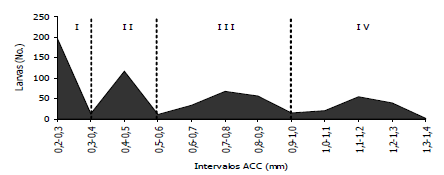
Figura 1. Distribución de frecuencias del ancho de la cápsula cefálica de larvas sanas de Tecia solanivora bajo condiciones de laboratorio. (I: Instar 1, II: Instar 2, III: Instar 3, IV: Instar 4)
EFFECT OF GRANULO VIRUSES INFECTION ON THE DEVELOPMENT OF Tecia solanivora (Lepidoptera: Gelechiidae)
Carlos Espinel Correal1; Alba Marina Cotes Prado2 y Laura Villamizar Rivero3
1 Investigador. Corporación Colombiana de Investigación Agropecuaria-CORPOICA. Kilómetro 14 vía Mosquera. Bogotá, Colombia. <cespinel@corpoica.org.co>
2 Investigadora. Corporación Colombiana de Investigación Agropecuaria-CORPOICA. Kilómetro 14 vía Mosquera. Bogotá, Colombia. <amcotes@corpoica.org.co>
3 Investigadora. Corporación Colombiana de Investigación Agropecuaria-CORPOICA. Kilómetro 14 vía Mosquera. Bogotá, Colombia. <lvillamizar@corpoica.org.co>
Recibido: Abril 16 de 2008; Aceptado: Marzo 17 de 2009.
Palabras claves: Baculovirus, papa.
Abstract. Biological control of Tecia solanivora by the granulovirus of Phthorimaea operculella from Peru is one of the main components for its management. Corpoica has five native isolates of granulovirus obtained from T. solanivora larvae. Taking into account that these isolates could be adapted to the insect and could have a differential infection expression as compared to Peruvian isolate, the objective of the present work was to study the effect of this virus on the insect development. Initially, the duration and development stages of healthy larvae were determined under laboratory conditions. Width cephalic capsule was determined (WCC) and symptoms of illness were described by evaluating neonate larvae after 2, 4, 6, 8, 10, 12, 14, 16, 20 and 22 days of emergency. Growth of larvae was in accordance with the Dyar law, since the cephalic capsule growth of successive instars followed a regular geometric progression. Analysis of frequency distribution of WCC determined four larval instars in healthy larvae. Delay and overlap of larvae instars were caused by viral infection. Differences in the expression of signals and symptoms among native viruses, and between these and granulovirus of Peru were observed, suggesting differences in the isolates pathogenicity, which could be related with genotypic differences between the viruses.
Key words: Baculovirus, potato.
La polilla guatemalteca de la papa Tecia solanivora es la plaga de mayor importancia económica de este cultivo en Colombia. Su ataque causa grandes pérdidas económicas en la producción de papa y reducción en su valor comercial ya que los tubérculos severamente afectados no se pueden utilizar para semilla ni para consumo humano o animal. El manejo de este insecto se ha basado en el uso indiscriminado de productos químicos registrados por el ICA pertenecientes a la categoría toxicológica I y II, los cuales han resultado insuficientes para controlarlo, además de generar alta contaminación ambiental debido a que son altamente tóxicos (SINAIPA, 2002). Considerando dichas limitantes ha surgido la necesidad de desarrollar alternativas de control biológico, como el uso de los virus entomopatógenos que aseguren el manejo sostenible y eficiente de la polilla y que puedan ser incluidas dentro de programas de manejo integrado de esta plaga. Dentro de estos virus se destaca el uso de granulovirus pertenecientes a la familia Baculoviridae, ya que representa el grupo más estudiado para el control de este grupo de insectos.
Con el fin de responder a la emergencia sanitaria causada por T. solanivora en la década de los 90, se introdujo y se multiplicó masivamente en Colombia un aislamiento de un virus de la granulosis de Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae) proveniente del Perú. En el año 2000, el Laboratorio de Control Biológico de CORPOICA se enfrentó al reto de fabricar el producto a base de este baculovirus a escala semicomercial. Teniendo en cuenta que dicho aislamiento viral era una cepa foránea aislada de un hospedero diferente a T. solanivora que es el objeto de control en Colombia, se realizó una colección de aislamientos nativos de granulovirus obtenidos a partir de T. solanivora, de diferentes zonas productoras de papa en Colombia, encontrándose cinco potenciales aislamientos denominados VG 001, VG 002, VG 003, VG 004 y VG 005, provenientes de Túquerres (Nariño), Chocontá (Cundinamarca), Funza (Cundinamarca), Cármen de Carupa (Cundinamarca) y Chitagá (Norte de Santander), respectivamente (Villamizar et al., 2005).
Estos aislamientos nativos presentan alto potencial para el desarrollo de un bioplaguicida dirigido al control del insecto bajo condiciones de campo. Sin embargo, debe tenerse en cuenta que el desarrollo de este tipo de productos requiere del cumplimiento de etapas biológicas y tecnológicas que garanticen la obtención de un producto seguro eficaz y confiable (Villamizar et al., 2005). De tal manera, es necesario conocer a fondo las características biológicas del insecto blanco, del microorganismo y la compleja interacción patógeno-insecto, tales como su modo de acción, virulencia y la influencia que el microorganismo pueda tener sobre la biología del insecto. Por tal razón, los objetivos del presente trabajo fueron determinar el efecto de los granulovirus nativos en el desarrollo larval de Tecia solanivora y describir los signos de la infección de los granulovirus sobre larvas de la polilla guatemalteca de la papa.
MATERIALES Y MÉTODOS
]]> El trabajo se realizó en el laboratorio de Control Biológico de CORPOICA, ubicado en Mosquera, Cundinamarca, (Colombia) a una temperatura de 23ºC ± 3 y una humedad relativa de 65% ± 10.Desarrollo de los signos de infección. Con el propósito de determinar el efecto de la infección sobre el desarrollo de T. solanivora se llevó a cabo un bioensayo bajo condiciones de laboratorio. Para tal fin, previamente se establecieron los estadios de desarrollo de las larvas sin infección viral bajo las condiciones experimentales. Se tomó como referencia la ley de Dyar, la cual postula que la mayoría de las estructuras del cuerpo de los insectos incrementan su tamaño linealmente con relación a un radio constante de dicha estructura (crecimiento isométrico) (Dyar, 1890).
Posturas de Tecia solanivora depositadas en un corte de papel toalla procedentes de una cría del laboratorio de Entomología del C.I. Tibaitatá-CORPOICA, se incubaron y se esperó la emergencia de las larvas. Una vez emergidas, se colocaron 15 larvas sobre cada unidad experimental, la cual consistió en un tubérculo de papa parda pastusa Solanum tuberosum subespecie andigena de 80 g aproximadamente; cada tubérculo se colocó en el interior de una cubeta plástica de 16 onzas con tapa; para un total de 60 unidades experimentales por tratamiento.
Posteriormente, a partir del segundo día y cada dos días hasta el día 20, se escogieron al azar seis cubetas y se realizó un muestreo destructivo de los tubérculos con el fin de extraer las larvas presentes en cada uno. Las larvas se ubicaron sobre una lámina portaobjeto en vista dorsal a las que se les midió el ancho de la cápsula cefálica (ACC) (distancia entre genas) por medio de un micrómetro adaptado a un microscopio óptico con un aumento de 4 x. Se tomó como referencia del día cero las larvas recién emergidas. Con los resultados obtenidos se realizó una distribución de frecuencias con el fin de determinar los ínstares larvales y el rango del ancho de la cápsula cefálica.
Una vez establecidos los ínstares larvales bajo las condiciones experimentales se determinó el efecto de los granulovirus sobre el desarrollo larval de T. solanivora. A posturas del insecto mantenidas en el laboratorio, se les aplicó por goteo mediante una jeringa, una suspensión viral de 3 mL a la concentración de 105 CI•mL-1 (CI: Cuerpos de Inclusión) de cada uno de los virus nativos. Cada tratamiento correspondió a un aislamiento viral codificado como VG 001, VG 002, VG 003, VG 004 y VG 005, provenientes de Túquerres, Chocontá, Funza, Cármen de Carupa y Chitagá, respectivamente y el virus de la granulosis aislado de P. operculella codificado como Perú, se utilizó como cepa de referencia; asimismo, se tuvo en cuenta un testigo absoluto, con posturas de T. solanivora sin aplicación viral. Después de inoculadas las posturas se dejaron secar a la temperatura del cuarto de bioensayos y se esperó hasta la emergencia de las larvas.
Al observarse la emergencia de las larvas, con ayuda de un pincel Nº 0, se ubicaron 15 individuos en la superficie de cada tubérculo, representando la unidad experimental con tres repeticiones como se indicó anteriormente. Teniendo en cuenta los resultados obtenidos en el ensayo del establecimiento de los estadios de desarrollo con larvas sin infección, se decidió hacer el muestreo de los tratamientos en los días 7, 12, 17 y 20, en los cuales, el mayor porcentaje de larvas se encontrarían en primero, segundo, tercer y cuarto estadio de desarrollo, respectivamente. Se tomaron tres recipientes por tratamiento, incluido el testigo absoluto, y se llevó a cabo un muestreo destructivo con el fin de medir el ACC tal como se explicó anteriormente. El día cero (0) de evaluación se tomó como el día de la emergencia de las larvas. Los resultados obtenidos se sometieron a una distribución de frecuencias, con el fin de determinar los ínstares larvales con la infección viral y se comparó su desarrollo con las larvas del testigo.
Paralelamente, en cada día de evaluación se llevó a cabo una descripción macroscópica de los signos de infección producidos por cada uno de los aislamientos virales en las larvas de T. solanivora.
RESULTADOS Y DISCUSIÓN
Con el análisis de distribución de frecuencias del ancho de la cápsula cefálica con un intervalo de 0,2 mm, se estableció la escala de ACC para cada uno de los ínstares de la polilla guatemalteca sin infección viral (Tabla 1) encontrándose cuatro grupos definidos y sin traslape sugiriendo la formación de cuatro instares larvales en T. solanivora, evidenciado por la presencia de igual número de picos independientes (Figura1). Este número de ínstares larvales de T. solanivora ha sido registrado por Niño y Notz (2000), quienes establecieron cuatro grupos de población larval claramente definidos bajo las condiciones experimentales del estudio. Aunque otros estudios similares han mencionado pequeñas diferencias en los intervalos; esto podría deberse a cambios de las condiciones ambientales en cada estudio, a condiciones nutricionales variables por las diferencias en el sustrato alimenticio y a variaciones en el estado fisiológico de las larvas provenientes de una cría (Llanderal, 1993).
]]> Tabla 1. Valores del ancho de la cápsula cefálica (ACC) de larvas sanas de Tecia solanivora distribuidos según el estadio de desarrollo.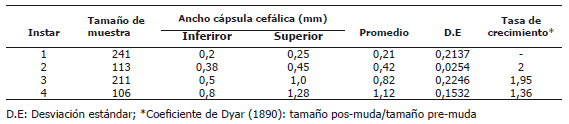
Figura 1. Distribución de frecuencias del ancho de la cápsula cefálica de larvas sanas de Tecia solanivora bajo condiciones de laboratorio. (I: Instar 1, II: Instar 2, III: Instar 3, IV: Instar 4)
Al establecer el tiempo de duración de cada ínstar se observaron larvas de primer instar desde el día cero (100%) hasta el día 10 (1,9%), larvas de segundo ínstar desde el día 6 (85,4%) hasta el día 16 (2,4%), larvas de tercer ínstar desde el día 10 (69,2%) hasta el día 20 (5,6%) y de cuarto ínstar desde el día 14 (39%) hasta el día 20 (94,3%). En el desarrollo de las larvas sanas se observaron algunos individuos con un desarrollo más lento, evidenciado por una presencia inferior al 10%, especialmente del primer ínstar en los días 8 y 10, en el segundo ínstar en los días 14 y 16 y en el tercer ínstar al día 20. Esta población de primero, segundo y tercer instar se caracterizó por su crecimiento más lento, el cual podría atribuirse a las condiciones de muestreo establecidas, debido a que por ser muestreos destructivos, en cada día de medición se utilizaron individuos completamente diferentes. Cada uno de estos individuos podría presentar diferencias genéticas, de edad y de condiciones nutricionales, puesto que provenían de varias progenitoras que pudieron poner sus huevos en el mismo sustrato de oviposición (papel toalla) a pesar de ser originarias de una misma cría mantenida en laboratorio. Heeley et al. (2003) encontraron que en el ciclo de Rhyacionia buoliana (Lepidoptera:Tortricidae) se presentan algunas variaciones en la distribución de frecuencias de los estados larvales, atribuidas a diferencias en la nutrición y fisiología del insecto.
Desarrollo de signos de infección de los granulovirus nativos sobre larvas de Tecia solanivora. Al realizarse la infección de las larvas con los aislamientos provenientes de Funza, Chocontá, Carmen de Carupa y Chitagá, se observó un mayor retraso y traslape en el desarrollo de los ínstares larvales con estos aislamientos. Tal es el caso del primer ínstar, en el que el desarrollo de las larvas sanas (testigo) presentó una frecuencia del 100% hasta el día 4 y entre el 2 y el 14% en los días 6, 7 y 8. Estas frecuencias contrastaron con las observadas en las larvas infectadas, debido a que en el día 7, las larvas de primer ínstar estuvieron presentes en un 66,6% con el aislamiento de Funza, 69,2% con el aislamiento de Carmen de Carupa, 50% con el aislamiento de Chocontá y 60% con el aislamiento proveniente de Chitagá (Figuras 3 a 6). En las larvas infectadas con el aislamiento proveniente de Túquerres, no se observó una frecuencia tan alta en primer ínstar en el día 7 (20%), sin embargo, cinco días después (día 12), aún había una frecuencia del 22,2% de larvas en primer ínstar (Figura 7). En comparación con la mayoría de los tratamientos incluyendo el testigo donde ya no había presencia de este ínstar al día 12 de evaluación. En las larvas infectadas con el aislamiento proveniente del Perú, no se observó retraso en el primer ínstar debido a que al día 7 la frecuencia fue del 10% comparable con las larvas del testigo (Figura 8), lo cual sugiere que el proceso de infección primaria para este aislamiento es más lento y por ende los signos de infección no se evidenciaron tan rápido como se constató con los otros aislamientos.
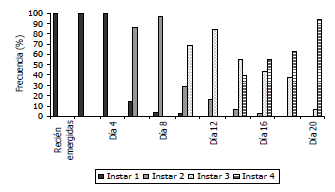
Figura 2. Comportamiento de los instares larvales de larvas sanas Tecia solanivora bajo condiciones de laboratorio.
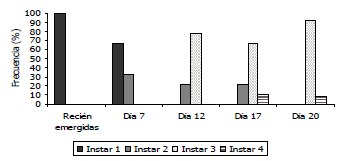
Figura 3. Desarrollo larval de Tecia solanivora ante la infección de un aislamiento nativo del virus de la granulosis proveniente de Funza - Cundinamarca, Colombia (VG003).
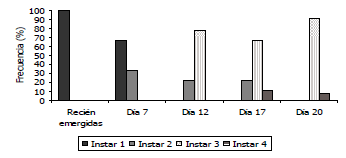
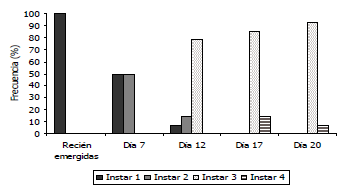
Figura 5. Desarrollo larval de Tecia solanivora ante la infección de un aislamiento nativo del virus de la granulosis proveniente de Chocontá - Cundinamarca, Colombia (VG002).
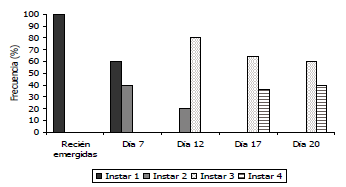
Figura 6. Desarrollo larval de Tecia solanivora ante la infección de un aislamiento nativo del virus de la granulosis proveniente de Chitagá - Norte de Santander, Colombia (VG005).
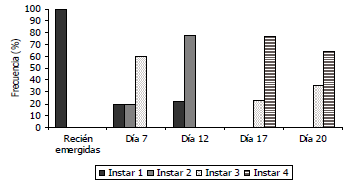
Figura 7. Desarrollo larval de Tecia solanivora ante la infección de un aislamiento nativo del virus de la granulosis proveniente de Túquerres - Nariño, Colombia (VG001).
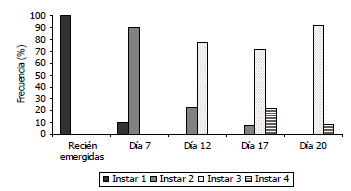
Figura 8. Desarrollo larval de Tecia solanivora ante la infección de un aislamiento nativo del virus de la granulosis proveniente del Perú.
Sin embargo, en las larvas infectadas con los aislamientos de granulovirus se observó de nuevo un retraso en el desarrollo el cual fue más evidente en el tercer ínstar. Los primeros individuos de tercer ínstar se observaron hacia el día 12, tal como en el testigo, con frecuencias de 77,8; 69,2; 78,5; 80 y 77,7% para las larvas infectadas con los aislamientos provenientes de Funza, Carmen de Carupa, Chocontá, Chitagá y Perú, respectivamente, en cambio con el aislamiento de Túquerres se presentó en el día 7 con una frecuencia del 60%. Para todos los aislamientos la evidencia del retraso en el desarrollo se observó hacia el día 20, debido a que mientras en el testigo la frecuencia era sólo del 5,6%, en las larvas infectadas la frecuencia fue del 91,6; 91,6; 92,8; 60; 35,7 y 91,6%, con los aislamientos de Funza, Carmen de Carupa, Chocontá, Chitagá, Túquerres y Perú, respectivamente (Figuras 3 a 8).
En el cuarto ínstar larval también se observaron diferencias entre las larvas sanas e infectadas con los aislamientos virales, debido a que en las larvas sanas, este ínstar comenzó hacia el día 14 con una frecuencia de 39%, mientras que en todos los tratamientos con granulovirus el cuarto ínstar comenzó al día 17 con porcentajes de frecuencia de 11,1; 14,3; 35,8; 77 y 21,4%, para los aislamientos provenientes de Funza, Chocontá, Chitagá, Túquerres y Perú, respectivamente. Con el aislamiento proveniente de Carmen de Carupa, el cuarto ínstar comenzó al día 12 con una frecuencia del 32% de las larvas.
Hacia el día 20 de evaluación, la mayoría de la población de las larvas testigo (94,3%) estuvo en el cuarto ínstar larval, mientras que para ese mismo día en las larvas infectadas con los diferentes aislamientos, el cuarto ínstar estuvo presente con una frecuencia del 8,4; 8,4; 7,1; 40; 64,5 y 8,4%, en los aislamientos provenientes de Funza, Carmen de Carupa, Chocontá, Chitagá, Túquerres y Perú, respectivamente (Figuras. 2 a 8). Cabe resaltar, que en el día 20 se presentaron pupas provenientes del tratamiento de larvas sanas, lo que indicaría que la duración del estado larval sería de 20 días; mientras que en los tratamientos con infección viral aún la mayor frecuencia estuvo en el tercer ínstar. Esto indicaría que el virus tiene un efecto sobre el desarrollo larval evidenciado por un desorden en la presencia de los ínstares, afectando la duración de los mismos y prolongándose los últimos ínstares sin llegar a formar el estado de pupa. Aunque esta evaluación se realizó hasta el día 20, sería recomendable llevarla a cabo con una mayor duración, con el fin de evidenciar si las larvas infectadas logran mudar al estado de pupa. Asimismo, se recomienda evaluar con intervalos de días más cortos, diariamente o máximo cada dos días, con el fin de establecer la duración exacta de los instares de desarrollo.
El traslape en la duración de los estados de desarrollo de las larvas infectadas con los diferentes virus y el retraso en su desarrollo, indicaría que los granulovirus tienen un efecto sobre el desarrollo larval generando una mayor duración en su ciclo de vida. Niño y Notz (2000), encontraron que las larvas infectadas por un virus nativo de T. solanivora presentaron un traslape, debido a que no hubo presencia de grupos discontinuos. Esto se reflejó en un incremento en la duración de su estado larval.
El retraso en el desarrollo de las larvas infectadas con los diferentes aislamientos de granulovirus puede tribuirse al efecto negativo que tiene el virus en los hábitos alimenticios y en la expresión hormonal de las larvas durante el cambio sucesivo de ínstares. Según Muscardi (1999), una característica de la infección ocasionada por baculovirus es el retraso del desarrollo y la reducción de la capacidad alimenticia de las larvas; estas características se pueden deber a la expresión de genes que codifican para proteínas extrañas en el genoma de los baculovirus, tales como los genes que codifican para toxinas específicas para hormonas de insectos como la esterasa juvenil y para enzimas como la UDP-glucosiltransferasa. De igual manera, este comportamiento fue registrado por Subramanian et al. (2005) quienes afirmaron que el proceso de infección viral tiene una influencia negativa en la expresión de los genes del hospedero y en la producción de proteínas. Asimismo, Burand y Park (1992) establecieron que en el desarrollo de las larvas infectadas con virus no ocurren los procesos de apolisis y hay un bloqueo en la formación de pupas, debido a la expresión del antígeno de proliferación celular, el cual detiene el crecimiento de las larvas y ayuda a la replicación viral.
Signos del desarrollo de la infección por granulovirus. Al realizar el muestreo destructivo de cada uno de los tratamientos se hizo la descripción de los signos de la infección por granulovirus en las larvas.
Al día 7 después del tratamiento con todos los aislamientos virales, se observaron larvas sin signos evidentes de la infección por granulovirus. Las larvas tenían un aspecto translúcido, en el cual se podían ver los órganos internos del insecto. Hacia el día 12 comenzaron a presentarse "parches" o aglomeraciones granulares blancas en los últimos segmentos abdominales, similar a la leche cortada (Figura 9). Hacia el día 17 post infección, las aglomeraciones blancas se presentaron en la totalidad de los segmentos abdominales y torácicos, dando una coloración blanca uniforme en las larvas. En el día 20 se observó la coloración rosada intensa en el dorso de las larvas provenientes del testigo, común en los últimos ínstares de larvas sanas de esta especie, aunque en algunas las larvas infectadas se observó una pigmentación rosado claro debido a la presencia de la infección viral evidenciada por el color blanco de los cuerpos de inclusión de los virus de la granulosis (Figura 9).
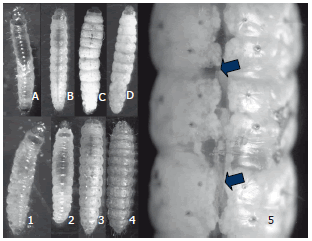
Figura 9. Signos del desarrollo de la infección de granulovirus sobre el estado larval de Tecia solanivora. A. Larva enferma día 7. B. Larva enferma día 12. C. Larva enferma día 17. D. Larva enferma día 20. 1. Larva sana día 7. 2. Larva sana día 12. 3. Larva sana día 17. 4. Larva sana día 20. 5. Detalle de aglomeraciones de cuerpos de inclusión de granulovirus sobre el 5º, 6º y 7º segmento abdominal.
CONCLUSIONES
AGRADECIMIENTOS
Los autores agradecen al programa ECOS de Francia, al ICFES, ICETEX y COLCIENCIAS, por su apoyo financiero para la realización de este trabajo.
BIBLIOGRAFÍA
Burand, J.P. and E.J. Park.1992. Effect of nuclear polyhedrosis virus infection on the development and pupation of gypsy moth larvae. Journal of Invertebrate Pathology. 60(2): 171-175. [ Links ]
Caballero, P., M. López y T. Williams. 2001. Los baculovirus y sus aplicaciones como bioinsecticidas en el control biológico de plagas. Editorial Phytoma, España. 518 p. [ Links ]
Dyar, H.G. 1890. The number of molts of lepidopterous larvae. Psyche, Camb 5: 420-422. [ Links ]
SINAIPA (Sistema Nacional de Información de Papa). 2002. Boletín de la Papa 4(12): 1-19 [ Links ]
Heeley, T., R.I. Alfaro, L. Humble and W. Strong. 2003.Distribution and life cycle of Rhyacionia buoliana (Lepidoptera:Tortricidae) in the interior of British Columbia. Journal of the Entomological Society of British Columbia 100:19-25. [ Links ]
Llanderal, C.1993. Definición de ínstares larvales de Phthorimaea operculella por medición de la cápsula cefálica. Agrociencia. Serie Protección Vegetal 4(2): 179-186. [ Links ]
Muscardi, F. 1999. Assessment of the application of baculoviruses for control of Lepidoptera. Annual Review of Entomology 44: 257-289. [ Links ]
Niño, L. y A. Notz. 2000. Desarrollo y sintomatología de las larvas de la polilla de la papa Tecia solanivora (Povolny) 1973 (Lepidoptera: Gelechiidae) infectadas por un virus granulosis nativo. Boletín Entomológico Venezolano 15(1): 29-38. [ Links ]
Subramanian, S., R. Rabindra, S. Palaniswamy, N. Sathiah and B. Rajasekaran. 2005. Impact of granulovirus infection on susceptibility of Spodoptera litura to insecticides. Biological Control 33: 165-172. [ Links ]
Villamizar, L., J. Zeddam, C. Espinel y A. Cotes. 2005. Implementación de técnicas de control de calidad en el proceso productivo de un bioplaguicida a base del granulovirus de Phthorimaea operculella PhoGV. Revista Colombiana de Entomología 31(2): 127-132 [ Links ] ]]>