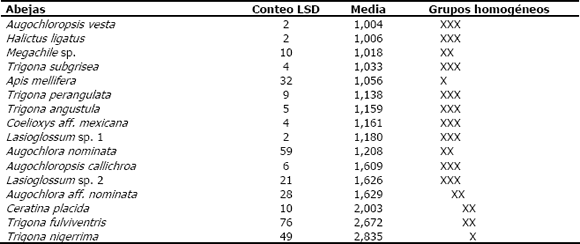
Tabla 4. Prueba de rangos múltiples que compara el tiempo promedio total de visita de las diferentes especies de abejas observadas en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), visitando a Aspilia tenella(X en líneas diferentes denotan diferencias estadísticas).
El histograma de la Figura 1, ratifica la importancia de A. tenella en la dieta de las abejas que la visitan. El análisis de agrupamiento para los tipos polínicos, representado en la Figura 2, permite señalar que A. tenella es un grupo polínico separado de los otros tipos identificados.
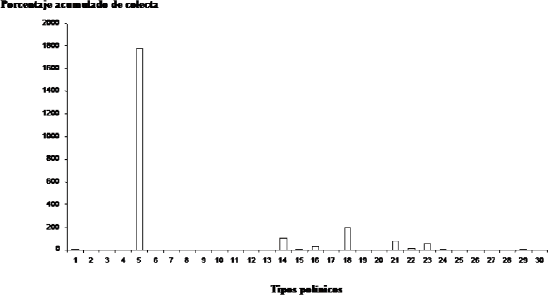
Figura 1. Porcentaje acumulado de los tipos polínicos colectados por las abejas capturadas en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), visitando a Aspilia tenella. Tipos polínicos: 5 Aspilia tenella; 18 Piper aduncum; 14 Miconia minuti flora; 21 Psidium guajava
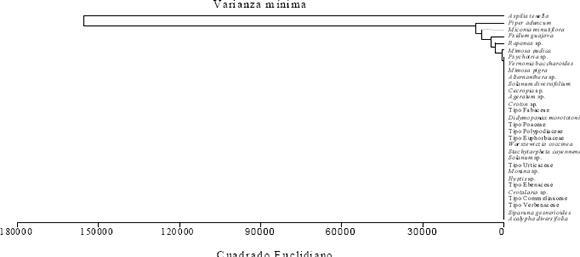
Figura 2. Análisis de agrupamiento para los tipos polínicos colectados por las abejas capturadas en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia), visitando a Aspilia tenella.
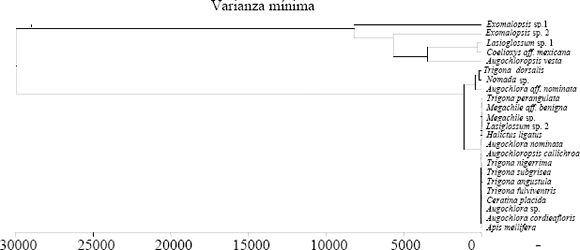
En el análisis de agrupamiento para las abejas capturadas visitando a A. tenella (Figura 3) se observan dos grupos: el primero, constituido por las abejas que pecorearon en las flores de esta especie hasta completar más de 70% de sus cargas, y el segundo, por aquellas que recolectaron porcentajes mayores de granos de polen de otras plantas.
]]>
El dendrograma de la Figura 4 permite distinguir tres grupos, conformados de acuerdo a la riqueza de tipos polínicos colectados por las abejas visitantes de A. tenella; dicha riqueza varió entre 2 y 14 tipos recolectados, los cuales reflejan la amplitud de sus nichos tróficos.
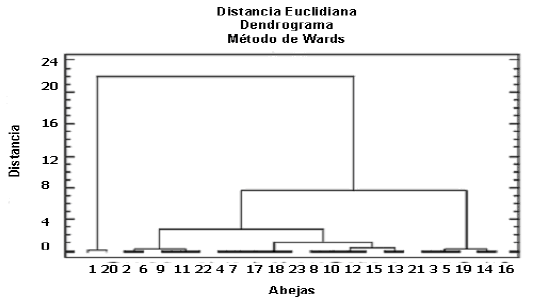
Figura 4. Análisis de agrupamiento para la riqueza de tipos polínicos de las cargas de abejas visitantes de Aspilia tenella en cuatro localidades de la zona de influencia del embalse Porce II (Antioquia, Colombia). (Abejas: Apis mellifera; 2 Augochlora cordiaefloris; 3 Augoclhora nominata ; 4 Augoclhora aff. nominata; 5 Augochlora sp.; 6 Augoclhoropsis callichroa; 7 Augochloropsis vesta; 8 Ceratina placida; 9 Coelioxys aff. mexicana; 10 Exomalopsis sp. sp. 1; 11 Exomalopsis sp. 2; 12 Halictus ligatus; 13 Lasioglossum sp. 1; 14 Lasioglossum sp. 2; 15 Megachile aff. benigna; 16 Megachile sp.; 17 Nomada sp.; 18 Trigona angustula; 19 Trigona dorsalis; 20 Trigona fulviventris; 21 Trigona nigerrima; 22 Trigona perangulata; 23 Trigona subgrisea).
DISCUSIÓN
En todas las cargas polínicas estudiadas se encontraron granos de polen de A. tenella (Anexo 1); ello evidencia la importancia de esta planta como fuente de alimento para las abejas que la visitan. Esto puede verificarse en la Figura 1, en la cual este tipo polínico alcanzó un porcentaje acumulado de colecta de 1777,2%; P. aduncum, que ocupa el segundo lugar, solamente acumuló un porcentaje de 197,2%, seguido de Miconia minutiflora, con 102,9% y de Psidium guajava, con 82,5%. En la literatura revisada no se encontró ningún reporte de visitas a A. tenella para colectar sus recursos florales. Girón (1996) describe la recolección de polen por A. mellifera en las espigas de P. aduncum; Ramalho et al. (1990) consideran a P. guajava y a otras especies de Miconia como fuentes de polen y néctar para la abeja melífera africanizada y para las abejas sin aguijón (Melipona y Trigona); y Absy et al. (1984) registran a Miconia y a Piper como géneros visitados por obreras de Meliponini para obtener polen, en la región del Amazonas Medio.
T. fulviventris y T. nigerrima son las especies visitantes más abundantes (200 y 143 individuos, respectivamente), seguidas de Augochlora nominata y Augochlora aff. nominata. Por su parte, a T. nigerrima le corresponde el mayor tiempo total de visita (22:26) y Lasioglossum sp. 2 ocupa el segundo lugar (15:48).
En términos generales, los géneros de especies parásitas (Coelioxys, Triepeolus) permanecieron menos tiempo en las flores, debido a que no requieren colectar recursos para aprovisionar sus nidos y solamente las visitan para su sostenimiento. Estas abejas pueden acarrear granos de polen por casualidad pero sólo las abejas que no son parasíticas, colectan y transportan polen entre las flores y lo llevan a sus nidos. Por su parte, en abejas solitarias y no parasíticas, como algunas especies de Augochloropsis, el tiempo de visita fue intermedio y en los géneros de abejas sociales (Trigona y Apis) fue superior, dada la necesidad de colectar una mayor cantidad de recursos florales no sólo para alimentar a las numerosas crías de sus colonias, sino también para almacenar alimentos que utilizan en épocas de escasez (Smith, 1999).
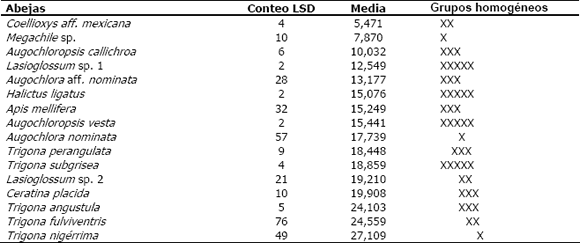
]]> El análisis de varianza para el número de individuos (Tabla 2) muestra una relación estadísticamente significativa entre las diferentes especies de abejas (nivel de significancia 0,0000). También hay una covariación significativa con la temperatura y la humedad relativa (nivel de significancia 0,0067 y 0,0001, respectivamente); es decir, ambas variables ambientales afectan el número de individuos de cada una de las especies de abejas visitantes. La prueba de rangos múltiples correspondiente (Tabla 3) muestra que T. nigerrima, T. fulviventris y Ceratina placida son las especies con los mayores valores promedio para el número de individuos; entre estas no existen diferencias estadísticamente significativas, pero sí las hay, con relación a las demás especies.De acuerdo con el análisis de varianza para el tiempo total de visita (Tabla 2), hay una relación estadísticamente significativa entre las diferentes especies de abejas visitantes de A. tenella (nivel de significancia 0,0000) y la duración de la visita sólo es afectada significativamente por la temperatura (nivel de significancia 0,0060). La prueba de rangos múltiples de la Tabla 4, confirma que Trigona nigerrima es la especie con un tiempo promedio total de visita mayor; sin embargo, ella no difiere estadísticamente de Lasioglossum sp. 1, Halictus ligatus, Augochloropsis vesta, Ceratina placida, Trigona subgrisea, T. angustula y T. fulviventris.
Los resultados anteriores permiten concluir que T. nigerrima, T. fulviventris y Ceratina placida conforman un gremio de abejas que muestra los mayores valores, sin diferencias estadísticamente significativas entre sí, con respecto al número de individuos visitantes y al tiempo total de visita. Esto les confiere una mayor probabilidad de ser el grupo de abejas que polinizó a A. tenella durante el tiempo en el que se realizó el presente estudio.
El análisis de agrupamiento para los 30 tipos polínicos identificados (Figura 2), permite observar que A. tenella constituye un grupo separado de los demás; en efecto, la abundancia relativa del polen de esta especie es superior a 90% en la mayoría de las cargas polínicas estudiadas. Dentro del otro grupo, que reúne a los 29 tipos polínicos restantes, se destacan en su orden: P. aduncum, M. minutiflora, Psidium guajava, Rapanea sp. y Mimosa pudica, plantas que pueden considerarse como importantes fuentes alternativas de polen para muchos de los taxones de abejas visitantes de A. tenella (Anexo 1).
En la Figura 3 pueden diferenciarse dos grupos principales de abejas visitantes de A. tenella. Uno de ellos incluye a 18 de las 23 especies de abejas capturadas, las cuales recolectaron granos de polen de la planta estudiada con una abundancia relativa superior a 77,7%; dentro de este grupo 15 especies presentan una alta constancia floral hacia A. tenella debido a que sus cargas polínicas contienen más de 90% de granos de polen de esta planta. El segundo grupo está conformado por cinco especies de abejas cuyas cargas muestran porcentajes mayores de otros tipos polínicos: Exomalopsis sp. 1 (90,4% de M. minutiflora); Lasioglossum sp. 1 y Coelioxys aff. mexicana (77,4% y 64,1% de P. aduncum, respectivamente); Exomalopsis sp. 2 (67,1% de P. guajava) y Auglochloropsis vesta (55,5% de Rapanea sp.).
Con relación al número de tipos polínicos presentes en las cargas de las abejas (Anexo 1), el análisis de agrupamiento para la riqueza de los mismos (Figura 4) muestra tres grupos: el primero abarca a T. fulviventris y a A. mellifera con 14 y 13 tipos polínicos, respectivamente; son especies polilécticas, con nichos tróficos amplios. El segundo grupo, con una riqueza intermedia, comprende 16 especies de abejas con un número de tipos polínicos que oscila entre cuatro y ocho. El tercer grupo, con la menor riqueza de tipos polínicos, incluye a Augochlora nominata, Augochlora sp., Trigona dorsalis, Lasioglossum sp. 2 y Megachile sp., cinco abejas nativas que visitaron entre dos y tres especies de plantas; sus nichos tróficos son más reducidos.
Vale la pena llamar la atención sobre el alto número de tipos polínicos colectados por A. mellifera, una especie introducida, considerada con una alta constancia floral en la zona templada (Roubik, 1992). Sin embargo, este mismo autor, presume que una constancia floral estricta debe ser rara entre abejas tropicales y Slaa et al. (1998) proponen que las estrategias de pecoreo óptimas difieren entre las zonas ecuatoriales y las templadas, de manera que las abejas de climas tropicales, generalmente, son menos constantes que las abejas de climas templados.
Estos resultados también están de acuerdo con los de Rezende et al. (1996), quienes estudiaron el grado de infidelidad a la misma especie de flores, en tres especies de meliponinos: se encontró que Melipona seminigra, una abeja amazónica, tiene una tasa de infidelidad floral casi tres veces mayor a la de dos especies del mismo género en el centro de Brasil. Cuando compararon estos resultados con los informes de un alto porcentaje de infidelidad para Apis, en Asia tropical, proponen la hipótesis de que M. seminigra y las especies de Apis tropicales deben haber sido seleccionadas para una mayor infidelidad floral, debido a que en estas latitudes, las plantas en flor de una misma especie están a distancias mayores, y de esta manera, una obrera tiene más posibilidades de encontrar varias especies en floración simultánea y colectar el polen o el néctar en más de un tipo de flor; de no hacerlo, estaría colectando menos alimento por unidad de tiempo o espacio recorrido durante el vuelo.
AGRADECIMIENTOS
Este trabajo fue posible gracias a la financiación de las Empresas Públicas de Medellín (EPM E.P.S). Agradecemos muy especialmente a Charles D. Michener por la lectura crítica del manuscrito; a John Jairo Ramírez y a Alvaro Lema por la asesoría en el análisis estadístico de los resultados; y a la Universidad Nacional de Colombia, Sede Medellín, por apoyar a los autores durante esta investigación.
]]>BIBLIOGRAFÍA
Absy, M.L., J.M.F. Camargo, W.E. Kerr y I.P. Miranda. 1984. Espécies de plantas visitadas por Meliponinae (Hymenoptera, Apoidea) para coleta de pólen na regiao do Médio Amazonas. Rev. Brasil. Biol. 44(2): 227-237. [ Links ]
Aizen, M., D. Vásquez y C. Smith. 2002. Historia natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Rev. Chil. Hist. Nat. 75(1): 79-97. [ Links ]
Cano, J.A. 1995. Utilización de un sistema de información geográfico (SIG) en la elaboración de un plan de uso y manejo de la cobertura vegetal, Trabajo de grado. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 210 p. [ Links ]
Erdtman, G. 1960. The acetolysis method. A revised description. Svensk Bot. Tidskr. 54: 561-564. [ Links ]
Fonnegra, R. 1989. Métodos de estudio palinológico. Centro de Investigaciones Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia. Medellín. 57 p. [ Links ]
Feinsinger, P. 1987. Approaches to nectarivore plant interactions in the new world. Rev. Chil. Hist. Nat. 60: 285-319. [ Links ]
Girón, M. 1996. Melitopalinología. Recolección de polen y néctar por Apis mellifera en algunas especies de plantas silvestres y cultivadas del municipio de Salgar (Antioquia). ColcienciasUniversidad del Quindío, Armenia. 83 p. [ Links ]
Kerr, W.E. 1978. Papel das abelhas sociais na Amazônia. p. 1-11. En: Memorias. Annais Simp. Intern. APIMONDIA sobre Apicultura em clima quente. Florianópolis. [ Links ]
Marques, A.C., M.L. Absy, P.A. Condé e H. Coelho. 1993. Dados da obtençao do pólen por opérarias de Apis mellifera no município de Ji-Paraná (RO), Brasil. Acta Amazonica 23(1): 59-76. [ Links ]
Marques, A.C., M.L. Absy, W.E. Kerr e F.J. Agilera. 1995. Pólen coletado por duas espécies de Meliponíneos (Hymenoptera: Apidae) da Amazônia. Rev. Brasil. Biol. 55 (4): 855-864. [ Links ]
Michener, C.D. 2000. The bees of the world. John Hopkins University Press, Baltimore. 913 p. [ Links ]
Michener, C.D., R. McGinley and B. Danforth. 1994. The bee genera of North and Central America (Hymenoptera: Apoidea). Smithsonian Institution Press, Washington D.C. 209 p. [ Links ]
Ramalho, M., A Kleinert and V.L. Fonseca. 1989. Utilization of floral resources by species of Melipona (Apidae, Meliponinae): floral preferences. Apidologie 20(3): 185-195. [ Links ]
Ramalho, M., A. Kleinert and V.L. Fonseca. 1990. Important bee plants for stingless bees (Melipona and Trigonini) and africanized honeybees (Apis mellifera) in neotropical habitats: a review. Apidologie 21(5): 469-488. [ Links ]
Rezende, E.B.S., M.L. Absy e W.E. Kerr. 1996. Infidelidade à mesma espécie de flores em três espécies de abelhas (Apidae: Meliponinae). An. Soc. Entomol. Brasil. 25(3): 549-552. [ Links ]
Roubik, D. 1992. Ecology and natural history of tropical bees. Cambridge University, New York. 520 p. [ Links ]
Slaa, E.J., A. Cevaal and M.J. Sommeijer. 1998. Floral constancy in Trigona stingless bees foraging on artificial flower patches: a comparative study. Journal of Apicultural Research 37(3): 191-198. [ Links ]
Smith, A.H. 1999. Abejas (Hymenoptera: Apoidea) de la zona de influencia del embalse Porce II (Antioquia, Colombia). Tesis Magister en Entomología. Facultad de Ciencias. Universidad Nacional de Colombia. Medellín. 341 p. [ Links ]
Thorp, R.W. 2000. The collection of pollen by bees. Plant Syst. Evol. 222(1-4): 211-223. [ Links ]
Vergeron, P. 1964. Interprétation statistique des résultants en matière danalyse pollinique des miels. Ann. Abeille 7: 349-364. [ Links ]
Vogel, S. 1969. Flowers offering fatty oil instead of nectar. En: Memorias. XIth Internat. Bot. Congr. Seattle. [ Links ]
Vogel, S. and C. Westerkamp. 1991. Pollination: An integrating factor of biocenoses. pp. 159-170. En: Seitz, A. and V. Loeschcke. Species conservation: A population-biological approach. Birkhäuser, Basel. [ Links ]
Wcislo, W.T. and J.H. Cane. 1996. Floral resource utilization by solitary bees (Hymenoptera: Apoidea) and exploitation of their stored foods by natural enemies. Ann. Rev. Entomol. 41: 257-286. [ Links ] ]]>