Effects of light intensity on the morphology and CAM photosynthesis of Vanilla planifolia Andrews
Efectos de la intensidad lumínica sobre la morfología y la fotosíntesis CAM de Vanilla planifolia Andrews
M. Claudia Díez1, Flavio Moreno1 and Edison Gantiva1
1 Facultad de Ciencias Agrarias. Universidad Nacional de Colombia. AA 1779. Medellín, Colombia. <mcdiez@unal.edu.co>
Received: May 6, 2016; Accepted: October 18, 2016
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
Key words: Chlorophyll fluorescence, Gas exchange, Orchids, Photosynthetic pigments, Titratable acidity, Tropical crops.
RESUMEN
Vanilla planifolia es una orquídea neotropical, cuyos frutos producen la vainilla natural, un ingrediente fundamental para la industria alimenticia y cosmética. Debido a su importancia en el mercado internacional, se cultiva en varios países tropicales y recientemente ha comenzado su cultivo en Colombia. Esta especie requiere sombra para su desarrollo; sin embargo, se desconocen sus rangos óptimos de condiciones de iluminación. En este trabajo evaluamos el efecto de diferentes intensidades lumínicas sobre la fotosíntesis CAM, la fisiología, la morfología y el crecimiento de esta especie. Para esto, plantas jóvenes de vainilla se sometieron a cuatro tratamientos de iluminación relativa (IR) (T1=8%, T2=17%, T3=31% y T4=67%). La mayor proporción de asimilación de CO2 ocurre durante la noche en todos los tratamientos, lo cual confirmó que la vainilla es una especie CAM fuerte. Las plantas que crecieron bajo alta iluminación (67% IR), tuvieron casi la mitad de la fotosíntesis de las plantas en los tratamientos de iluminación intermedia (17 and 31%), lo cual coincide con la baja acumulación nocturna de ácidos en este tratamiento. Así mismo, la eficiencia fotoquímica del fotosistema II (Fv / Fm) mostró que en las plantas del 67% IR ocurrió estrés por alta radiación. De otro lado, las plantas de vainilla alcanzaron mayor longitud, área foliar, y biomasa total cuando crecieron bajo radiación intermedia (17 y 31% IR). Estos resultados sugieren que la alta radiación altera el funcionamiento de las plantas de vainilla, inhibiendo la fotosíntesis y el crecimiento, y que los ambientes altamente sombreados no afectan significativamente la fotosíntesis CAM de la vainilla; sin embargo, en el largo plazo, esta especie muestra mayor fotosíntesis y crecimiento bajo condiciones intermedias de radiación.
Palabras claves: Fluorescencia de la clorofila, Intercambio de gases Orquídeas, Pigmentos fotosintéticos, Acidez titulable, Cultivos tropicales.
Vanilla (Vanilla planifolia Andrews) is a hemiepiphyte orchid that faces seasonal water stress throughout its life cycle under natural conditions, which explain its CAM photosynthetic pathway (Goh and Kluge, 1989); likewise, this species faces different light environments: initially it grows in the shady interior of neotropical forests, and then, leaning on the tree, reaches the canopy where flowers and fruits at more light (Fouché and Jouve, 1999). Since vanilla plants are exposed to spatial and temporal (daily and seasonal) variations in the amount of radiation received (Fouché and Jouve, 1999), it is expected that this species had high acclimation capacity to different light environments, such as other epiphytes and hemiepiphytes of tropical forests (Zotz and Winter, 1994; Zotz and Andrade, 2001; Haslam et al., 2003).
Because of its importance in the world market of spices, vanilla is cultivated in many tropical countries (Bory et al., 2008). The success of this crop depends, among other factors, of the proper management of light because it is the environmental factor with the greatest influence on plant photosynthesis and therefore on growth, survival, and ultimately, the ability of acclimation to different habitats (Puthur, 2005). Hence, the importance of establishing the acclimation capacity of vanilla plants to different light environments and determining the most favorable for their growth.
The role of radiation on photosynthesis of CAM plants is related to the processes that occur during the four phases of this metabolism (Osmond, 1978), since the radiation can modulate their magnitude and duration (Cushman, 2001). The intensity of photosynthetically active radiation (PAR) during the day (Phase III), determines the rate of mobilization of organic acids from the vacuole (Barrow and Cockburn, 1982). On the other hand, the amount of radiation during the day also influences the extent of CO2 absorption at night (Phase I), affecting the abundance of carbohydrates generated through the Calvin cycle and glucogenesis, which are required for the provision of PEP at night (Nobel and Hartsock, 1983). Thus, the radiation during the day affects the extent of acid accumulation during the following night, and therefore, the amplitude of daily fluctuations of acids in CAM plants depends on radiation intensity (Lüttge, 2004).
]]> Sometimes low lighting can be limiting for CAM photosynthesis, such as in epiphytes of cloud forests during the rainy season (Pierce et al., 2002). Excess of radiation and over-energization of the CAM photosynthetic apparatus can also be limiting; systems of energy dissipation exhibited by C3 plants, such as photorespiration and the presence of carotenes, have also been reported in CAM plants (Lüttge, 2004).In regions where vanilla is cultivated, located mainly in the subtropical zone, different values of lighting have recommended for the proper development of plants: 30 - 80% of relative illumination (RI) in Mexico (Hernandez-Hernandez, 2011), 50 - 60% in Costa Rica (Varela, 2011), 60% in Reunion, Comoros and Madagascar Islands (Kahane et al., 2008), 30 - 50% in India (Anilkumar., 1994; Sarma et al, 2011; Zaubin et al, 2011), 50 - 60% in Australia (Exley, 2011), and 30 - 50% in Indonesia and Islands in the south Pacific (Bianchessi, 2004). The lower values of these ranges are generally considered appropriate for vegetative growth and the higher ones to stimulate flowering (Fouché and Jouve, 1999; Puthur, 2005). However, these recommendations are rarely based on published research or results that support higher growth or development of plants in these environments.
According to former reports, our hypothesis is that RI close to 30% is the most favorable for photosynthesis and growth of vanilla plants in the initial phase of establishment. To test it, the objective of this study was to evaluate the behavior of V. planifolia seedlings under different lighting levels in terms of: i) vegetative growth and morphological characteristics; ii) characteristics of CAM photosynthesis; iii) the response to extreme light environments, expressed as acclimation to low light conditions as well as the susceptibility and defense mechanisms to photo inhibition by high lighting.
MATERIALS AND METHODS
Experimental site
The trial was conducted in an area of alluvial terraces of flat topography in the inter-andean valley of the Cauca river (6o33'32'' N and 77°51''04'W), altitude of 540 m, on lands of the Agricultural Center Cotové (Universidad Nacional de Colombia). Data from the weather station located on the same site are: annual mean temperature of 27 °C, average annual precipitation of 1058 mm (bimodal distribution, with dry seasons from December to March and from June to August) and mean relative humidity of 75% (data of the Cotové Station, IDEAM). The site corresponds to the life zone tropical dry forest (sensu Holdridge, 2000).
Vanilla plants
Vanilla cuttings used in this research were about 80 cm long, taken from the apical portion of the branches of healthy and adult plants of V. planifolia from a crop established in San Pedro de Urabá (Antioquia, Colombia). Cuttings were hardened in the shade for two weeks, and then planted in plastic containers (6 L capacity); a wooden stand was previously installed in each container to serve as a tutor for subsequent plant growth.
Lighting treatments ]]> Shade houses of four different RI conditions were built: 8%, 17%, 31%, and 67%. Maximum values of photosynthetic active radiation (PAR) in the shade houses occurred between 13 and 14 h, and were 142, 369, 577, and 1285 µmol m-2 s-1 for the treatments of 8, 17, 31, and 67% RI, respectively. These values were obtained with quantum sensors (LI-190, LI-COR, Lincoln, Nebraska, USA) connected to a data logger (LI-1000, LI-COR, Lincoln, Nebraska, USA) installed for three days in one shade house by lighting treatment, with measurements every hour in cycles of 24 h, at the end of the first dry season of the year. The total daily radiation at full sun light averaged 2.4, 12.8, 28.7, and 43.8 mol m-2 d-1 in the treatments of 8%, 17%, 31%, and 67% RI, respectively. Shade houses were large enough (width: 4.5 m, length 6 m, and height 2.4 m) to ensure aeration and spacing between plants and prevent the occurrence of diseases. Vanilla plants were grown in the shade houses for six months before starting the measurement of variables to ensure acclimation to the corresponding light environment; then the monitoring of biometric parameters was done for twelve months. Plants received permanent irrigation to avoid differences by drought. Average temperatures in the shade houses varied between 25.3 oC in the treatment of 8% RI and 26.2 in the treatment of 67%.
Variables evaluated
CO2 exchange. Data of CO2 exchange were taken in fully expanded and healthy young leaves, located in the fourth internode from the terminal bud with a portable gas exchange system (LI 6400 XT, LI-COR Biosciences, Lincoln, NE, USA). The CO2 concentration was kept constant at a value of 400 µmol mol-1 and the block temperature at 26 °C. Automatic records were taken every two hours during cycles of 24 hours to three plants per treatment. For each RI treatment, one curve was estimated as the average of the three curves performed. The net absorption/release of CO2 was quantified for the day, night and the whole day, by integrating the areas under the 24 h curves of CO2 exchange (Griffiths, 1989).
Night accumulation of H+. To determine the nocturnal acid accumulation, the change of titratable acidity of leaf tissues between dusk and dawn of the next day was evaluated. To this, samples of 4.5 cm2 area were taken with a metal punch in young fully expanded leaves of five plants per treatment. Immediately after collected, samples were frozen in liquid nitrogen, brought to the laboratory and stored in an opaque container at -80 °C until processing, which consisted in immersing of samples in 70 mL of ethanol (20%) and boiling for 20 min. Titratable acidity of the solution was measured as the volume of NaOH (5 mM NaOH) necessary to reach a pH of 7.0 (Silvera et al., 2005) using a digital burette (Titrette, Brand, Wertleim, Germany) and a pH meter (Hanna 211, Woonsocket, RI, USA). With the values of titration, the H+ µequivalents were calculated (expressed in terms of leaf area). The change of titratable acidity of leaf tissue H+) was calculated as the difference between the μequivalents of H+ at dawn and the μequivalents of H+ at dusk the day before.
Biometric parameters of plants. After 18 months of growth of vanilla plants in each lighting treatment, leaf area was measured with a portable meter (LI-3000C, LI-COR Biosciences, Lincoln, NE, USA) and the total stem length with metric tape of all plants. Besides, three plants per repetition were harvested in each treatment to assess biomass of leaves, stems, and roots; these samples were oven dried (70 °C to constant weight) and weighed. Then, total biomass and specific leaf area (leaf area per gram of biomass) were calculated.
Chlorophyll fluorescence. The maximum quantum efficiency of photosystem II (PSII), called quantum yield potential, was measured with a portable fluorometer (OS30p, Optisciences, Hudson, NH, USA). This variable expresses the relationship between maximal fluorescence and variable fluorescence (Fv / Fm); Fv is the difference between maximal fluorescence (Fm) and the basal fluorescence (Fo). Measurements were performed in fully expanded and healthy young leaves, located at position three to seven from the terminal bud in three plants per repetition in each RI treatment; before sampling, leaves were dark-adapted for 30 min. Data were taken every two hours during 24 h.
Pigment analysis. Leaf discs of 1.5 cm2 were taken in three plants per repetition per RI treatment. After collected, discs were frozen in liquid nitrogen, transported to the lab, and stored at -80 °C until processing. Pigment extraction was performed with 80% acetone (Lichtenthaler and Wellburn, 1983; Poorter et al., 2011). Samples were centrifuged (5 °C for 5 min) and the supernatant was taken for reading in a Visible - UV spectrophotometer at 470, 647 and 663 nm (Evolution 600 UV-Vis, Thermo Scientific, Waltham, MA, USA). The following equations developed by Lichtenthaler (1987) were used to obtain the concentration of chlorophylls (Chlo) and total carotenes (Tot car) (mg/g):
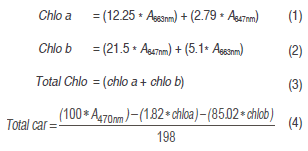
Experimental design and statistical analysis. Data were analyzed for a completely randomized design, with four RI treatments (67%, 31%, 17% and 8%) and four repetitions per treatment. Each repetition consisted of a shade house where six vanilla plants were installed (24 plants per treatment) for a total of 120 plants for the whole trial. For the statistical analysis, initially the assumptions of normality and homoscedasticity of each variable were evaluated with the tests of Shapiro-Wilk, Kolmogorov-Smirnov, and frequency histograms. Then, an analysis of variance (ANOVA) for each variable was performed. Finally, post hoc tests to determine significant differences among means were used. In order to evaluate the effect RI treatments on chlorophyll fluorescence throughout the day, a split plot model over time was used; variances of fluorescence were homogenized by ln (x) transformation (Steel and Torrie, 1980).
]]> RESULTS AND DISCUSSION
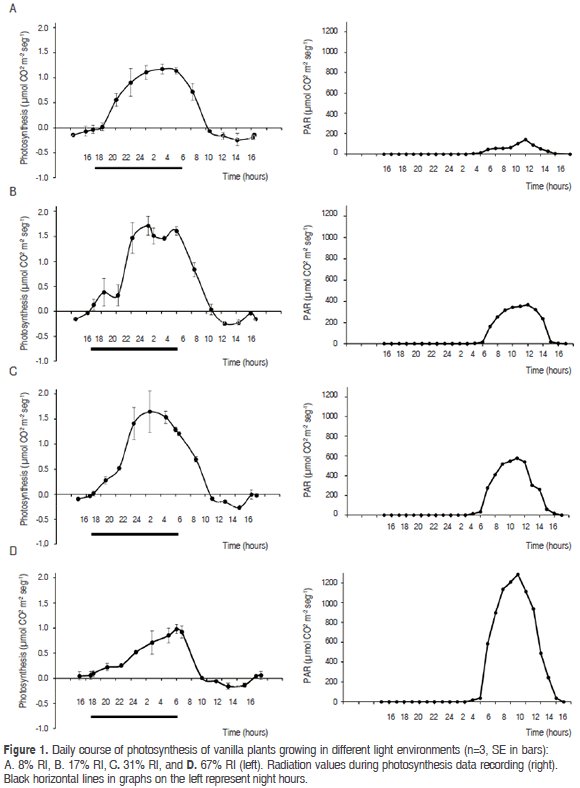
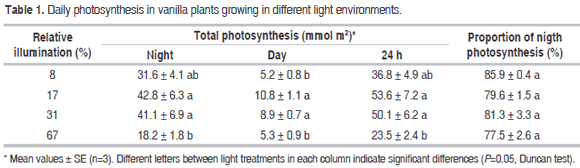
CO2 exchange. Most CO2 assimilation occurred at night in all treatments. At dawn, around 6:00 h, with the sunlight of the early hours of the morning, CO2 assimilation began to decline drastically until 10:00 h. From this time, with high solar radiation, CO2 assimilation was negative; after 16:00 h the uptake began to increase, but usually did not reach positive values until 18:00 h when it was already dark, so carbon assimilation did not occur at the end of the afternoon (Figure 1). The average values of maximum instantaneous assimilation (± SE) were 1.17 ± 0.10, 1.71 ± 0.19, 1.65 ± 0.42 and 0.98 ± 0.09 µmol m-2 s-1 for plants grown under 8, 17, 31 and 67%, RI respectively, and occurred late in the evening between 0 and 6 h. At night, during the Phase I of CAM metabolism (Osmond, 1978), plants absorbed the largest proportion of CO2 per unit of leaf area, equivalent on average to 81.2% of the total absorption for the 24 h period (Tabla 1), which confirms that vanilla is a strong CAM plant (Silvera et al., 2005).
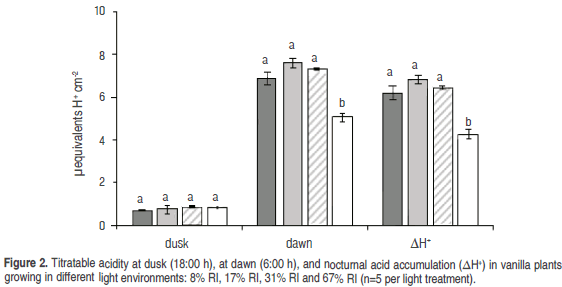
Nocturnal acid accumulation. No significant differences occurred among treatments in acidity values at the end of the afternoon (18:00 h), but they did early in the morning (6:00 h) (P=0.0005). Values of nocturnal acid accumulation H+) were significantly higher in the treatments of 8, 17 and 31% RI as compared with the treatment of 67% (P=0.0007) (Figure 2).
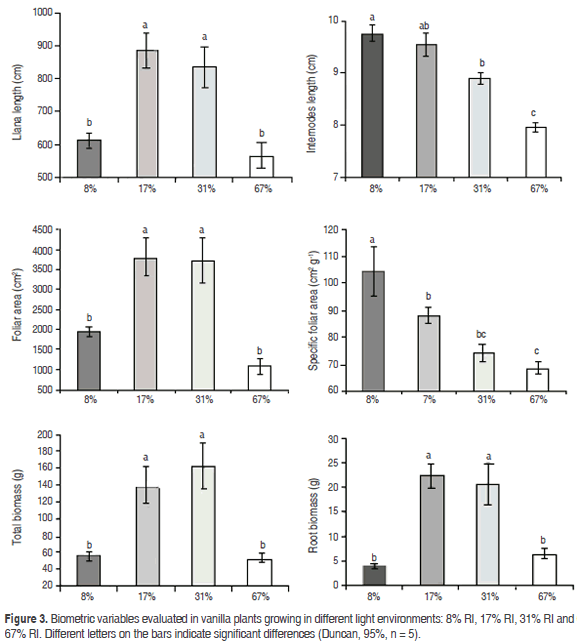
Biometric variables: Mean liana length was significantly higher (P=0.0017) in the intermediate RI treatments (17 and 31%) compared to low and high ones (8 and 67%): 860.1 cm vs. 589.8 cm, respectively (Figure 3), which represents an increase of 45.8% in plants grown under intermediate light; however, internodes became longer with the decrease in RI (P=0.0002); values were 9.7 and 7.9 cm in plants that grew at RI of 8 and 67%, respectively. Mean values of leaf area were also significantly higher (P=0.0003) in plants under intermediate RI treatments (17 and 31%) compared to low and high ones (8 and 67%): 3737.4 and 1510 cm2, respectively (Figure 3), which represents an increase of 147.4% in plants grown under conditions of intermediate light.
Specific leaf area was significantly greater in the most shaded treatment (8% RI) (P=0.0018), with a mean value of 104.4 cm2 g-1, which represents an increase of 51, 8% with respect to the most illuminated treatment (67% RI= 68.7 cm2 g-1). Similarly, root biomass was significantly higher in intermediate light treatments (17% and 31% RI) compared with treatments of high and low lighting (P=0.0002), with mean values of 21.55 g vs. 5.17 g, respectively. Finally, also the total biomass was significantly higher (P=0.0013) in the intermediate RI treatments (17 and 31%) compared with low and high illumination (8 and 67% respectively): on average 149, 2 vs. 53.9 g (176.8% increase).
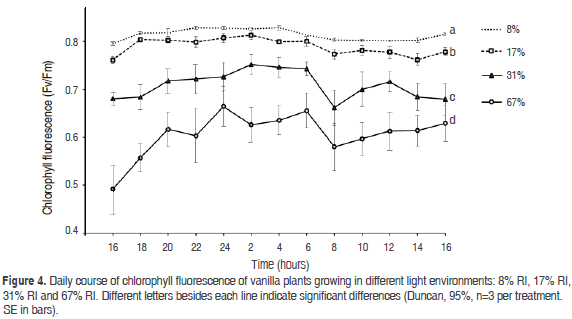
Chlorophyll fluorescence. Significant differences in values of chlorophyll fluorescence among illumination treatments (P=0.0000), as well as among hours along day (P=0.0000) and for the interaction treatment x hour (P=0.0032). The values were significantly higher in the RI of 8%, followed by 17%, 31% and 67% (Figure 4).
]]> Leaf content of photosynthetic pigments. The concentrations of chlorophyll a, chlorophyll b, and total chlorophyll were significantly higher in plants established in the most shaded treatment (8% RI) (P=0.0000 in all cases), followed by the intermediate (17 and 31% RI), and high illumination treatment (67% RI), which had the lowest values (Table 2). The concentration of carotenoids showed an inverse trend: it was significantly higher in plants developed in high illumination (P=0.0001), and decreased in more shaded treatments. The ratio chlorophyll a/b had no significant differences between lighting treatments (P=0.6266), while the ratio total chlorophyll / carotenoids was significantly higher in plants established in low light (P=0.0001).The results clearly show that the light environment of growing affects the CAM photosynthesis and vegetative growth of vanilla, with optimum illumination range at intermediate to low levels (8.6 - 12.8 mol m-2 d-1 of PAR, which the study area correspond to RI between 17 and 31%). Most values reported as suitable for vanilla growth in other regions where this species is cultivated, range between 30 and 60% RI (Anilkumar, 1994; Bianchessi, 2004; Kahane et al., 2008; Exley, 2011; Hernández-Hernández, 2011; Sarma et al., 2011; Varela, 2011; Zaubin et al., 2011); however, comparisons based on RI probably are biased without knowing the PAR values at these sites. Results suggest that vanilla has better acclimation capacity to low light environments (though with suboptimal growth) than to high lighting conditions.
The highest absorption values occurred in the intermediate to low light treatments. These results agree with those of nocturnal acid accumulation H+), which were higher in the treatments of 8, 17 and 31% RI; i.e., plants grown in environments RI intermediate to low (which absorbed more CO2) transformed that CO2 into organic acids along night (Dodd et al., 2002) and thus showed greater accumulation of acids in the vacuoles. During the day, the highest CO2 uptake occurred in the treatments of 17 and 31% RI, which specifically occurred early in the morning (Phase II of CAM metabolism), because during the rest of the day (Phases III and IV) only respiration occurred.
As a result of this balance of carbon absorption during the day and night, the largest net amount of CO2 absorbed along the 24 h period occurred under the intermediate -low light treatments, with much lower values in the high lighting treatment. Consequently, the highest values of growth were obtained in plants developed in the intermediate treatments RI (17 and 31%), with daily PAR of 8.6 and 12.8 mol m-2 d-1, expressed as liana length, leaf area, root biomass, and total biomass (Figure 3). In these treatments, the maximum values of instantaneous radiation were 369 and 577 µmol m2 s-1, respectively, and occurred between the 13:00 and 14:00 h. Accordingly, the environments which favor the photosynthesis and growth of vanilla are apparently those with PAR levels at noon between these two values, while environments of greater radiation (for example, the ones in the 67% or RI: 1201, 4 µmol m2 s-1 at noon) seem to limit the photosynthesis and development of this species. These results are consistent with values reported in India as favorable for the vegetative development of vanilla, of 300 -600 µmol m2 s-1 (Puthur, 2005), although it would be necessary to know the total daily PAR for this region, in order to evaluate whether the data are comparable.
Plants growing under extreme lighting conditions (high and low), had the lowest values of growth, although showed some mechanisms of acclimation. On the one hand, plants that grew in 8% RI showed several adaptations to be more efficient under low light conditions, such as longer internodes and higher specific leaf area, since in low-radiation, an elongated morphology and larger leaf blades increase light uptake (Walters, 2005). However, the total leaf area per plant decreased probably as a consequence of the lower resources available under that condition; perhaps for that reason, plants under this light condition showed the lower growth rate over a long time period (Shipley, 2002), despite no significant differences of total daily photosynthesis per unit leaf area were found with intermediate RI treatments (Table 1).
At the other extreme, the low CO2 assimilation and growth in plants developed under high light (67% RI) could be explained by the signs of chronic photo inhibition detected, which damages the reaction centers of PSII and causes photo destruction of photosynthetic pigments (Powles, 1984). The PSII efficiency (Fv / Fm) in this study declined from 0.81 ± 0.001 in the treatment of 8%, indicative of absence of photo inhibition (Powles, 1984; Werner et al., 2001), to 0.61 ± 0.001 in the 67% RI treatment, indicative of presence of photo inhibition.
Plants from shadow environments are especially susceptible to chronic photo inhibition when are exposed to high light intensities. Under such conditions, plants show low potential quantum yield of photosynthesis as a result of fewer reaction centers in the PSII, resulting in a lower Fv / Fm throughout the day (Murchie et al., 2009). A further indication of irreversible damage is that leaves of plants grown in high light (67%) had white spots by chlorosis, probably as a result of a higher chlorophyll degradation, because leaves could not effectively channel the high energy received in photochemical reactions, which leads to chlorophyll whitening (Anderson, 1986).
The concentration of chlorophylls (a, b and total) was higher in leaf tissues of plants growing in lower lighting (8% RI). Under these conditions, the synthesis of greater amount of chlorophyll is a strategy of acclimation to increase the efficiency of photon capture and counteract the lower radiation reaching the leaf (Anderson, 1986). Perhaps this high chlorophyll content was effective to avoid very low values of total daily photosynthesis per unit leaf area, despite the limitations imposed by low radiation; this response is a further evidence in favor of the shade-tolerant character of vanilla.
It has been reported that plants adapted to low radiation have low values of the chlorophyll a / chlorophyll b ratio (approximately 2) and that plants adapted to high radiation have high values (about 2.8 and more) (Anderson, 1986). In this study no significant differences in this ratio were found among vanilla plants growing under different RI, whose average value was 2.2 (Table 2). Low values of this ratio are correlated with high degree of stacking of thylakoid membranes in the chloroplast, which increases the area of the cross section of the grana and produces increased risk of photo inhibition (Anderson and Aro, 1994). This inability of vanilla plants growing under high light intensity to increase chlorophyll a / chlorophyll b ratio, suggests that there were not significant reorganization of thylakoid membranes, which would allow lower stacking and thus avoid photo destruction of photosynthetic apparatus in plants exposed to high radiation (67% RI), which is a further evidence of the shade- tolerant character of this species.
]]>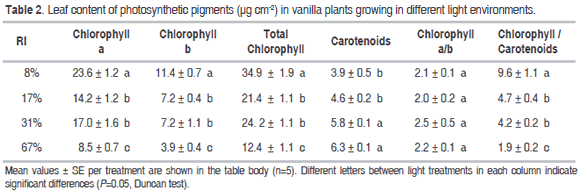
Carotenoids protect chlorophyll pigments against high radiation under conditions of light stress (Yamamoto and Bassi, 1996) and therefore, the ratio chlorophyll / carotenes is an indicator of the protection against the high radiation, since carotenes contribute to dissipate the excess of radiation (Cazzonelli et al., 2011). Low values, indicative of greater photo oxidative damage, were found in plants exposed to 67% RI (Table 2), whereas in plants developed under 17 and 31% RI, were intermediate, and higher values in plants under 8% RI. Vanilla plants showed ability to capture and use light by increasing the concentration of pigments when grown at low radiation; the decrease of chlorophyll / carotenes ratio under high radiation suggests their inability to minimize the damage caused by high radiation, which is a further evidence of their inability to acclimate to high radiation environments.
Since the proportion of carotenoids vs. chlorophyll decreased with increasing radiation intensity (Table 2), vanilla plants grown in high light did not have enough mechanisms to protect against the photo-destructive damage from high radiation. Such failure was evidenced in the low chlorophyll content, low CO2 assimilation, less chlorophyll fluorescence, and low productivity of plants exposed to radiation above 800 μmol m-2 seg-1. These results confirm that shade plants, do not have well developed mechanisms to counteract the effects of photo inhibition.
Therefore, shade plants such as vanilla are very sensitive to high radiation; this causes inactivation of reaction centers and inhibition of electron transport, since the antennas of their photosynthetic apparatus are unable of channeling the light energy into the photochemical reaction centers. The excess of energy can induce the production of reactive oxygen species and free radicals (Powles, 1984) which break the DNA, destroy the function of proteins, and are responsible for peroxidation of lipids, thus causing damage to the plant metabolism and decreasing the rate of photosynthesis and growth.
CONCLUSIONS
Most CO2 assimilation occurred along night in all treatments, which confirms that vanilla is a strong CAM plant. Results suggest that high radiation alters the functioning of vanilla plants, inhibiting photosynthesis and growth, and that highly shaded environments not significantly affected the CAM photosynthesis of vanilla; however, in the long-term this species showed higher photosynthesis and biomass growth under intermediate levels of radiation (17-31% RI).
ACKNOWLEDGEMENTS
This work was done as part of the project "Ecophysiology vanilla cultivation (Vanilla planifolia Andr.)", funded by the Administrative Department of Science, Technology and Innovation of Colombia - COLCIENCIAS- and the Universidad Nacional de Colombia (contract 618-2011).
]]>REFERENCES
Anderson JM and Aro E. 1994. Grana stacking and protection of photosystem II in thylakoid membranes of higher plant leaves under sustained high irradiance: an hypothesis. Photosynthesis Research 41 (2): 315-326. doi: 10.1007/BF00019409 [ Links ]
Anderson JM. 1986. Photoregulation of the composition, function, and structure of thylakoid membranes. Annual Review of Plant Physiology 37 (1): 93-136. doi: 10.1146/annurev.pp.37.060186.000521 [ Links ]
Anilkumar AS. 2004. Vanilla cultivation: A profitable agri-based enterprise. Kerala Calling 1: 26-30. [ Links ]
Barrow SR and Cockburn W. 1982. Effects of light quantity and quality on the decarboxylation of malic acid in crassulacean acid metabolism photosynthesis. Plant physiology 69 (3): 568-571. doi: 10.1104/pp.69.3.568 [ Links ]
Bianchessi P. 2004. Vanilla: agriculture and curing techniques. A photographic handbook for vanilla farmers. Venui Vanilla Co. Santo, Vanuatu. 62 p. [ Links ]
Bory S, Grisoni M Duval MF and Besse P. 2008. Biodiversity and preservation of vanilla: present state of knowledge. Genetic Resources and Crop Evolution 55 (4): 551-571. doi: 10.1007/s10722-007-9260-3 [ Links ]
Cushman JC. 2001. Crassulacean acid metabolism. A plastic photosynthetic adaptation to arid environments. Plant Physiology 127(4): 1439-1448. doi: 10.1104/pp.010818 [ Links ]
Dodd AN, Borland AM, Haslam RP, Griffiths H and Maxwell K. 2002. Crassulacean acid metabolism: plastic, fantastic. Journal of Experimental Botany 53 (369): 569-580. doi: 10.1093/jexbot/53.369.569 [ Links ]
Exley R. 2011. Vanilla Production in Australia. pp. 69-78. In: Havkin-Frenkel, D and Belanger F (ed.). Handbook of vanilla science and technology. Wiley-Blackwell, West Sussex, UK. 339 p. [ Links ]
Fouché JG and Jouve L. 1999. Vanilla planifolia: history, botany and culture in Reunion Island. Agronomie 19 (8): 689-703. [ Links ]
Griffiths H. 1989. Carbon dioxide concentrating mechanisms and the evolution of CAM in vascular epiphytes. pp. 42-86. In: Lüttge, U. (ed.). Vascular plants as epiphytes. Springer Berlin- Heidelberg, GE. 270 p. [ Links ]
Haslam R, Borland A, Maxwell K and Griffiths H. 2003. Physiological responses of the CAM epiphyte Tillandsia usneoides L. (Bromeliaceae) to variations in light and water supply. Journal of Plant Physiology 160 (6): 627-634. doi: 10.1078/0176-1617-00970 [ Links ]
Hernández-Hernández J. 2011. Mexican Vanilla Production. pp. 1-25. In: Havkin-Frenkel D. and Belanger F (ed.). Handbook of Vanilla Science and Technology. Wiley-Blackwell, West Sussex, UK. 339 p. [ Links ]
Holdridge LR. 2000. Ecología basada en Zonas de Vida. Instituto Interamericano de Cooperación para la Agricultura - IICA, San José, Costa Rica. 216 p. [ Links ]
Kahane R, Besse P, Grisoni M, Le Bellec F and Odoux E. 2008. Bourbon vanilla: natural flavour with a future. Chronica Horticulturae 48 (2): 23-28. [ Links ]
Lichtenthaler H and Wellburn AR. 1983. Determination of total carotenoids and chlorophyll a and b of leaf extracts in different solvents. Biochemical Society Transactions 11(5): 591-592. doi: 10.1042/bst0110591 [ Links ]
Lichtenthaler HK 1987. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology 148:350-382. [ Links ]
Lüttge U. 2004. Ecophysiology of crassulacean acid metabolism (CAM). Annals of Botany 93(6): 629-652. doi: 10.1093/aob/mch087 [ Links ]
Murchie EH, Pinto M and Horton P. 2009. Agriculture and the new challenges for photosynthesis research. New Phytologist 181(3): 532-552. doi: 10.1111/j.1469-8137.2008.02705.x [ Links ]
Nobel PS and Hartsock TL. 1983. Relationships between photosynthetically active radiation, nocturnal acid accumulation, and CO2 uptake for a Crassulacean Acid Metabolism plant Opuntia ficus-indica. Plant physiology 71(1): 71-75. doi: 10.1104/pp.71.1.71 [ Links ]
Osmond CB. 1978. Crassulacean Acid Metabolism: a curiosity in context. Annual Review of Plant Physiology 29(1): 379-414. doi: 10.1146/annurev.pp.29.060178.002115 [ Links ]
Pierce S, Winter K and Griffiths H. 2002. The role of CAM in high rainfall cloud forests: an in situ comparison of photosynthetic pathways in Bromeliaceae. Plant, Cell and Environment 25(9): 1181-1189. doi: 10.1046/j.1365-3040.2002.00900.x [ Links ]
Poorter H and de Jong-Van Berkel Y. 2011. Chlorophyll extraction and determination. In: Prometheus Wiki contributors, http://www.publish.csiro.au/prometheuswiki/tiki-pagehistory.php?page=Chlorophyll extraction and determination&preview=11; accessed: November 2015. [ Links ]
Powles SB. 1984. Photoinhibition of photosynthesis induced by visible light. Annual Review of Plant Physiology 35 (1): 15-44. doi: 10.1146/annurev.pp.35.060184.000311 [ Links ]
Puthur J. 2005. Influence of light intensity on growth and crop productivity of Vanilla planifolia Andr. General and Applied Plant Physiology 31(3-4): 215-224. [ Links ]
Sarma YR, Thomas J, Sasikumar B and Varadarasa S. 2011. Vanilla production in India. In: Odoux E and Grisoni M (ed.). Vanilla. Medicinal and aromatic plants-industrial profiles. CRC Press, Taylor and Francis Group, Boca Ratón, FL, USA. 420 p. [ Links ]
]]>Shipley B. 2002. Trade-offs between net assimilation rate and specific leaf area in determining relative growth rate: relationship with daily irradiance. Functional Ecology 16(5): 682-689. doi: 10.1046/j. 1365-2435.2002.00672.x [ Links ]
Silvera, K, Santiago LS and Winter K. 2005. Distribution of Crassulacean Acid Metabolism in orchids of Panama: evidence of selection for weak and strong modes. Functional Plant Biology 32(5): 397-407. doi: 10.1071/FP04179 [ Links ]
Steel RGD and Torrie JH. 1980. Principles and procedures of statistics: a biometrical approach. Mc Grow-Hill, New York, USA. 633 p. [ Links ]
Walters RG. 2005. Towards an understanding of photosynthetic acclimation. Journal of Experimental Botany 56(411): 435-447. doi: 10.1093/jxb/eri060 [ Links ]
Varela E. 2011. Vanilla Production in Costa Rica. pp. 40-49. In: Havkin-Frenkel D and Belanger F (ed.). Handbook of Vanilla Science and Technology. Wiley-Blackwell, West Sussex, UK. 339 p. [ Links ]
Yamamoto HY and Bassi R. 1996. Carotenoids: localization and function. pp. 539-563. In: Ort D and Yocum CF (ed.). Oxygenic Photosynthesis: The Light Reactions. Springer, Netherlands. doi: 10.1007/0-306-48127-8_30 [ Links ]
Zaubin R, Tombe M and Liew ECY. 2011. Vanilla production in Indonesia. pp. 283-294. In: Odoux E. y Grisoni M (ed.). Vanilla. Medicinal and Aromatic Plants-Industrial Profiles. CRC Press, Taylor and Francis Group, Boca Ratón, FL, USA. 420 p. [ Links ]
]]>Zotz G and Winter K. 1994. Annual carbon balance and nitrogen use efficiency in tropical C3 and CAM epiphytes. New Phytologist 126 (3): 481-492. doi: 10.1111/j.1469-8137.1994.tb04245.x [ Links ]
Zotz G and Andrade JL. 2002. La ecología y la fisiología de las epífitas y las hemiepífitas. pp. 271-296. In: Kattan, G.H. y M.R. Guariguata (ed.). Ecología y conservación de bosques neotropicales. Libro Universitario Regional, San José, Costa Rica. 691 p. [ Links ]
]]>